
Abstract
Identifying tissue sources of HIV that rebound following “failure” of antiretroviral therapy (ART) is critical to evaluating cure strategies. To assess the role of the uterine cervix and peripheral blood mononuclear cells (PBMC) as viral reservoirs, nearest-neighbor phylogenetic analyses compared genetic relatedness of tissue sequences during ART suppression to those detected in plasma at viral rebound. Blood and genital tract specimens from a natural history cohort of HIV-infected women were collected over 5 years. HIV DNA sequences extracted from PBMC and cervical biopsies during ART suppression and plasma RNA from rebound (defined as HIV RNA >3 log10 copies/mL) were derived by single-genome amplification. Phylogenetic and nearest-neighbor analyses of HIV env sequences and drug resistance in pol sequences were compared between tissues. Nine instances of plasma viral rebound (median HIV RNA 3.6 log10 c/mL; IQR: 3.1–3.8) were detected in 7 of 57 women. Nearest-neighbor analyses found rebound plasma sequences were closer to uterine cervical sequences in 4/9 (44%), closer to PBMC in 3/9 (33%), and ambiguous in 2/9 (22%) cases. Rebound plasma clades (n = 27) shared identical sequences in seven instances with the cervix versus two with PBMC. Novel drug resistance mutations were detected in 4/9 (44%) rebounds. The observed tendency for greater sharing of identical HIV variants and greater nearest-neighbor association between rebounding plasma and uterine cervical versus PBMC sequences suggests that the uterine cervix may be a relevant HIV reservoir. The cervix, a readily accessible tissue in women that can be repeatedly sampled, could help assess the HIV reservoir when evaluating cure strategies.
Introduction
Antiretroviral therapy (ART) can successfully suppress HIV replication, but does not eradicate the virus. 1,2 Treatment failures and analytic treatment interruptions result in rebound of HIV RNA. 3 –11 Rebound viruses are released from long-lived reservoirs that persist during ART. 1,4,12 –16 Central memory cells have been described as a primary HIV reservoir. 2 These cells reside to a greater degree in tissues compared with blood. 17 –19 However, the relative role of tissues versus blood reservoirs as a source of plasma HIV rebound viruses remains controversial. 8,15,20 –22
Many phylogenetic studies of rebound HIV, or viral RNA detected at virologic failure or after stopping ART, suggest the viruses arise from outside the blood. 1,4,12 –16 , 22–24 Studies of blood and tissues have identified peripheral blood mononuclear cell (PBMC), lymph nodes, and ileum as likely sources. 22,24,25 Determining the cell and tissue sources of plasma rebound viruses could inform researchers as to the relevant targets for therapeutic interventions to eradicate the virus. We undertook a study to determine whether the uterine cervix, a tissue that is readily accessible and can be repeatedly biopsied without morbidity, is a potential source of rebound viruses during ART.
Methods
Ethics statement
The study was conducted at the University of Washington and at the University of Rochester following approval by both Institutional Review Boards. All study participants provided written informed consent.
Study design
Women with chronic HIV infection on ART were enrolled in a 5-year observational study as previously described. 26 During the first year, blood and genital specimens were collected quarterly during the luteal phase of the menstrual cycle, and in the subsequent 4 years, collected biannually. Participants for this substudy were selected if their plasma virus rebounded (defined as HIV RNA ≥3.0 log10 copies/mL) from ART suppression (HIV RNA <1.5 log10 copies/mL). HIV DNA sequences were generated from PBMC and cervical biopsies from specimens collected during ART suppression before the plasma rebound. RNA from plasma and cervical vaginal lavage (CVL) specimens had HIV env and pol sequences generated from time points when HIV RNA became transiently or persistently detected.
Specimen processing, nucleic acid extraction, and real-time quantification
Blood plasma and PBMC were separated by density centrifugation (Accuspin™; Sigma-Aldrich, St. Louis, MO), and stored at −80°C. 26 The nucleic acids from up to 0.5 mL of CVL and 2 mL of plasma were extracted with silica (miniMag™; BioMerieux, Durham, NC). 26 HIV RNA was quantified in plasma and CVL by a gag real-time PCR assay (Roche Diagnostics, Indianapolis, IN; and ABI Prism 7700, Foster City, CA). 26 DNA was extracted (Gentra Systems, Minneapolis, MN) from PBMC and 5-μm-thick sections from frozen cervical biopsies 26 from the nearest time point before HIV plasma rebound at which cervical biopsies were available. 26
Derivation of HIV RNA and DNA genomes by single-genome amplification
RNA, after reverse transcription (Superscript III™; Invitrogen, Carlsbad, CA), 27 and DNA underwent single-genome amplification (SGA) of the C2–V5 region of HIV env and the region of pol that encodes protease and reverse transcriptase. 26,28 HIV-1 env and pol genomes were generated using limiting dilution of extracted nucleic acids (RNA or DNA) from a specimen into ∼100 PCR, resulting in ∼30% of reactions yielding amplicons, followed by direct sequencing of PCR products. 28
To determine whether HIV DNA contaminated RNA extracted from plasma and cervical secretions, purified nucleic acids from three specimens were pooled and PCR amplified without the reverse transcription enzyme. 26
Construction of phylogenetic trees and env sequence analysis
Sequences were assembled and checked for read errors and hypermutation.
26
An all-inclusive phylogram was constructed to identify mix-ups between specimens (data not shown). Maximum likelihood phylogenetic trees were constructed for each participant's sequences using DIVEIN (
Statistical analyses
Differences in divergence between each individual's tissues across time points were assessed using a paired t-test (GraphPad, La Jolla, CA) when data were normally distributed. Differences in genetic diversity within a participant were performed using a t-test developed by Gilbert et al., 32 and implemented in DIVEIN. Each participant's HIV blood and tissue sequences were analyzed using the Slatkin–Maddison analysis 33 and Hudson's nearest-neighbor (NN) tests for compartmentalization, 34,35 implemented in HyPhy. 36 The observed proportions of the plasma sequences' NN coming from each comparator tissue group (e.g., genital DNA) was calculated. Then under the assumption of random matching, the expected probability (proportion) that any plasma sequence would have as its NN, a sequence from a given comparator tissue group, is the number of sequences from that comparator tissue group divided by the total # of all comparator sequences (e.g., expected proportion from genital DNA = (# genital DNA sequences/(# total comparator sequences per patient)]. Confidence intervals for the expected proportions were derived using 10,000 permuted samples by randomly shuffling (permuting) the comparator tissue group labels, reidentifying the NNs, and recalculating the proportions. If the observed proportion fell outside of the permutation-based confidence intervals, this was taken as statistically significant evidence that plasma rebound sequences were more closely related to the uterine cervix, PBMC, or when within the confidence intervals of both, was termed “ambiguous”. 37
Results
Clinical characteristics
A total of 53 of the 57 enrolled women had ART suppression and sufficient follow-up for evaluation. Among these, seven women experienced nine HIV RNA plasma rebounds (median 3.6 log10 copies/mL interquartile range (IQR): 3.1–3.8 log10 copies/mL) (Table 1), with two of the seven participants having two rebounds. The women were all premenopausal with a median age of 38 (IQR: 35–45 years), with relatively low (<500) CD4 counts throughout the study (median: 318 cells/μL, IQR 250–492), and were primarily African American (Table 1). The median time between the last documented ART suppression (<1.5 copies/mL) and the rebound was 4 months (IQR: 3–6 months). The causes of viral rebounds, and whether associated with intermittent adherence or additive mutations leading to failure of the ART regimen, was not assessed in this study.
Clinical Parameters for Women Experiencing Viral Rebound from the Women's HIV Interdisciplinary Network Cohort Followed for Up to 5 Years
Bold indicates the study visit(s) at which plasma HIV RNA rebound was detected.
NA, not available.
HIV env and pol phylogenetic and env NN phylogenetic analyses
Specimens collected during ART suppression before plasma HIV rebound yielded a median of 15 env (IQR 11–33) and 14 pol (IQR 11–19) sequences from PBMC, and a median of 14 env (IQR 13–21) and 13 pol (IQR 12–16) sequences from uterine cervix DNA. At the time of plasma viral rebound, a median of 18 env (IQR: 13–23) and 18 pol (IQR: 14–20) sequences were generated from the plasma. A median of 4 env (IQR: 4–8) and 7 pol (IQR: 2–12) sequences were obtained from CVL RNA from Participants 1 and 6 who both had detectable HIV RNA in cervical secretions.
HIV plasma RNA rebound sequences were identical in env and pol genes to HIV DNA sequences during ART suppression of four participants; two were identical to cervical sequences (Supplementary Fig. S1, Participants 4, 7); one to PBMC (Supplementary Fig. S1, Participant 5); and one to both tissues (Supplementary Fig. S1, Participant 6).
Nearest neighbor analysis demonstrated that HIV RNA sequences from 4/9 (44%) plasma rebounds in three women were closer to sequences from the uterine cervix versus PBMC; 3/9 (33%) rebounds were closer to PBMC, and 2/9 (22%) rebounds were ambiguous (Table 2). Both viral rebounds from Participant 6 were closer to the uterine cervix than the PBMC, however, each rebound clustered in different locations on her phylogenetic tree. Viral rebounds from Participant 7 included one rebound closer to the PBMC and the other is ambiguous, but the two rebounds have different locations on the tree. In this small sample of rebound virus, there did not appear to be a relationship between the time from suppression to rebound and the tissue source of that rebound (Table 2).
Nearest-Neighbor Phylogenetic Analysis of HIV Env Sequences Comparing the Genital and Peripheral Blood Mononuclear Cell to the Plasma Rebound
Cell-associated sequences were amplified from the available tissues before plasma rebound, where HIV RNA was suppressed to <1.5 log10 copies/mL in the plasma and in most cases the tissues. The number of sequences amplified from the tissues was pooled and used to establish the observed proportion of each tissue and used as a comparator for the referent rebound plasma sequences to establish the expected proportion of each tissue and a CI.
Pair-wise distances compared the nearest neighbor of rebound plasma viral sequences to genital tract versus PBMC sequences from antecedent ART-suppressed time point. The actual proportion of rebound plasma sequences with a nearest neighbor to each tissue was calculated based on the sequence frequency from each tissue are shown. The expected proportion is the predicted proportion under the assumption of random matching of the plasma rebound to either PBMC or tissues. The significant (p < .05) variations of actual from expected proportions support a closer genetic linkage of rebound plasma virus to the uterine cervix in 4/9 episodes, to PBMC in 4/9 rebound episodes, and ambiguous in 1/9 episodes (indicated in bold).
ART, antiretroviral therapy; PBMC, peripheral blood mononuclear cell.
Compartmentalization of HIV sequences by tissues was observed in a total of three participants (Table 3) (Participants 2, 5 and 7), driven by large phylogenetic clades of monotypic (identical) sequences in env and pol in one, and in env in the two others (Table 3). 26 In all cases, if the monotypic sequences were collapsed into a single sequence, compartmentalization was no longer observed (data not shown).
Compartmentalization of Viral Sequences Between Peripheral Blood Mononuclear Cells and Cervical Biopsy Sequences During Suppressive Antiretroviral Therapy at Study Visit Before Rebound of Plasma HIV RNA Is Infrequent
Slatkin–Maddison and Snn tests for compartmentalization demonstrate that sequences from the genital tract and blood frequently mix and only rarely does virus from these tissues appear compartmentalized. The compartmentalization observed in Participant 2, 5, and 7 was primarily due to multiple identical sequences, as when collapsed into a single sequence before analysis compartmentalization was no longer detected.
Bold values indicate those with statistically significant evidence of compartmentalization.
Viral variants predicted to use the X4 coreceptor were detected in the PBMC and cervical DNA during ART suppression in 3/7 (43%) (Participants 1, 6, and 7), and as a majority (>50%) variant in the plasma rebound HIV RNA in 2/3 women (Participants 6 and 7), but the frequency decreased in both participants' second rebound.
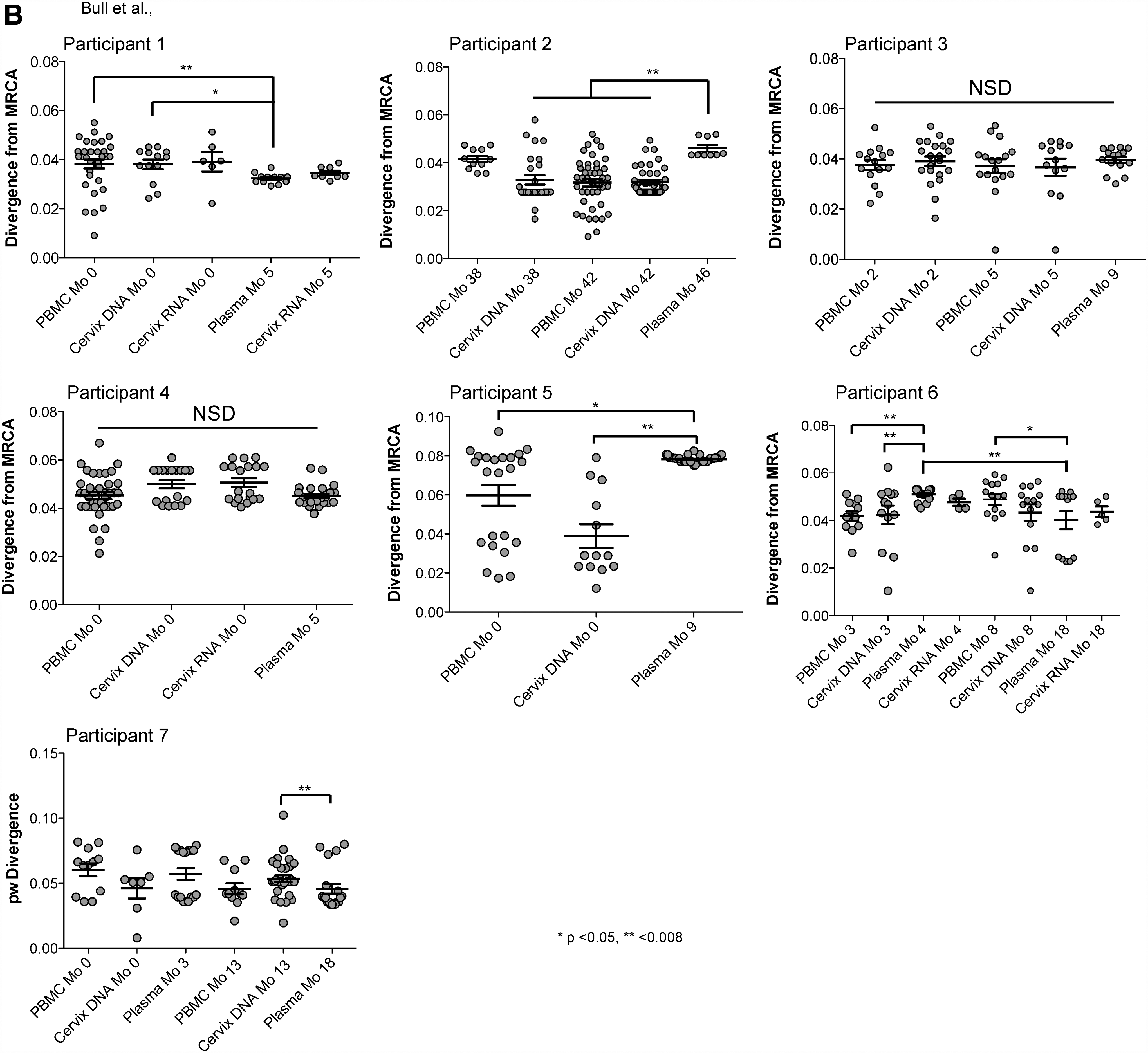
HIV env sequences in rebound plasma were less diverse compared with PBMC and uterine cervix DNA sequences from ART suppression (p < .05) in 5/9 (56%) rebounds (Fig. 1 and Supplementary Fig. S1, Participants 1–3, 5, and 6 study month 4), and similar in 4/9 (44%) (Participants 4, 6 (at study month 8), and 7 both rebounds). Sequences from plasma rebound HIV RNA diverged to a greater degree from the MRCA compared with HIV DNA during ART suppression in 3/9 (33%) rebounds (Supplementary Fig. S1B; Participants 2, 5, and 6 at study month 4), decreased in 2/9 (22%) (Participants 1 and 6 at study month 18), and similar in the other 4/9 (44%) rebounds (Participants 3, 4, and 7 at both rebounds).
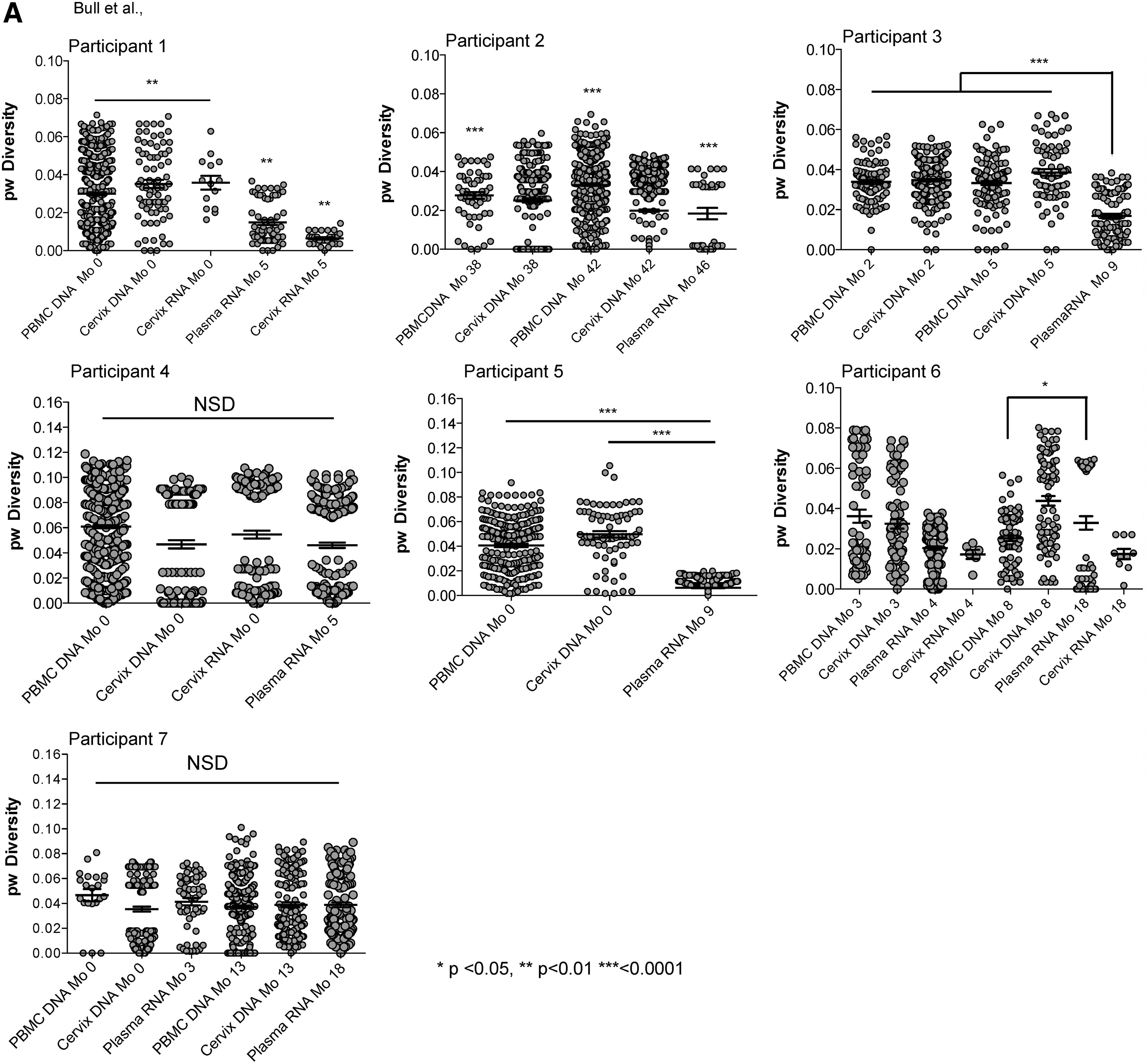
HIV env pairwise diversity and divergence plots. Maximum likelihood phylogenetic analyses of HIV C2–V5 env single-genome sequences from PBMC and cervical DNA when HIV was suppressed by ART and viral rebound.
26
HIV drug resistance
All participants had virus with drug resistance mutations detected among viral DNA sequences in tissues before plasma RNA rebound. The majority of participants' (6/7, 86%) plasma rebound HIV RNA sequences included mutations associated with drug resistance. The mutations were detected as three patterns: (1) the same mutations detected during ART suppression before plasma HIV RNA rebound (Participants 1, 4, and 6 at study month 4); (2) novel resistance mutations at low frequencies (Participants 2, 5, 6 at study month 18 and 7), or (3) novel mutations at high frequencies (Participants 3 and 6 at study month 18) (Supplementary Table S1).
Discussion
This study contributes novel observations to the literature that explores tissue sources of plasma HIV RNA rebound, 20,21,24,38,39 with an evaluation of the uterine cervix as a potential source of HIV plasma rebound. Our observations that (1) nearly half of the participants' plasma HIV RNA rebound sequences were genetically closer to, and (2) included sequences identical to the uterine cervix during ART suppression, suggest the cervix and/or tissues with similar viral variants include and/or have frequent genetic exchange with relevant HIV reservoirs during ART suppression. These observations suggest that the uterine cervix offers a readily accessible tissue where multiple biopsies are well tolerated 26 that could be examined during cure studies. In contrast to previous studies, 4,8,15 plasma viral rebound sequences from participants included sequences identical to PBMC sequences from ART suppression. This observation suggests that some plasma rebounds may come from circulating cells. 11,24,25
In the current study, most participants had a history of multiple ART regimens and episodes of virologic failure. Thus, the viral rebounds included re-emergence of drug-resistant variants selected by past regimens that were detected in the uterine cervix and/or blood during suppressive ART. Additionally, there was one participant with a novel drug-resistant variant detected in the plasma rebound that was not detected in the tissues (blood or cervix) during suppressive therapy that was likely selected due to low levels of one or more ARV in their ART regimen. 40 This suggests that rebounds were from impotent ART regimens, with additive selection of mutant variants dispite adherence, or intermittent adherence to ART.
Studies of HIV RNA plasma rebound from suspension of ART have primarily examined genetic distances compared with PBMC sequences to infer the likelihood that circulating cells are a source of rebounding viruses, and in most cases, the observed distances suggested that PBMC were not the source of the rebounding viruses. 20,22,24 Gastrointestinal tissues were evaluated in four studies, with conflicting results; one found the gut was not a source of viral rebound, 20 whereas the other three found evidence that the gut 22,24,25 and lymphoid tissues may be a significant source of plasma rebound. 24 In the current study, we found that genetic linkages to rebound sequences varied across the participants, with almost half being closer to sequences from the uterine cervix compared with a third to PBMC. However, because other tissues were not evaluated, we cannot preclude the emergence of rebounding virus from other tissues. Collectively, ours and others' studies 20,21,24,38,39 suggest that multiple tissues, likely those where memory cells are prevalent, serve as a source for or share viruses that emerge during for HIV plasma rebound. Preliminary studies of HIV DNA sequences from multiple tissues from autopsies of individuals dying when ART was suppressed found the same clones of virus extending across multiple tissues. 41 Of note is that the uterine cervix is relatively simple to sample and well tolerated by the participant, and appears to be a relevant infectious reservoir; thus, it could be evaluated during cure treatment interruptions.
Approximately 40% of participants' rebound viral sequences in this study grouped within a single clade within a phylogenetic tree of their sequences, is consistent with their rebound originating from a cellular clone undergoing proliferation 22,25 or being produced from recently infected cells that are undergoing bursts of viral replication. 8,15,22,24,42 –44 Studies that employed phylogenetic analysis similar to ours also showed large clades of identical sequences in the rebounding plasma viremias. 8,15,22,24,45,46 Four of the five studies that performed phylogenetic analysis of PBMC or tissues compared with rebounding virus within weeks after treatment interruption, noted diverse viral sequences that suggested rebound originated from multiple sources similar to several participants in this study. 8,22,24,45
Variants predicted to use the X4 receptor were found in the plasma viral rebound in a minority of participants (3/7, 43%). Previously we 47 and others 44 found that X4 variants are prevalent in low-level viremias of otherwise ART-suppressed participants. The presence of X4 variants in the plasma rebound might reflect the late disease stage of participants 48 –50 or it could indicate a previously unappreciated latent reservoir in naive T cells or those of the myeloid lineage that express high concentrations of the CXCR4 receptor. 42,51 –55
The number of women in our study of rebound virus was limited because most in this cohort experienced ART suppression. The number of women was too small for statistical comparisons of timing of rebound from ART suppression and genetic similarity of cervix or PBMC sequences. Others' work has shown increases in inflammatory biomarkers as HIV-infected women age into menopsause, and none of the women in this study was entering menopause, 56 but there are, to date, no studies specifically looking at menopause or drug use as they relate to rebound.
Other study limitations include that (1) few enrolled women experienced viral rebound; (2) most women had complex treatment histories, and different ART regimens; (3) only two tissues were assessed because they were not asked to consent to lymph node biopsies; and (4) HIV sequences were assessed instead of infectious viruses. The study was not designed to sample multiple tissues. While repeated biopsies of lymph nodes, a recognized site for persistence of infectious HIV during ART, 40,57,58 would likely have been informative, repeated lymph node biopsies would not be acceptable to most human participants.
The current study demonstrates that HIV rebound can arise from multiple cells across multiple tissues, such as the uterine cervix. Implications for ongoing cure strategies suggest that tissue sources other than blood should be evaluated for clearance of infectious variants before suspension of ART, and that sampling of the uterine cervix could serve as a tissue that is accessible and can be repeatedly sampled as a relevant HIV reservoir.
Footnotes
Acknowledgments
This work was supported by the National Institutes of Allergy and Infectious Disease (NIAID) at the National Institutes of Health [PO1 HD 40540-01] (to RWC), R56AI091550 (to LMF); R01 AI071212 (to LMF); R01 AI091550 (to LMF); CFAR STD/AIDS Fellowship AI 07140 (to MEB); and by the University of Washington Center for AIDS Research (CFAR) [P30 AI027757].
Author Contributions
Marta E. Bull designed and directed the studies and was primary author for this original article. Jennifer McKernan-Mullin performed the laboratory assays and sequencing for the study. Sheila Styrchak performed the laboratory assays and sequencing for the study. Kelli Kraft performed the laboratory assays and sequencing for the study. Jane Hitti recruited the patients for the clinical cohort and provided guidance on the study and article. Susan Cohn recruited the patients for the clinical cohort and provided guidance on the study and article. Kenneth Tapia performed the statistical analysis for the study. Wenjie Deng supported the statistical analysis for the study, and assisted in automating the nearest-neighbor permutation analysis. Sarah Holte provided oversight of the statistical analysis for the study. James I. Mullins provided guidance on the phylogenetic analysis of the study and provided input on the article. Robert W. Coombs provided oversight of the laboratory that performed the HIV RNA quantification for the study. Lisa M. Frenkel is principal investigator of the study who designed and directed the studies and authored the original article.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.