
Abstract
Persistence of replication-competent viral reservoirs during infection remains a barrier to HIV cure, despite the ability of combination antiretroviral therapy (cART) to effectively suppress viral replication. Simian-tropic HIV (stHIV) is a minimally chimeric HIV-1 that is comprised of 94% HIV-1 sequence, contains HIV-1 drug and immunologic targets, and is capable of replicating to high levels and causing authentic HIV-like pathogenesis leading to clinical AIDS in pigtail macaques. Suppression of stHIV replication by cART provides a model for study of viral reservoirs and HIV-specific intervention strategies targeting them. Accurate measurement of reservoir size is crucial for evaluating the effect of any such intervention strategies. Although there are a variety of assays that allow for indirect monitoring of viral reservoir size ex vivo, they each quantify a different aspect of viral reservoirs, and are characterized by conceptual and/or technical limitations. Measurement of viral protein in ex vivo cell culture assays captures the immunologically relevant viral-antigen producing component of the reservoir. This study demonstrates the utility of an ultrasensitive digital HIV Gag p24 immunoassay, which enabled earlier, and more sensitive detection of viral protein in culture supernatants from stimulated CD4+ T cells from stHIV-infected pigtail macaques receiving cART compared with conventional enzyme-linked immunosorbent assay. Protein measurements were highly correlated with cell-free stHIV RNA, as measured by quantitative reverse transcription polymerase chain reaction. This ultrasensitive p24 assay can be used to complement other reservoir measurement tools to assess ongoing replication and reactivation of infectious virus from reservoirs in stHIV-infected pigtail macaques.
Introduction
Although current regimens of combination antiretroviral therapy (cART) are effective at suppressing HIV-1 replication in infected humans to levels below the limit of quantification for standard clinical assays, treatment is unable to cure infection. During cART, the virus persists in the form of “viral reservoirs,” sources of residual virus from which active replication may be reinitiated upon treatment interruption. 1 Evaluation of any candidate AIDS virus “cure” intervention will require accurate quantification of residual virus, including inducible latent virus during treated infection. Laboratory assays measuring reservoir size provide an alternative to using in vivo cART interruption, with time to viral rebound as a presumptive measure of viral reservoir size. At present, there are a plethora of assays used to quantify residual virus material, including assays for directly measured and induced levels of viral DNA, RNA, and protein, along with culture-based measures of replication-competent virus. 2
Of the many parameters that may be used to assess the viral reservoir, measurement of induced viral protein in ex vivo cell culture assays captures the immunologically relevant viral-antigen producing component of the reservoir. Detection of HIV p24CA protein in virus induction assays has been limited to the picomolar level of detection of a capture enzyme-linked immunosorbent assay (ELISA), which is not sensitive enough to detect the low levels of p24 produced in many reservoir reactivation studies without extended culture. The recent development of single molecule array (Simoa) technology has resulted in ultrasensitive protein detection. 3 Previous work has shown the utility of this assay for detecting femtomolar concentrations of HIV p24 in plasma samples from seronegative HIV+ individuals and viral protein released from ex vivo stimulated CD4+ T cells isolated from ART suppressed HIV+ individuals. 4,5
Simoa is a digital bead-based immunoassay platform in which enzyme-linked immunocomplexes are formed on paramagnetic microbeads, which are then individually dispensed into 40-femtoliter microwells. Interaction between the enzyme and its substrate in the wells results in a fluorescent signal that is detected by a charge-coupled device camera. The combination of assay wells small enough to accommodate only a single bead and a digital positive or negative signal for each well result in a significant increase in the signal-to-noise ratio and enhanced sensitivity. 3 Consequently, many Simoa immunoassays have a dynamic range that spans four orders of magnitude and can measure femtomolar concentrations of protein.
Nonhuman primate models of AIDS, particularly those involving the use of effective cART regimens to suppress replication of simian immunodeficiency virus (SIV) and chimeric HIV/SIV viruses, such as simian-HIV (SHIV) and simian-tropic HIV (stHIV), enable studies of viral reservoirs and proof of concept testing of AIDS virus cure interventions. 6,7 Many of the laboratory assays used to measure HIV reservoirs have been adapted for use in SIV and SHIV systems, including Simoa. 8 Suppression of stHIV replication by cART provides a novel model for study of viral reservoirs and HIV-specific intervention strategies targeting them. Accurate measurement of reservoir size is crucial for evaluating the effect of any such intervention strategies. Here, we seek to expand the application of Simoa technology to measurement of viral reservoirs in cART treated stHIV-infected pigtail macaques.
We cultured primary CD4+ T cells from 11 pigtail macaques infected with the stHIV clone A19 (Schmidt F, Keele BF, Del Prete GQ, et al. Submitted) that were on a suppressive cART regimen or were virologic controllers at the time of sampling (plasma viral loads <2–86 RNA copies/mL). Cells were maintained under one of three conditions: stimulated, stimulated with antiretrovirals (ARVs), or unstimulated, with 2–20 × 10 6 cells plated per condition. Stimulated cultures were treated with Concanavalin A (ConA) and interleukin (IL)-2 for 24 h, at which point the ConA was washed out and the cultures were maintained in IL-2. ARV treatment consisted of emtricitabine (FTC, reverse transcriptase inhibitor) and dolutegravir (DTG, integrase strand transfer inhibitor), and was added back to cultures on days 1, 3, and 5. The CD4+ T cells were cultured for 7 days, with sampling of the culture supernatant on days 0 (before stimulation), 1 (after 24 h of stimulation), 3, and 7. stHIV p24 Gag in culture supernatants was quantified using a conventional ELISA (Advanced Biosciences Laboratories; standard curve range of 3.1–100 pg/mL) and by the Quanterix HIV p24 Gag assay (standard curve range of 0.017–40 pg/mL), both used according to the manufacturer's instructions. In addition, stHIV gag RNA in culture supernatants was measured with a quantitative reverse transcription polymerase chain reaction (qRT-PCR) assay. 9 Finally, cell-associated (ca) gag RNA and DNA in samples were measured on days 0 and 7 by qRT-PCR and qPCR, respectively. 10
Of 86 samples (out of 132 from infected macaques) that were stHIV Gag p24 positive by the Simoa immunoassay, only 15 (17.4%) were detected by the conventional p24 ELISA, demonstrating the enhanced sensitivity of the Simoa immunoassay below the LOD of the p24 ELISA (3.1 pg/mL). On comparison of results from the ConA stimulated alone (Fig. 1A, D) versus stimulated + ARV (Fig. 1B, E) conditions, two sources of stHIV p24 Gag in culture supernatants were revealed: (1) virus arising from spreading infection in the culture and (2) induced virus released directly from stimulated latently infected cells. The low level of stHIV p24 Gag (∼1 pg/mL) observed in cultures on day 1 is likely the result of virions released from latently infected cells after stimulation; as expected this virus was nearly unaffected by the presence of ARVs. In addition, there is clear evidence of exponential viral replication over time in stimulated alone cultures. Importantly, CD4+ T cells from three uninfected pigtail macaques were cultured under the same conditions and all time points were negative by every viral protein and RNA quantification method (n = 36, data not shown).
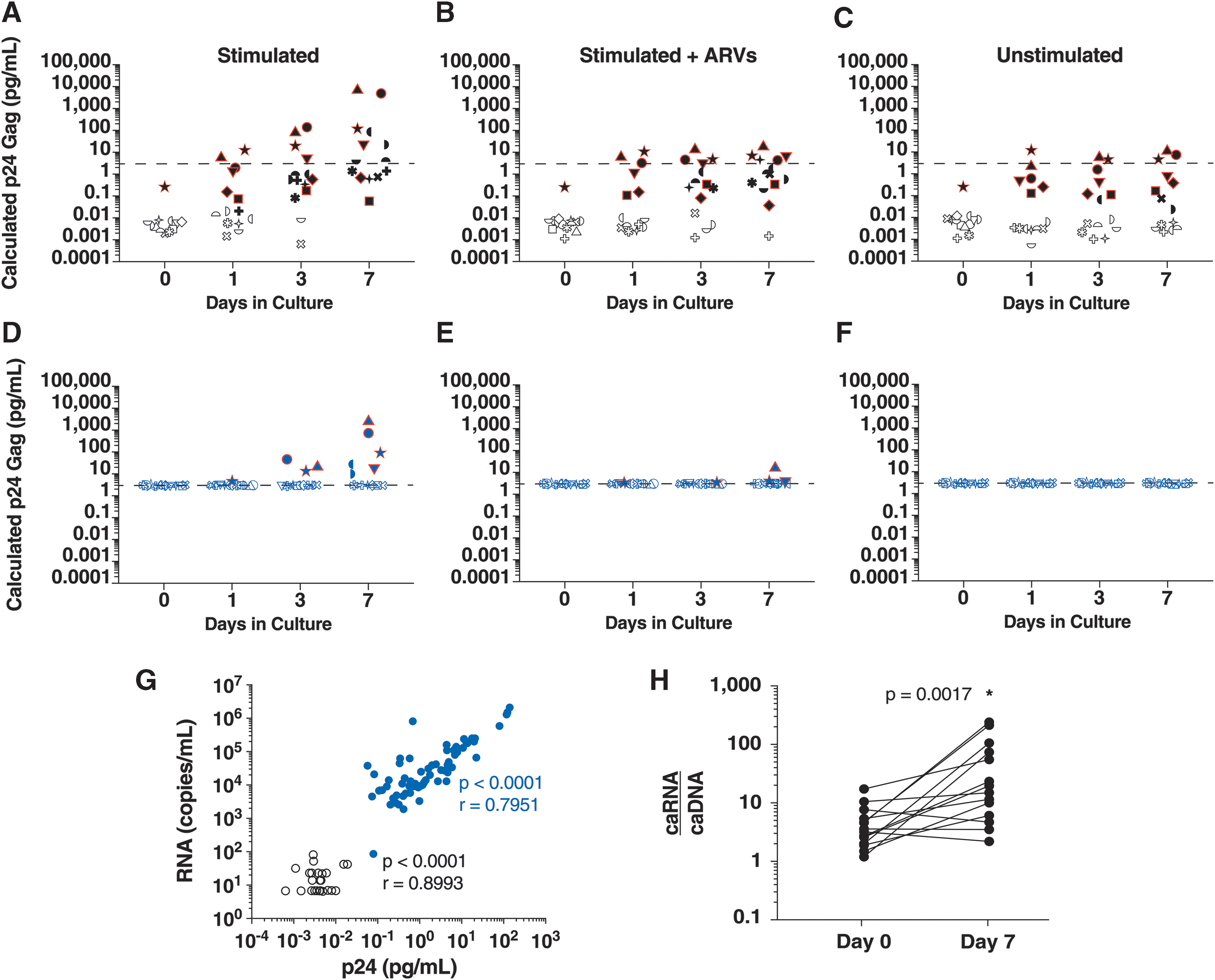
Detection of stHIV ex vivo. stHIV Gag p24 was quantified in culture supernatants by both Simoa
Virus reactivation was also measured by quantification of gag RNA in culture supernatants as well as caRNA and caDNA. A significant correlation was observed between the quantities of stHIV p24 Gag and gag RNA (Fig. 1G), as measured by the ultrasensitive immunoassay and qRT-PCR methods, respectively. The correlation was significant in cultures positive by both methods (blue circles) and when samples that were RNA positive but between the Simoa limits of quantification and detection were included (open black circles). A significant increase in the ratio of caRNA to caDNA from day 0 to 7 was only observed in stimulated cultures (Fig. 1H), consistent with ongoing active viral replication.
This study has shown the application of the Simoa ultrasensitive digital immunoassay technology to detect ex vivo induced p24 Gag in culture supernatants expressed from CD4+ cells from macaques infected with stHIV. The enhanced sensitivity of the Simoa p24 assay compared with the conventional ELISA allows for the distinction between induced virus released directly from stimulated latently infected cells and virus resulting from spreading infection in culture. There is a significant correlation between gag RNA quantified by RT-PCR and p24 Gag quantified by Simoa in culture supernatants, suggesting that stHIV gag RNA quantification and the ultrasensitive p24 Gag immunoassay are complementary tools for the characterization of virus persistence and reactivation in this model. This nonhuman primate model and ultrasensitive immunoassay format should be useful for analyzing the effectiveness of various latency reversing agents, as has been previously been reported using latently infected cells from HIV-1-infected patients, 4 and other HIV-specific reservoir reduction strategies.
Footnotes
Acknowledgments
The authors thank the members of the Nonhuman Primate Research Support Core of the AIDS and Cancer Virus Program: Vicky Coalter, Adam Wiles, Rodney Wiles, Jacob Kiser, and Colleen Boyenton, as well as the nonhuman primate care staff in the Laboratory Animal Sciences Program at Leidos Biomedical Research, Inc., Frederick National Laboratory for Cancer Research. The authors thank the members of the Scientific Publications Graphics and Media department of the National Cancer Institute, Frederick National Laboratory for Cancer Research, for their assistance with figure preparation. All pigtail macaques (Macaca nemestrina) in this study were treated according to a protocol approved by the IACUC of the National Cancer Institute and housed and cared for in accordance with the Association for the Assessment and Accreditation of Laboratory Animal Care (AAALAC) standards in AAALAC-accredited facilities. This study was supported in part with federal funds from National Institutes of Health grant R01AI78788 and from the National Cancer Institute, National Institutes of Health, under contract no. HSN261200800001E. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of any trade names, commercial products, or organizations imply endorsement by the U.S. government.
Author Disclosure Statement
No competing financial interests exist.