
Abstract
Most HIV-1 transmissions occur at mucosae and involve exposure to semen. Semen contains immunomodulatory factors, which inhibit anti-HIV-1 natural killer cell and T cell responses. We demonstrate high concentrations (1:2 dilution) of seminal plasma (SP) inhibit monocyte phagocytosis and anti-HIV-1 Fc-dependent functions of both neutrophils and monocytes. In addition, slightly lower SP concentrations (1:2–1:10 dilutions) inhibit granulocyte phagocytosis and oxidative burst of both monocytes and granulocytes. These observations may have implications for HIV-1 infectivity after mucosal exposure.
T
Most new HIV-1 infections are established after sexual exposure at mucosal membranes, often involving semen. Semen is a complex solution containing factors that can drive or inhibit HIV-1 infection. 6 Semen contains proinflammatory components, which can serve to recruit leukocytes, including macrophages, to the female reproductive tract postcoitus. 7 Semen also contains immunosuppressive factors that inhibit immune responses in vitro. 8 –10 We previously showed T cell and NK cell anti-HIV-1 functions are inhibited by seminal plasma (SP), 9 but have not analyzed the effect of SP on neutrophils and monocytes.
We first tested the effect of SP on phagocytosis of opsonized heat-inactivated fluorescein isothiocyanate-labeled Escherichia coli (PHAGOTEST™ kit; Glycotope Biotechnology). Whole blood (50 μL) was mixed with 50 μL of an HIV-1-uninfected SP pool (BioIVT) diluted in RF10 or RF10 alone. E. coli (20 μL of 2 × 10 9 /mL) was added and incubated for 10 min at 37°C. Samples were then placed on ice and quenched to prevent detection of surface-bound bacteria. Finally, samples were treated with 500 μL of ACCUTASE (StemCell Technologies) for 10 min to promote detachment before analysis by flow cytometry, using forward and side scatter to identify monocytes and granulocytes. Robust phagocytosis of opsonized bacteria was detected in the absence of SP by both leukocyte populations, with between 83.0%–94.0% monocytes and 68.9%–96.5% granulocytes containing FITC-labeled bacteria (Fig. 1A). SP inhibited phagocytosis activity of both leukocyte populations in a dose-dependent manner. Significant decreases in granulocyte-mediated phagocytosis were noted in the presence of 1:2, 1:5, and 1:10 SP dilutions. A significant decrease in monocyte-mediated phagocytosis was only observed in the presence of a 1:2 SP dilution.
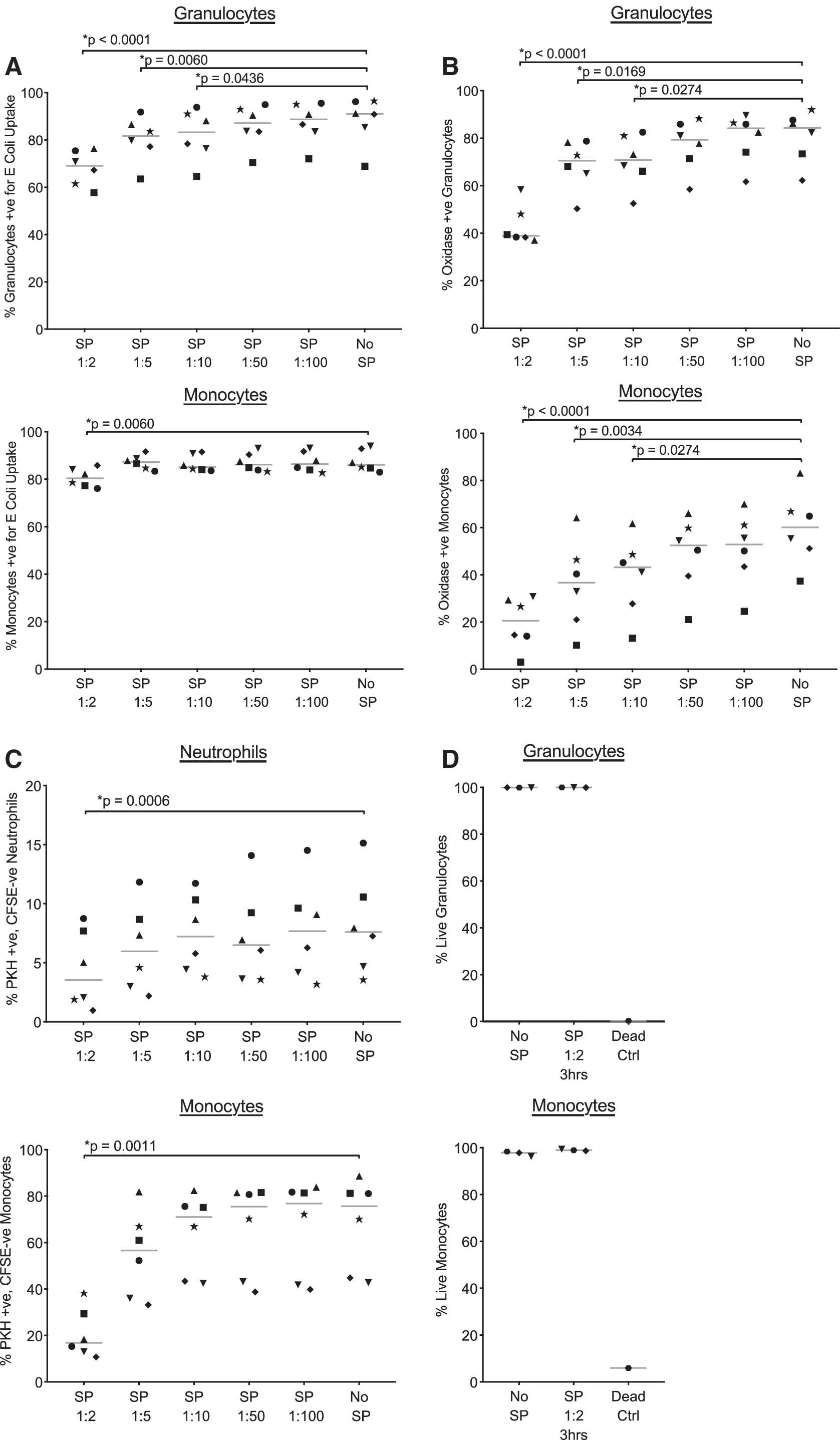
Effects of SP on the functions of phagocytes.
Oxidative burst functions of phagocytes are critical to the killing of engulfed pathogens by granulocytes and monocytes. We next tested the effect of SP on oxidative burst, as measured by conversion of a fluorogenic substrate dihydrorhodamine 123 to rhodamine 123, of granulocytes and monocytes after phagocytosis of E. coli (PHAGOBURST™ kit; Glycotope Biotechnology). Samples were prepared as previously, except that after the 10 min incubation at 37°C, 20 μL of substrate was added to tubes, which were then incubated for another 10 min at 37°C. Oxidative burst activity was detected in 37.4%–83.1% monocytes and 62.2%–92.0% granulocytes in the absence of SP (Fig. 1B). The addition of 1:2–1:10 dilutions of SP significantly inhibited oxidative burst activity for both granulocytes and monocytes.
Anti-HIV-1 antibody-mediated functions of phagocytic cells may be important effector mechanisms of HIV-1 prevention, 11 but the effect of SP on these functions has not been studied. The rapid fluorescent ADCC (RFADCC) assay detects anti-HIV-1 antibody-dependent functions of effector cells using a labeled HIV-1 envelope-coated CD4+ T cell line as targets. 4,5 Although envelope-coated target cells do not recapture the trimeric closed conformation of envelope observed on HIV-1-infected cells, antibodies targeting CD4-bound envelope are common in plasma IgG from HIV-1-infected donors. 12 As such, utilization of gp120-coated target cells allows detection of robust anti-HIV-1 antibody-dependent responses using plasma IgG. Peripheral blood mononuclear cells or enriched neutrophils (EasySep™ Direct Human Neutrophil Isolation Kit; StemCell Technologies) were coincubated for 1 h with CFSE and PKH26 labeled HIV-1BaL gp120-coated CEM.NKr-CCR5 cells at a 5:1 effector to target cell ratio. Incubations were conducted in the presence of 100 μg/mL of pooled HIV-1 immunoglobulin (HIVIG), as the source of anti-HIV-1 antibody, and in the presence or absence of SP. The HIV-1 specificity of monocyte and neutrophil RFADCC responses has been previously confirmed. 4,5 Purified neutrophils, or monocytes within PBMC, that took up PKH26+ CEM.NKr-CCR5 membrane were quantified by flow cytometry. SP had a modest negative impact on anti-HIV-1 RFADCC, significantly inhibiting the response at a 1:2 dilution only (Fig. 1C).
High concentrations of SP were employed to assess modulation of leukocyte responses. Importantly, SP is known to have cytotoxic effects after cell culture. 13 To determine if cytotoxicity explained inhibition of leukocytes, we incubated 50 μL of blood from three donors with a 1:2 dilution of SP for 3 h. Next, blood was washed twice with phosphate buffered saline (PBS), to remove serum proteins, and stained with LIVE/DEAD™ far red stain (Invitrogen). Lastly, samples were treated with lysis buffer (BD Biosciences), washed in PBS and analyzed by flow cytometry. As shown in Figure 1D, granulocytes and monocytes within blood treated with SP exhibited similar viability as those not exposed to SP. To confirm the viability dye identified dead cells, blood from one donor was fixed with 1% formaldehyde for 10 min before washing and staining. Cell death of monocytes and granulocytes were detected in this sample (Fig. 1D).
The capacity of SP to influence HIV-1 susceptibility is likely multifactorial, involving both induction of mucosal inflammation and alterations in immune function. We previously showed SP at very low concentrations (i.e., 1:100–1:1,000 dilutions) significantly inhibited NK cell and T cell functions. 9 In this study, we show SP inhibits monocyte phagocytosis and monocyte and neutrophil RFADCC responses, but relatively high concentrations (i.e., 1:2 dilution) of SP are required for this effect. In the absence of SP exposure to underlying innate cells through microtears of the mucosa, this concentration may not be achieved. Interestingly, intermediate concentrations of SP (i.e., 1:5 and 1:10) were sufficient to inhibit granulocyte phagocytosis and monocyte and granulocyte oxidative burst responses. These data are consistent with previous literature demonstrating SP to inhibit monocyte and neutrophil/granulocyte responses. 8,10 The heightened sensitivity of the PHAGOBURST response to inhibition by SP led us to speculate that inhibition of oxidative burst could create a situation whereby SP-exposed phagocytic cells engulf but fail to destroy viral particles. This could increase the likelihood of noninfected virus-harboring cells infecting new target cells through infectious synapses. Future research is required to confirm that SP-mediated inhibition of monocyte and granulocyte function occurs under physiologically relevant conditions. Several studies have identified macrophages and neutrophils in cervicovaginal fluid, 14,15 which would represent mucosal immune cells exposed to the highest concentration of SP. The contribution of these cells to immune function, however, is difficult to determine. Once released from tissue and shed into the vaginal lumen, these cells are likely transitory and not long retained in the reproductive tract. In addition to determining the effects of SP on leukocytes within vaginal fluid, the impact of vaginal fluid itself on SP-mediated immunomodulation should be assessed. Furthermore, determining the degree that SP is diluted by vaginal fluid postcoitus will establish the physiological relevance of SP dilutions that suppress leukocyte function in vitro. Additional work in tissue explants and animal models should evaluate how immunomodulation by SP interacts with other features of semen, such as the capacity to recruit cells to mucosal tissues 7,16 and the presence of amyloid fibrils reported to enhance HIV-1 infectivity, 13,17,18 to shape susceptibility to HIV-1 infection.
Footnotes
Acknowledgments
This study was funded by grant 1124680 from the Australian National Health and Medical Research Council (NHMRC). The authors thank the study participants for donating blood samples and also thank the NIH AIDS Reagent Program for the following reagents: HIVIG, HIV-1BaL gp120, and the CEM.NKr-CCR5 cell line.
Author Disclosure Statement
The authors have no conflicts of interest to declare.