
Abstract
We hypothesized that PD-1expressed by regulatory T cells (Tregs) would be functional and their expression levels may associate with activation status of CD4+ T and CD8+ T cells and the disease progression of HIV-1-infected patients. To prove it, we dynamically examined PD-1 expression levels by Tregs in peripheral blood of HIV-1-infected individuals not receiving antiretroviral therapy. Eighty-one HIV-1-infected individuals not undergoing antiretroviral therapy and 22 HIV-1-seronegative donors were enrolled in our study. Tregs were defined as CD4+CD25+CD127lo/− by flow cytometry. Expression of PD-1 and the activation markers CD38, HLA-DR, and Ki67 by Tregs and CD4+ T and CD8+ T cells was also determined by flow cytometry. TGF-β and IL-10 were measured to evaluate the suppressive function of Tregs. In all Tregs, the proportion of PD-1+ Tregs observed in HIV-1-infected persons was significantly greater than that seen in HIV-1-seronegative donors, and correlated with the activation of Tregs and CD4+ T and CD8+ T cells. This increased proportion of Tregs was also statistically associated with the disease progression. Blockade of PD-1/PD-L1 pathway with anti-PD-L1 mAb profoundly increased the level of intracellular TGF-β and IL-10 in CD4+CD25+CD127lo/− Tregs. Our data not only support that PD-1 plays a critical role to predict the activation status of cellular immunity and disease progression during HIV-1 infection but also indicate that blockade of PD-1/PD-L1 pathway represents a novel therapeutic approach to AIDS.
Introduction
Immune deficiency due to chronic HIV-1 infection is usually described as the loss of CD4+ T cells and dysfunction of effector CD8+ T cells that have been suggested not only to control HIV proliferation but also to result in immune exhaustion by killing virus-infected CD4+ T cells. 1,2 Therefore, it is very critical to maintain a balance of immune response and tolerance during HIV-1 infection. Regulatory T cells (Tregs) are regarded as one of the regulators that contribute to maintain self-tolerance and protect the body from autoimmunity. 3,4 In chronic HIV-1-infected subjects not undergoing antiretroviral therapy, the proportion of Tregs has been shown to increase and correlate with disease progression. 5 –7 Further studies showed that depletion of Tregs can promote the activation of CD4+ and CD8+ T cells. 8 –10 Collectively, evidence supports that Tregs exert immune regulatory functions by inhibiting CD4+ and CD8+ T cell activation.
Although the precise mechanism by which Tregs restrain immune activation is not completely understood, accumulating data indicate Tregs can perform suppressive functions through cell-cell interactions and/or the production of immunosuppressive cytokines such as TGF-β and IL-10. Naive T cells undergoing continuous antigenic stimulation have been shown to differentiate into TGF-β and IL-10, producing CD4+CD25+ Tregs that exclusively proliferate upon activation. 11 –16 Furthermore, CD4+CD25+ Tregs significantly increased in peripheral blood and were able to respond to HIV p24 antigen by secreting TGF-β and IL-10 17 ; similar results were found by Aandahl et al. 18 These observations suggest that Tregs exert their suppressive activity by augmenting their activation and production of immunosuppressive cytokines, including TGF-β and IL-10. However, in HIV-1 individuals, the regulation of this response, in particular, factors that are involved in maintaining homeostasis to prevent overactivation of Tregs, is still unclear. These negative regulatory factors may include CTLA-4 and PD-1, for example.
PD-1 and its ligand (PD-1L), a member of B7:CD28 family, have been demonstrated to play a critical role in viral infection. 19,20 Previous studies suggest that PD-1 expression is significantly enhanced on CD4+ and CD8+ T cells in peripheral blood of HIV-1-infected patients compared to healthy controls, and that expression of PD-1 by Tregs is statistically correlated with disease progression. 21,22 Moreover, further studies also showed that HIV-specific CD4+ and CD8+ T cells with high PD-1 expression were more easy to induce exhaustion and dysfunction, 23 –25 and that blockade of the PD-1/PDL pathway could restore the function and proliferation of CD4+ and CD8+ T cells. 26,27 While these data support the hypothesis that PD-1 is an important regulator in controlling effector T cell responses during HIV infection, it is unclear whether PD-1 is expressed by Tregs and if it plays a role in regulating the activation and function of Tregs in HIV-1-infected persons.
In this study, we hypothesize that PD-1 is an activation-induced inhibitor expressed by Tregs that functions to indirectly upregulate CD4+ and CD8+ T cell activation by a downregulation of Treg suppressive function during HIV-1 infection. To test this, we analyzed PD-1 expression levels by Tregs in a cohort of 81 HIV-1-infected patients and then correlated PD-1 expression dynamics with Treg and CD4+ and CD8+ T cell activation.
Materials and Methods
Subjects
Eighty-one HIV-1-positive individuals not undergoing antiretroviral therapy (ART) and 22 HIV-1-seronegative donors were recruited from CDC in Shanghai city and Yuncheng city of Shanxi Province in China, as previously reported. 25,28 All the donors enrolled in this study gave written informed consent as approved by the local Institutional Review Board.
Collection of peripheral blood mononuclear cells, and sorting of CD4+CD25+CD127lo/− Tregs and peripheral dendritic cells
Peripheral blood mononuclear cells (PBMCs) were isolated from freshly heparinized blood by Ficoll-Hypaque (Amersham Biosciences, Piscataway, NJ) density gradient centrifugation. CD4+CD25+CD127lo/− Tregs were purified by using the CD4+CD25+CD127dim/− Regulatory T Cell Isolation Kit II (Miltenyi Biotec, Bergisch-Gladbach, Germany) and peripheral Dendritic Cells (pDCs) were sorted by CD1c (BDCA-1)+ Dendritic Cell Isolation Kit (Miltenyi Biotec). CD4+CD25+CD127lo/− Treg and pDC purity exceeded 90%, as shown by flow cytometric analysis.
Antibodies and reagents
The following directly conjugated monoclonal antibodies were used: CD3-Pacific Blue (clone UCHT1), CD4-PerCP (clone L200), CD25-FITC or CD25-APC (clone M-A251), CD127-PE (clone HIL-7R-M21), CD279-APC (clone MIH4), Ki67-FITC (clone B56), and IL-10-APC (clone JES3-19F1) and with their respective isotype control antibodies were purchased from BD Biosciences, and HLA-DR-ECD (clone Immu-357) was purchased from Beckman Counter; TGF-β-PE (clone 27232) with its isotype control antibody was obtained from R&D Systems. In addition, anti-PD-L1 blocking Ab (clone MIH1) was from eBioscience.
Surface and intracellular staining
mAbs to surface molecules, including anti-CD3-Pacific Blue, anti-CD4-PerCP, anti-25-FITC, anti-127-PE, anti-38-PE-Cy7, anti-HLA-DR-ECD, anti-PD-1-APC, or the corresponding isotype control anti-IgG1-APC, were used to stain PBMCs for measuring PD-1, CD38, HLA-DR expression on CD4+CD25+CD127lo/− Tregs, and CD4+ and CD8+ T cells (defined as CD3+CD4−) at 4°C for 30 min. For intracellular staining, PBMCs were further fixed and permeabilized with Cytofix/Cytoperm solution (BD Biosciences) at 4°C for 20 min, washed with Perm/Wash Buffer (BD Biosciences), and intracellularly stained with anti-Ki67-FITC to determine the level of Ki67 on CD4+CD25+CD127lo/− Tregs, and CD4+ and CD8+ T cell population at 4°C for 30 min in the dark. After rewashing with Perm/Wash Buffer (BD Biosciences), PBMCs were fixed in 1% paraformaldehyde, and stained cells were acquired with a FACSCanto flow cytometer and analyzed by FACSDiva software (BD).
Cell stimulation and flow-based intracellular TGF-beta detection
Freshly isolated CD4+CD25+CD127lo/− Tregs (5 × 10E 4 ) and pDCs were cocultured at a 1:1 cell ratio and stimulated with anti-CD3 (1 μg/mL; BD Biosciences) and anti-CD28 (2 μg/mL; BD Biosciences), or PMA (50 ng/mL; Sigma) and ionomycin (1 μg/mL; Sigma) in RPMI 1640 containing 10% human AB serum and either purified anti-PD-L1 (10 μg/mL) or isotype control antibodies in the presence of IL-2 (10 ng/mL; Sigma). GolgiStop (BD Pharmingen) was added to the cocultured cells after 10 days of stimulation, and on day 10.5, cells were stained with surface antibodies and intracellular IL-10-APC and TGF-β-PE antibodies according to the abovementioned intracellular staining protocols.
Plasma HIV-1 RNA testing
Plasma RNA was purified from 1 mL of venous blood derived from the studied subjects. Five microliters RNA and 90 μL primers mixture were added into an Eppendorf tube, followed by incubation at 65°C for 2 min and 41°C for 2 min. Five microliters enzyme mix and 45 μL diluent reagents were then added, mixed thoroughly, and centrifuged for 2 min. The plasma HIV-1 RNA was quantified in a NucliSens EasyQ instrument.
Statistical analysis
All data were analyzed by Wilcoxon matched pairs test and Mann–Whitney test of GraphPad Prism, Version 5.0 software, to determine the significance between different groups. Values are expressed as mean ± standard deviation. Correlations between variables were evaluated using the Spearman rank correlation test. p-Values <.05 were considered statistically significant.
Results
Patient characteristics
Demographic information and CD4 T cell count were obtained at the time of enrolment and blood draw. HIV RNA level was determined from plasma. The gender, average of age, HIV RNA copies, log HIV RNA copies, and CD4+ T cell counts are shown in Table 1 and Supplementary Table S1.
Characteristics of HIV-1-Infected Patients
All patients were divided into three groups depending on the HIV RNA copies.
VL, viral load.
PD-1 expression is elevated on Tregs following HIV-1 infection
In our study, Tregs were defined by a population of CD4+CD25+CD127lo/− cells. We found that the HIV-1-infected patients had significantly higher percentage of PD-1-expressing Tregs as viral load (VL) increases [VL <2,000 copies/mL (mean 2.49% ± 1.28%); VL 2,000–20,000 copies/mL (mean 3.83% ± 2.74%); VL >20,000 copies/mL (mean 5.12% ± 2.85%); and HIV-1-seronegative donors (mean 1.64% ± 1.12%, all p < .05)] (Fig. 1A, B). Furthermore, this enhanced level of PD-1 was also found in patient cohorts stratified by CD4+ T cell count with mean at 2.82% ± 2.40% for group with CD4 > 500, 4.37% ± 2.97% for group with CD4 at 200–500, and 5.75% ± 3.21% for group with CD4 < 200 compared to healthy subjects (all p < .01; Fig. 1C). The results reveal that PD-1 expression was elevated on Tregs of HIV-1-infected subjects without antiretroviral therapy.

PD-1 expression on CD4+CD25+CD127lo/− Tregs is higher at different stages of HIV-1-infected patients than in HIV-1-seronegative subjects (healthy subjects) and correlate with disease progression.
Since VL and CD4+ T cell count are widely accepted predictors of viral persistency in untreated HIV-1-infected individuals, 29 –31 we evaluated whether PD-1 expression on CD4+CD25+CD127lo/− Tregs correlated with these two disease progression markers. We found that the frequency of PD-1-expressing Tregs was positively correlated with VL (r = 0.359, p = .001; Fig. 1D) and negatively correlated with CD4+ T cell count (r = −0.4414, p < .0001; Fig. 1E). These data demonstrate that the frequency of PD-1-expressing Tregs of total may also serve as an indicator of disease progression in chronic HIV-1 infection.
Increased frequency of PD-1-expressing Tregs is also associated with activation of Treg and CD4+ and CD8+ T cell populations
Previously published articles have demonstrated that CD38, HLA-DR, and Ki67 were all important indicators of T cell activation during viral infections. 32 –37 We found expression levels of CD38, HLA-DR, and Ki67 were increased on Tregs in HIV-1-infected individuals compared to healthy donors (data not shown). Furthermore, the frequency of PD-1-expressing cells was positively correlated with the frequency of CD38-, HLA-DR-, and Ki67-positive Tregs (CD38: r = 0.4256, p < .0001; HLA-DR: r = 0.501, p < .0001; and Ki67: r = 0.4456, p < .0001; Fig. 2A–C). The results suggested that PD-1 might be associated with activation of Treg during HIV-1 infection.

PD-1 expression on CD4+CD25+CD127lo/−Tregs is associated with immune activation.
As the frequency of PD-1+ Treg cells increased with HIV-1 disease progression in this study, we examined whether PD-1 expression by Tregs was associated with inhibition of reactive functions of effector T cells by the evidence of the decreased activation of CD4+ and CD8+ T cells. We found Treg PD-1 expression was positively associated with levels of CD38, HLA-DR, and Ki67 on CD4+ T cells (r = 0.2272, p < .0414 for CD38, r = 0.5559, p < .0001 for HLA-DR, and r = 0.4465, p < .0001 for Ki67; Fig. 2D–F), but also positively correlated with CD38 and HLA-DR expressions on CD8+ T cells (r = 0.5036, p < .0001 for CD38 and r = 0.5037, p < .0014 for HLA-DR; Fig. 2G, H). There was no correlation between PD-1 levels of Tregs and Ki67 levels of CD8+ T cells (r = 0.0078, p = .9451; Fig. 2I). Collectively, these evidences indicated that the homeostasis of PD-1 on Tregs was positively associated with activation of CD4+ and CD8+ T cell populations.
PD-1 levels of Tregs are associated with exhaustion of CD4+ and CD8+ T cells
Since PD-1 was reported to be a predictor of effector T cell exhaustion, and positively correlated with activation of CD4+ and CD8+ T cells during HIV-1 infection, 21,38 we correlated PD-1 elevation on Tregs with exhaustion of CD4+ or CD8+ T cells, as determined by expression of PD-1. We found that the frequency of PD-1-expressing Tregs positively correlated with the frequency of PD-1-expressing CD4+ and CD8+ T cells (r = 0.7365, p < .0001 and r = 0.6381, p < .0001 respectively; Fig. 3A, B). Our analysis implied that the elevation of PD-1 on Tregs positively correlated with exhaustion of CD4+ and CD8+ T cells.

PD-1 expression on CD4+CD25+CD127lo/−Tregs is associated with immune exhaustion.
Blockade of PD-1/PD-L1 pathway enhances the function of Tregs
Although emerging data have demonstrated that blockade of PD-1/PD-L1 pathway can restore proliferation and reactive function of CD4+ and CD8+ T cells, 19,39 there is little evidence for the effect of Tregs in blocking PD-1/PD-L1 pathway in vitro. We assessed whether interrupting the ligation of PD-1/PD-L1 enhanced functional properties of Tregs by monitoring the production of TGF-β and IL-10 by Tregs from HIV-1-infected persons. We sorted CD4+CD25+CD127lo/− Tregs and dendritic cells (define) from chronic HIV-1-infected patients, and cultured them at a ratio of 1:1 before stimulation with PMA/ionomycin or anti-CD3/CD28 antibodies in the presence or absence of anti-PD-L1 antibodies for 10 days. We found that the production of TGF-β and IL-10 was both increased after blockade of PD-1/PD-L1 pathway (Fig. 4A, B). Our results suggested that blockade of PD-1/PD-L1 signal pathway can enhance the secretory function of Tregs.
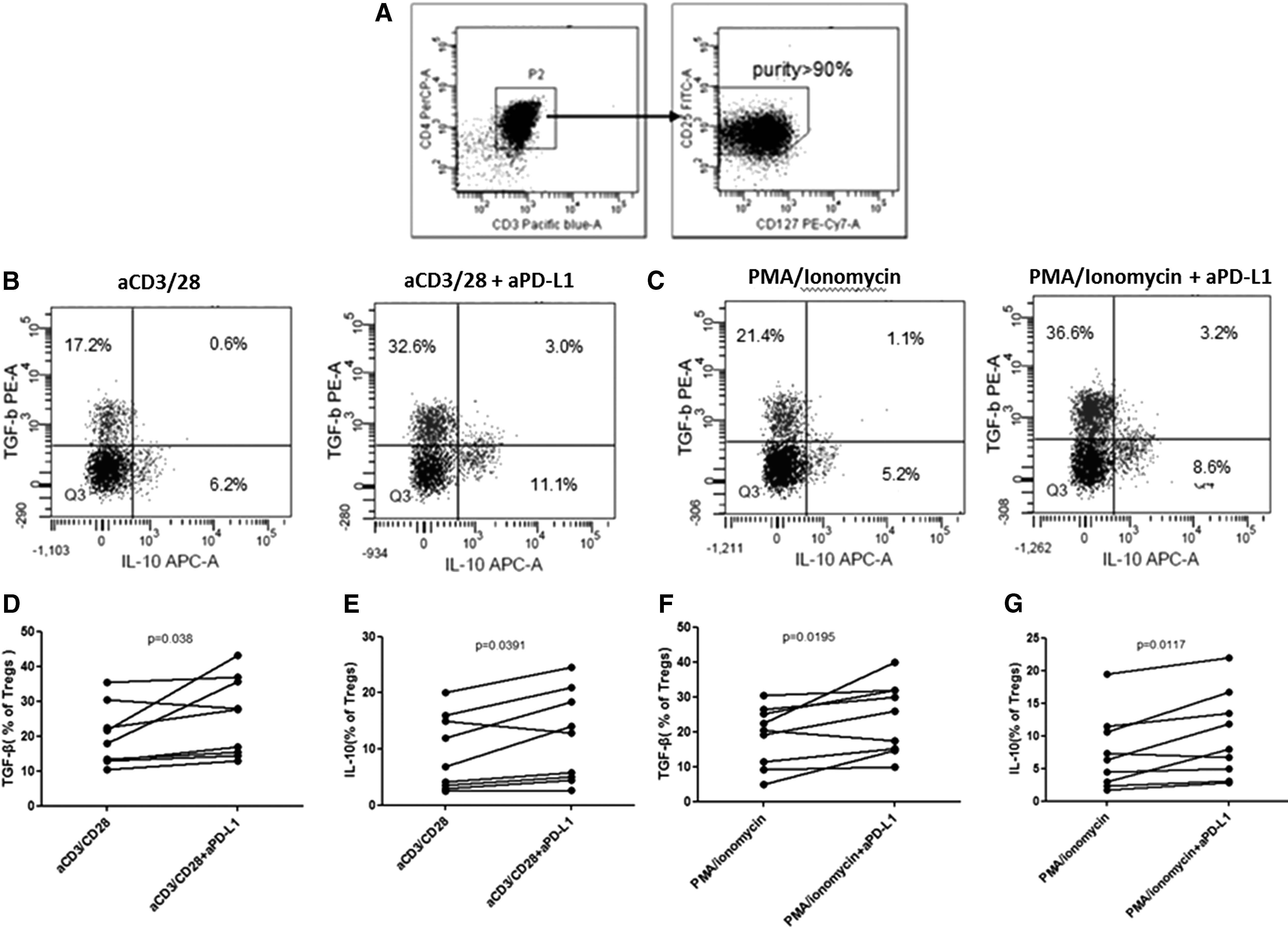
Blocking PD-1/PD-L1 pathway with anti-PD-L1 enhances levels of TGF-β and IL-10 on CD4+CD25+CD127lo/−Tregs.
Discussion
It is well known that Tregs play a critical role in preventing autoimmune disease by the inhibition of antigen-reactive T cell activation. 3,4 However, in viral immunity, overactive Tregs could lead to persistent survival of virus due to suppressed effector response. As a member of the costimulatory CD28/B7 family, PD-1 has been demonstrated to act as a central mediator for the activation and proliferation of HIV-specific CD4+ or CD8+ T cells. 23,40 The role of PD-1 in regulating the activation and function of Tregs during HIV infection is less clear. We hypothesized that PD-1 plays an important role in the activation of Tregs in HIV infection. This study showed an increased frequency of PD-1-expressing Tregs in HIV-1-infected patients compared to HIV-1-seronegative donors. In addition, we found the frequency of PD-1-expressing Tregs to be positively correlated with HIV disease progression (as evidenced by a significant association with increased VL and reduced CD4+ T cell counts), but also activation of Tregs and CD4+ and CD8+ T cell populations (as evidenced by a positive correlation with increasing CD38, HLA-DR, and Ki67). In vitro stimulation data also suggest that blockade of PD-1/PD-L1 pathway could enhance TGF-β and IL-10 production by Tregs. Taken together, our findings indicate that PD-1 is a central regulator of Tregs in HIV-1-infected patients and are in agreement with previous observations that PD-1 expression by CD4+CD25+Foxp3+ Tregs is important for determining the persistence of viral infection and pathogenesis. 41
Accumulating evidence suggests that in HIV-1-positive individuals, PD-1 expressing is increased by CD4+ and CD8+ T cells compared to healthy donors, 23 –26 and that upregulation of PD-1 was statistically associated with activation of these two cell subsets. 22,40 Our results support this. Another study showed that Tregs could suppress the function and proliferation of CD8+ T cells from HIV-infected patients. 42 We hypothesized that expression of PD-1 by Tregs modulated activation of CD4+ and CD8+ T cells in HIV-1-infected patients. Our data showed that the frequency of PD-1-expressing Tregs was associated with frequency of CD38-expressing and HLA-DR-expressing CD4+ and CD8 + T cells, as well as Ki67-expressing CD4+ T cells. Accordingly, our results, at least partly, support our hypothesis that PD-1 expression by Tregs is not only an indicator of Treg activation but also a predictor of CD4+ and CD8+ T cell activation during chronic HIV-1 infection.
Because the previous data have indicated that activation can induce PD-1 upregulation on reactive T cells, 43,44 and our results also showed that Treg PD-1 expression correlated with activation, we then hypothesize that exhaustion of Tregs may increase exhaustion of these two populations. In this study, we demonstrated that PD-1 on Tregs positively correlated with PD-1 on CD4+ and CD8+ T cells. This study indicated that upregulation of PD-1 on Tregs might be associated with exhaustion of CD4+ and CD8+ T cells and PD-1 should be a notable indicator of T cell exhaustion in HIV-1-infected subjects.
The use of CD4, CD25, and CD127 to define Tregs may be perceived as a limitation of this study. While many studies suggest that coexpression of CD4, CD25, and Foxp3 was the best and most reliable Treg phenotype, 45 –47 it is necessary to fix and permeabilize cells before performing intracellular staining for Foxp3, which can negatively impact cell surface marker expression and prohibit the isolation of viable Tregs for functional studies and ex vivo expansion. Recent studies have revealed that the positive expression of Foxp3 and low, or negative, expression of CD127 were highly correlated within CD4+CD25+ population, and suggested CD4+CD25+CD127lo/− T cells as an appropriate substitute of CD4+CD25+Foxp3+ T cells in healthy individuals. 48 –52 Considering that some researchers still insist on the inclusion of Foxp3+ when phenotyping Tregs in HIV-infected persons, 53 we first checked the correlation between Foxp3+ and CD127lo/− expression by CD4+CD25+ T cells and found that CD127lo/− was strongly correlated with Foxp3 positivity in HIV-1-infected chronic subjects not undergoing antiretroviral therapy (Supplementary Fig. S1). Thus, we chose to define Tregs as CD4+CD25+CD127lo/− to sort pure Tregs for functional assays.
Data from several groups have demonstrated that blocking PD-1/PD-L1 pathway could significantly increase cytokine production, including TNF-α, IFN-γ, and IL-2 of CD4+ or CD8+ T cells during viral infection. 54 –56 Franceschini et al. also found that blockade of the interaction between PD-1 and PD-L1 enhanced the in vitro expansion and function of CD4+CD25+ Tregs isolated from the livers of chronic HCV-infected patients. 41 Furthermore, other researchers described that CD4+CD25+ Tregs could respond to p24 antigen by secreting TGF-β and IL-10, although the suppressive function of Tregs was not dependent on these cytokines. 17 Basing on these previously described evidences, we want to know whether PD-1/PD-L1 signals correlate with the production of TGF-β and IL-10. In this study, we found that blockade of PD-1/PD-L1 complex just partly increased levels of these two cytokines in CD4+CD25+CD127lo/− Tregs. These data further demonstrated that PD-1 was a key factor of Treg exhaustion, and more studies are expected to clarify the exact mechanism. On the other hand, our current data suggested that the function of PD-1 on Tregs is same with it on non-Tregs. It will be very interesting to investigate whether PD-1 on Tregs has other functions and the functions of Tregs in tissue, because we only investigated the circulation Tregs in this study.
The existence of the latent reservoir of HIV-1 virus is one of the biggest obstacles for treatment of HIV-1 infection. Previous studies show that during HIV infection, elevated expression of indoleamine 2,3-dioxygenase (IDO) skews CD4+ T cell differentiation into Treg instead of T-helper 17 cells 57 ; and plasma IDO activity is associated with the size of the HIV reservoir in patients receiving ART. 58 In addition, in contrast to progressors, HIV controllers have been shown to exhibit Treg levels similar or even lower than healthy donors. 59 So Treg is believed to help in the establishment of a latent reservoir of HIV-1 virus in multiple ways. 60 Target Treg is a possible way to reduce latent reservoir. In addition, VA Evans etc. showed that PD-1 expressed on CD4+ T cells contribute to the establishment and maintenance of HIV-1 latency. 61 Furthermore, Teigler et al. showed that MCP-1, MIP-3β, sTNFR-II, and IL-10 levels before ART were associated with the establishment and persistence of the viral reservoir during ART. 62 Therefore, it would be very interesting to investigate whether blocking PD-1 and IL-10 signal could reduce latent reservoir in future study. In addition, the correlation of Treg frequency and exhaustion level with circulating IL-10 also will be investigated in our future study.
Overall, our data indicate that PD-1 expression on Tregs is preferentially elevated as the increase of VL during HIV infection. Furthermore, our study implied that the level of PD-1 on Tregs may indirectly impact the function of CD4+ and CD8+ T cells and blocking PD-1/PD-L1 pathway in vitro can enhance the cytokine production of Tregs. These results shed light on vaccine research and potential therapeutic strategy for HIV-infected patients. The latter observation could suggest more functional studies to testify the exact mechanism of how Tregs balance the immune response and how multisignals, including PD-1, CD95, and CTLA-4, cooperate to regulate the function of regulatory T cells, just like as mentioned above.
Footnotes
Acknowledgments
We thank all HIV-infected individuals and healthy volunteers in this study for their cooperation. This work was supported by Natural Science Foundation of China (project no. 81260463) and by the National Grand Program on Key Infectious Disease Control from Ministry of Sciences and Technology and Ministry of Health.
Author Disclosure Statement
The authors declare no financial conflicts of interest.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.