
Abstract
Impaired immunity is a common symptom of aging and advanced Human Immunodeficiency Virus type 1 (HIV-1) disease. In both diseases, a decline in lymphocytic function and cellularity leads to ineffective adaptive immune responses to opportunistic infections and vaccinations. Furthermore, despite sustained myeloid cellularity there is a background of chronic immune activation and a decrease in innate immune function in aging. In HIV-1 disease, myeloid cellularity is often more skewed than in normal aging, but similar chronic activation and innate immune dysfunction typically arise. Similarities between aging and HIV-1 infection have led to several investigations into HIV-1-mediated aging of the immune system. In this article, we review various studies that report alterations of leukocyte number and function during aging, and compare those alterations with those observed during progressive HIV-1 disease. We pay particular attention to changes within lymphoid tissue microenvironments and how histoarchitectural changes seen in these two diseases affect immunity. As we review various immune compartments including peripheral blood as well as primary and secondary lymphoid organs, common themes arise that help explain the decline of immunity in the elderly and in HIV-1-infected individuals with advanced disease. In both conditions, lymphoid tissues often show signs of histoarchitectural deterioration through fat accumulation and/or fibrosis. These structural changes can be attributed to a loss of communication between leukocytes and the surrounding stromal cells that produce the extracellular matrix components and growth factors necessary for cell migration, cell proliferation, and lymphoid tissue function. Despite the common general impairment of immunity in aging and HIV-1 progression, deterioration of immunity is caused by distinct mechanisms at the cellular and tissue levels in these two diseases.
Leukocyte cellularity and function are drastically altered throughout the normal aging process. The developing immune system found in the very young and the declining immune system in the elderly lack focused functionality, and thus leave these populations vulnerable to infections and cancers not typically found in individuals between extremes of age. In addition to aging, immune dysfunction can be caused by radiation, chemotherapy, or through infectious agents such as Human Immunodeficiency Virus type 1 (HIV-1). Similarities of immune impairment caused by HIV-1 and aging were immediately evident at the beginning of the epidemic when opportunistic infections normally found in the elderly were being reported in younger adults.
Does the degeneration of effective immunity seen in aging and advanced HIV-1 disease have the same root cause? There is intense interest in this question, especially with an increasing population of aging HIV-1-positive individuals. Answering this question is difficult since we still do not have a clear mechanistic understanding of the natural aging process. However, anatomical and physiological signs and symptoms of aging are numerous, including bone and muscle atrophy, cardiovascular disease, and susceptibility to infection. At the cellular and tissue levels, aging has been attributed to intracellular changes, including metabolic alterations, DNA damage, and telomere attrition. Extracellular characteristics of tissue including changes in extracellular matrix (ECM) composition and organization, tissue hardening, and fibrosis are also often seen during aging. In this article, we compare cellular properties within the peripheral blood of aging or HIV-1-infected individuals, and then shift our focus to changes in the extracellular microenvironments of lymphoid compartments that occur during aging and HIV-1 disease. Are there common cellular and histoarchitectural changes that occur in the immune compartments during aging and HIV-1 infection that influence the diminished immunity seen in both?
Although both aging and HIV-1 progression cause dysregulated innate and adaptive immune responses, these two diseases have distinct mechanisms for degenerative immunity. Myeloid and lymphoid cell population numbers and function are altered differently in aging and progressive HIV-1 disease. These differences at the cellular and tissue levels that both lead to impaired immunity still share a key similarity. Depletion of one or more subsets of leukocytes causes a disruption of a delicate relationship between leukocytes and surrounding stromal cells within lymphoid compartments. These populations provide growth factors and vital survival cues to each other that help build the tissue architecture required to build an effective immune response. Restoring the interdependent relationship between leukocytes and stromal cells may reverse the impairment of immunity in aging and in HIV-1 disease.
Aging and HIV-1-Mediated Alterations of the Peripheral Blood
A precipitous decline in absolute numbers of peripheral blood lymphocytes (PBMC), decreased myeloid cell function, chronic immune activation, and proinflammatory cytokine production are common manifestations in both aging and HIV-1 disease. In aging, there is a dramatic decline in peripheral blood B cells, a gradual decline in naïve lymphocytes, and an increasingly dysfunctional innate immune system. In HIV-1 disease, CD4+ T cell depletion leads to abnormal activation and dysfunction of innate and adaptive immunity. Differences between peripheral blood myeloid and lymphoid populations in aging and HIV-1 infection are discussed below.
Aging has a profound effect on both innate and adaptive immune functions; however, the decreases in absolute number of peripheral blood subsets are predominantly found in lymphocyte populations. As an individual ages, the absolute numbers of circulating neutrophils and other myeloid cells do not change significantly 1 ; however, they tend to produce increasing levels of proinflammatory cytokines and are impaired in effector functions such as phagocytosis. This decline in innate immunity leads to increased opportunistic infection in the elderly. 1 –3 Hearps et al. investigated the peripheral blood of 146 healthy adults, and observed an age-related alteration in monocyte phenotype and function. Aged monocytes were less capable of phagocytosis, produced more proinflammatory cytokines such as TNF-α, and also had shortened telomeres. 4 In contrast to myeloid subsets, lymphocyte numbers declined greatly during the aging process. Valiathan et al. measured the percentage and absolute number of peripheral blood lymphocyte subsets of 191 individuals separated into five different groups based on age, ranging from infants within their first year of life to the elderly, defined as being 70–92 years of age in their study. 5 They reported that the age-related decrease in peripheral blood lymphocytes was primarily due to drastic decreases in CD19+ B cell numbers and to a lesser extent in CD4+ and CD8+ T cells. The absolute count of B cells dropped from its peak of 1,375 ± 141 cells/mm3 in childhood to 198 ± 34 cells/mm3 in the elderly. Decreased lymphocyte number and function corresponded to reduced responsiveness to vaccination in the elderly.
Although aging and HIV-1 disease both lead to a decline in immune function, there are differences in both peripheral blood myeloid and lymphoid cell alterations. In advanced HIV-1 disease, various hematological abnormalities occur, which can affect myeloid subsets. Parinitha and Kulkarni 6 analyzed blood samples from 250 HIV-1-infected individuals and observed anemia in 91.4%, leukopenia in 26.8%, lymphopenia in 80%, and thrombocytopenia in 21.7% of individuals with CD4+ T cell counts <200 cells/mm3. Granulocytopenia and decline in myeloid dendritic cells (mDC, CD11c+) and plasmacytoid dendritic cells (pDC, CD11c−) occurred when CD4+ T cell counts declined <200 cells/mm3 and HIV-1 viral loads increased. 7 The average peripheral blood mDC counts dropped from 6,978 cells/mL to 2,298 cells/mL and pDC counts decreased from 9,299 cells/mL to 1,640 cells/mL between uninfected controls and individuals with viral loads >105 HIV-1 RNA copies/mL. 7 After antiretroviral therapy (ART), mDC numbers returned to baseline; however, the pDC number did not return to baseline and their function remained suppressed. 8 Unlike in aging, there is a profound decline of peripheral blood myeloid cells in advanced HIV-1 disease.
Similar to aging, lymphocyte subsets are significantly altered in HIV-1 disease. However, there are differences in which lymphocyte subsets are most affected. Although CD4+ T cell loss is most profound in HIV-1 infection, other lymphocyte subsets are functionally altered. Kalayjian et al. investigated common T cell correlates of HIV-1 infection and aging. 9 They measured proliferative capacity, exhaustion markers, and functionality of CD4+ and CD8+ T cells in two adult age groups (≤30 years old and ≥45 years old). Naïve CD8+ T cells lost CD28 expression with increasing age. Although they observed decreased T cell proliferative capacity, increased apoptosis, and decreased delayed-type hypersensitivity responses in the HIV-1-infected groups, they were not able to associate these with age. Gianesin found reduced T-cell receptor excision circles and CD8+CD45RA+CD31+ recent thymic emigrants in the peripheral blood of the HIV-1-infected children compared with age-matched controls. Furthermore, they found that higher percentages of CD8+ T cells in the HIV-1-infected group express markers for senescence (CD28−CD57+), activation (CD38+HLA−DR+), and exhaustion (PD1+) compared with uninfected controls. 10 Cao et al. stratified HIV-1-infected individuals based on disease progression rate, and reported that fast disease progression was associated with increased loss of CD28 on all T cells, a selective decrease in CD31 on CD4+ T cells, and an increase of CD57 on CD8+ T cells. 11 In contrast, Lee et al. found that although there was an increase in CD8+CD28− cells, there was only an increase in CD8+CD57+ cells in aging, not during HIV-1 infection. 12 ART is reported to restore CD45RA+CD31+ T cells to age appropriate levels; however, the CD45RA+CD31− cells remain at decreased levels. 13 It is the CD45RA+CD31− cell population that is thought to maintain the numbers of naïve CD4+ T cells during aging without HIV infection. 14 A decrease in naïve T cells is one common outcome between aging and HIV-1 disease; however, the ablation of the CD4+ T cell subset is more attributable to HIV-1 infection.
Since the beginning of the HIV-1 epidemic, it was known that B cell number and functionality drastically change in HIV-1 infection. A historical perspective on B cell dysfunction in HIV-1 infection in the presence or absence of ART is well summarized by Moir and Fauci. 15 Similar to aging, chronic HIV-1 infection leads to decreased peripheral B cell number and functional impairment of class-switch recombination. 16 HIV-1-positive individuals have reduced antibody titers to previously exposed antigens and have impaired development of robust protective humoral immunity after vaccination compared with HIV-1-negative age-matched controls. 17
Aging and HIV-1 infection both lead to impaired immunity, but there are differences in leukocyte subset depletion. Despite these differences, immune impairment in both conditions may be linked to chronic inflammation and leukocyte telomere attrition. Similar to aging, HIV-1 infection leads to decreasing numbers of naïve lymphocytes and chronic inflammation along with increased production of proinflammatory cytokines such as TNF-α, IFN-γ, CXCL10, soluble CD163, soluble CD14, neopterin, and CD16+ monocytes. 18,19 Telomere shortening is also seen in both aging and HIV-1 infection. 13,20 In HIV-1 disease, the greatest period of PBMC telomere shortening occurs during acute infection. 21 Gianesin et al. assessed the impact of HIV-1 infection on accelerated aging in children, and reported shortened telomere length within PBMC of 71 HIV-1-infected children compared with 65 HIV-exposed uninfected children and 56 HIV-negative children. Their findings suggest that ART may dampen this shortening effect. 10 Telomere length was also measured in adult PBMC, CD4+ T cells, and CD8+ T cells to assess the effect of HIV-1 infection on aging. Bestilny et al. reported at least a fivefold increase in cellular aging due to HIV-1 infection, as measured by telomere decrease in the HIV-1-infected cohort. They indicate that a 37-year-old HIV-1-infected individual has similar telomere length to an uninfected 75-year old. 21 T cells, especially CD8+ T cells, are most profoundly affected by telomere shortening in HIV-1 infection. 20,21 Telomere shortening rate is significant in the elderly, but not nearly as profoundly affected as those with late-stage HIV-1 infection. 21
In summary, HIV-1 infection and normal aging both cause functional declines in adaptive and innate immunity. There is a large decrease in functional B cells in the periphery and a decline in circulating naïve lymphocytes. Myeloid cell function and innate immunity decrease as persistent chronic inflammation occurs. Despite these similarities, there are differences in circulating leukocyte subsets in normal aging and HIV-1 infection. In aging, there is a drastic reduction of peripheral B cells, whereas HIV-1 infection severely reduces the circulating CD4+ T cell numbers. As for myeloid subsets, HIV-1 infection affects the numbers of circulating myeloid cells more than the normal aging process. Normal aging of the immune system and HIV-1 infection both impair immunity, but not by the same mechanism. As only a small number of lymphoid cells are present in the peripheral blood while the majority of lymphoid cells are located in lymphoid tissues such as lymph nodes and mucosal associated lymphoid tissues (MALTs), we now shift our focus from circulating leukocyte composition to age- and HIV-1-mediated alterations of lymphoid tissue cellularity and histoarchitecture.
Aging and HIV-1-Mediated Alterations of the Bone Marrow
The bone marrow is a critical developmental niche for CD34+ hematopoietic stem cells (HSCs) that give rise to adult blood, including the myeloid and lymphoid cells required for innate and adaptive immunity. 1,2,22 –25 In the adult human, hematopoiesis occurs primarily in the red bone marrow of flat and irregular bones, but it is known that the progenies of the HSCs are skewed in both aging and HIV-1 disease. How is the bone marrow affected during the aging process and does HIV-1 infection mimic these age-related changes?
Fetal hematopoiesis begins in the liver and then migrates to the bone marrow, where the majority of adult hematopoiesis occurs. 26 In aging humans, the leukocyte progenitor cell composition within the bone marrow changes over time. Similar to the peripheral blood subsets, bone marrow myeloid lineage cells retain their production levels while common lymphoid and early B cell progenitors decline in number. 22,27 The aged bone marrow niche has been linked to peripheral myeloid lineage skewing and phenotypically different myeloid neoplasias seen in the elderly. 28,29
Bone marrow alterations in HIV-1 disease also mirror the various conditions seen in the peripheral blood but are more pronounced than those seen in normal aging. Several early studies reported a variety of hematological abnormalities in the bone marrow of HIV-1-infected individuals. A report of bone marrow abnormalities in 160 HIV-1-infected individuals showed that of the 107 individuals who were diagnosed with AIDS, 93.12% were anemic. The most significant difference between the AIDS and non-AIDS groups was evidence of myeloid dysplasia, primarily in granulocytes. 30 Tripathi et al. reported that erythroid dysplasia was found in the bone marrow of >50% of HIV-infected individuals, and one-third of them had abnormal granulocytic and megakaryocytic development and elevated levels of plasma cells. 31,32 Another analysis of 102 individual bone marrow aspirates showed indications of hypercellularity, dysplasia, plasmacytosis, and lymphoid infiltrates in AIDS individuals, which correlated with anemia and granulocytopenia. 33 Sun et al. examined bone marrow biopsies of 20 individuals diagnosed with AIDS and 39 individuals with asymptomatic HIV-1 infection. Anemia and leukopenia were associated more with an AIDS diagnosis, while thrombocytopenia was more common in the group with less advanced disease. 34 Delacretaz et al. examined bone marrow biopsies of 18 HIV-1-infected individuals between December 1981 and December 1986. 35 Although various bone abnormalities were found, 17 of 18 individuals had myelodysplasia, including dysmegakaryocytopoiesis (n = 16), dyserythropoiesis (n = 15), and dysgranulopoiesis (n = 6). Nearly half of the biopsies showed signs of lymphocytic clusters that appeared to be B cells while over half of the specimens had increased macrophages, a known target for HIV-1 infection.
Are the various bone marrow alterations seen in advanced HIV-1 disease due to direct infection? HIV-1 infection inhibited in vitro hematopoiesis from bone marrow histocultures 36 ; however, there is ongoing debate whether CD34+ HSC can be infected by HIV-1. 37 –40 Other cells within the bone marrow, including CD4+ monocyte precursors, may harbor HIV-1. Sun et al. detected HIV-1 nucleic acids through a cDNA probe in various cells, including mononucleated lymphocytes and histiocytes. However, endothelial cells, interdigitating reticulum cells, nucleated red blood cells, and immature myeloid cells were also reported to possibly contain HIV-1 nucleic acids. 34 Infection of these monocyte precursors or accessory stromal cells may alter important leukocyte to stromal cell communication and skew the myeloid compartment function during chronic HIV-1 infection. 41,42 Aside from immediate cytopathic effects of HIV-1, bone marrow fibrosis is seen in AIDS and other diseases, which may also contribute to dysfunctional hematopoiesis.
The bone marrow has an optimized microarchitecture of stiff bony trabeculae, mesenchymal cells, adipocytes, vasculature, and growth factors for the homeostatic balance of the HSC population and continuous production of red and white blood cells. 43 –45 The stromal cells produce several ECM proteins, including collagens type III and IV, fibronectin, hyaluronan, laminin, and tenascin, each with their own role in supporting resident cell functions. 22,43,44 The ECM provides a necessary scaffold for attachment, migration, and acts as a depot for survival and growth factors (Fig. 1). Irregular production or organization of the ECM can lead to niche dysfunction and impaired hematopoiesis. Overgrowth of certain types of ECM fibers including collagen type I within a tissue is termed fibrosis and is often a marker for disease. How do age- and HIV-1-infection-related fibrosis affect hematopoiesis in the bone marrow niche?
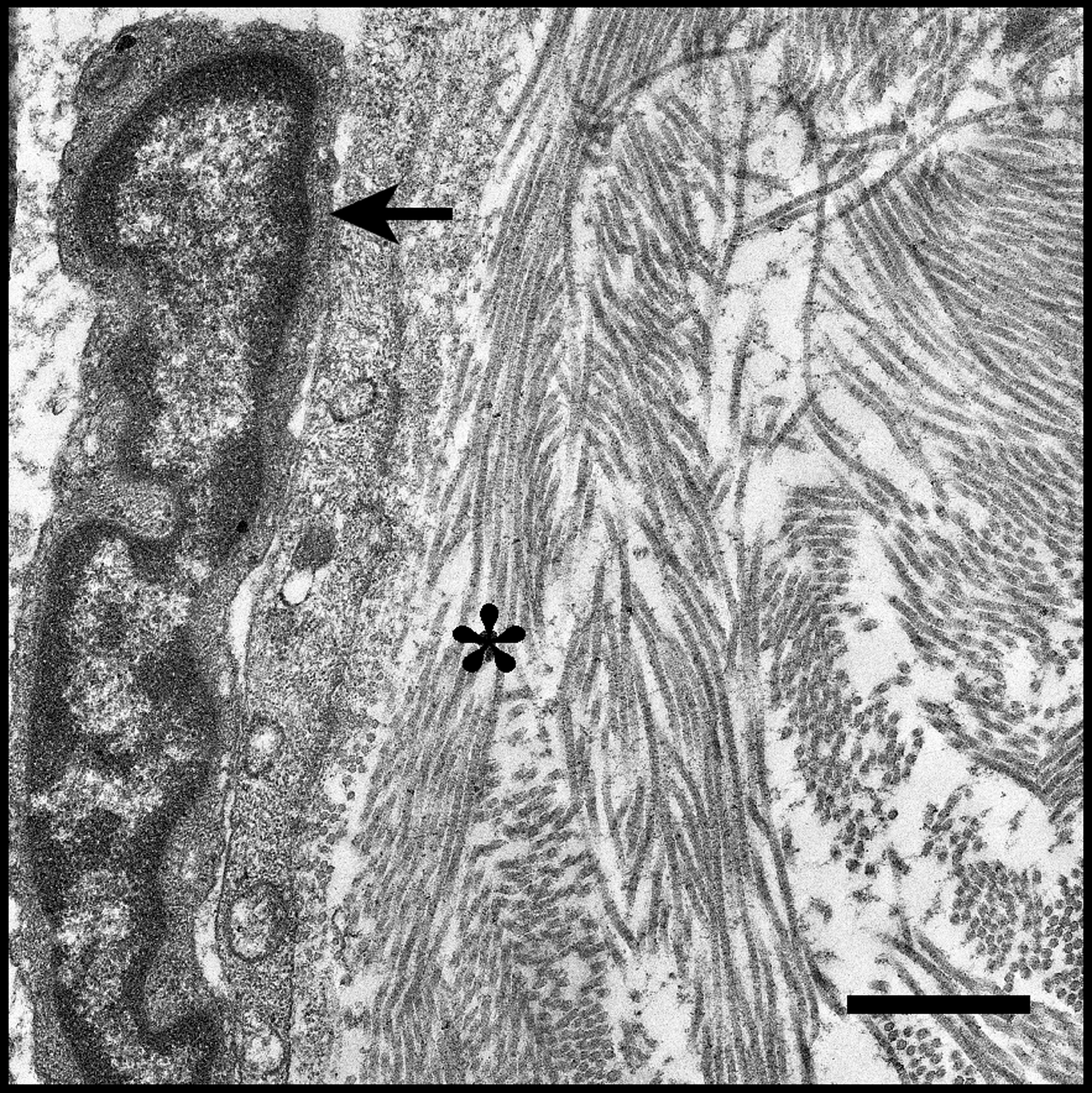
Leukocyte migrating along ECM in human tonsil tissue. The nucleus of the leading edge of a migrating cell (arrow) can be seen on the left of the image. Migrating cells adhere to ECM proteins and neighboring cells. Lymphoid tissue architecture is dependent on ECM proteins such as the collagen seen in this image (asterisk) and the stromal cells that produce ECM. Disruption of communication between leukocytes and stromal cells in lymphoid tissues leads to aberrant deterioration or excessive buildup of ECM proteins, which can hinder tissue function. Scale bar = 1 μm. ECM, extracellular matrix.
Kuter et al. associated two different types of bone marrow fibrosis, reticulin and collagenous, with severity of disease. 46 Reticulin fibrosis is composed of glycosaminoglycans along with individual or loose fibrils of collagen III surrounding a core of collagen I. Reticulin fibrosis appeared to be reversible, and often seen in nonmalignant diseases and to a lesser severity in a majority of healthy controls. 46 On the contrary, collagen fibrosis in the bone marrow was shown to be linked to more severe irreversible disease conditions. Kuter et al. defined collagen fibrosis as an abundance of collagen I within the bone marrow. Collagen I has a thicker fiber diameter than reticulin fibrosis, and increased collagen density may impede cell migration, communication, and tissue function. 46
Severe bone marrow collagen fibrosis is not a typical phenotype of the normal aging or HIV-1-infected bone marrow. 46 However, O'Malley et al. reported moderate to severe reticulin fibrosis within the bone marrow of the majority of the 35 HIV-1-infected individuals in their study. 47 As in previous studies, the reticulin fibrosis was associated with low-affinity nerve growth factor receptor positive adventitial reticular stromal cells (ARCs), 46,47 and ARC quantities inversely correlated with peripheral CD4+ T cell counts. Despite this correlation, the authors did not find a significant correlation between the CD4+ T cells count and the observed degree of bone marrow fibrosis. 47 Delacretaz et al. also found mild bone marrow reticulin fibrosis in 15 of 18 individuals in their study. 35 In seven of the individuals, they found niche deterioration, including hypoplasia, fat atrophy, and gelatinous infiltration. The ECM production of both types of fibrosis is associated with TGF-β1 signaling 46 in bone marrow stromal cells, including ARC, perisinusoidal adventitial cells, periarterial adventitial cells, endosteal cells, and adipocytes. Disruption of stromal cell function and ECM production by alteration of leukocyte numbers and phenotype may be the underlying common cause of histoarchitectural changes found in the aging and HIV-1-infected bone marrow.
In both aging and HIV-1 infection, the bone marrow compartment mirrored the peripheral blood leukocyte alterations; however, aging and HIV-1 infection differed in specific subset alterations. Although bone marrow lymphoid progenitors were the primary subset of cells that declined greatly in normal aging, multiple myeloid and lymphoid subsets are affected in progressive HIV-1 disease. While reticulin fibrosis was seen in both conditions, this was reported to be reversible compared with the collagenous fibrosis seen in bone marrows of leukemias and other hyperproliferative disorders. Aging and HIV-1 infection both skew hematopoiesis in the bone marrow; however, advanced HIV-1 infection causes an increased number of leukocyte abnormalities in the bone marrow compared with the normal aging process.
Aging and HIV-1-Mediated Alterations of the Thymus
The thymus is the primary site of normal T cell development during fetal and neonatal life. A human thymic rudiment with T cell precursors is present as early as 7–8 weeks of gestation, 48 and a fully formed thymus consists of several lobules with cortical and medullary areas, which contain developing thymocytes, epithelial cells, macrophages, dendritic cells, and other hematopoietic cells including B cells. T cell progenitor migration into the thymus, intrathymic T cell development, and emigration of mature naïve T cells to the periphery are all involved in maintenance of cellular immunity. 49
Several murine studies suggest that there is an interdependent relationship between the thymic stroma and the developing thymocytes, each providing vital cytokines. Cortical and medullary thymic epithelial cells (TECs) are of endodermal origin, and differ in their functions and expression of cell surface markers. Recent data show that cortical as well as medullary epithelial cells are heterogeneous. 50 The heterogeneity of the various epithelial subpopulations provides the proper developmental cues for functional and self-tolerant T cells. 50 It is important to note that TECs are essential for T cell development and reciprocally, developing thymocytes are critical for the maintenance of TECs and a functioning thymus. 51
T cell precursors from fetal liver or bone marrow adhere to the thymic vascular epithelium and cross the perivascular epithelial cell layer to enter the thymus. 52,53 Integrins such as VLA-4/α4β1 and VLA-5/α5β1 are expressed on thymocytes, and used to adhere to and migrate along thymic epithelium and fibronectin ECM. 54,55 The developing thymocytes are guided by chemokine gradients produced by the thymic stroma, 56 –58 including TEC-produced CXCL12. 59,60 As the thymocytes migrate throughout the cortex and medulla, they are provided prosurvival cues from cytokines like interleukin-7 (IL-7), which is produced by the thymic stroma. 61 IL-7 administration has been proposed as a potential therapeutic approach to thymus reconstitution after aging and HIV-1 infection. 62
T cell development is characterized by differentiation and proliferation of thymocytes. Loss and acquisition of cell surface markers characterize various stages of T cell differentiation in the thymus. 49 Positive selection and generation of CD4+ and CD8+ single positive cells require engagement of CD3/TCRαβ complex on the CD4+CD8+ double positive thymocytes at low affinity with self-peptide-MHC complexes on the cortical TECs. Thymic medullary epithelial cells and hematopoietic cells presenting self-antigens are essential for negative selection of T cells by inducing the deletion of cells with high affinity to self-antigens, thereby preventing autoimmunity. After positive and negative selection, only ∼5% of thymocytes reach the stage where they can exit the thymus. The most mature thymocyte subset, which is ready to leave the thymus, displays the following phenotype: CD3+/highCD45RA+CD27+CD31+, expresses either CD4 or CD8, and the Sphingosine 1 Phosphate Receptor 1 (S1P-R1), but not CD69. 63,64
A decline in thymus function related to thymic volume is observed in aging. 65 Mackall et al. showed a decline in T cell regeneration related to age after chemotherapy 66 ; however, normal thymus tissue continues to be present and functional late in adult life. 67 With age, thymic structures become less ordered and functional, and thymic output decreases consistently. It is also important to understand other major factors that may affect the function and degradation of the thymus, such as HIV-1 infection.
A powerful method to identify differences between typical age-dependent thymic involution and the manner in which the thymus of a HIV-1-infected individual changes over the course of infection is to compare the thymi of age-matched individuals. 68,69 With a cohort of nontreated HIV-1 seropositive adults (n = 99, median age 39 years), as well as one of HIV-1 seronegative adults (n = 32, median age 32 years), McCune et al. used noncontrast chest CT imaging to show evidence that support that younger individuals were more likely to retain thymic tissue than older counterparts, regardless of infection status. 70 These conclusions were made after the establishment of a thymic index of the cohort, which compared age, serostatus, years passed since seroconversion, and CD4+ T cell counts. The investigators added that in the HIV-1 seropositive subjects, a higher thymic index was associated with higher CD4+ T cell counts.
The thymus is severely affected by CCR5-tropic and CXCR4-tropic HIV isolates. 71 CCR5-tropic HIV-1 isolates impair T cell development although the percentage of thymocytes expressing the HIV-1 coreceptor CCR5 is much less than those expressing CXCR4. 58,72,73 However, despite low overall expression in the thymus, CCR5 is still expressed on the more mature thymocytes resulting in a significant impact of T cell reconstitution by CCR5-tropic HIV-1 isolates. 58,72,73 Infection of the thymus in utero and in early life by HIV-1 isolates pathogenic for T cell precursors was reported to have repercussions not only in situ, but also affect the overall course of disease in children. 73,74 Involvement of the thymus in pediatric AIDS was also suggested by histological studies showing thymic involution during HIV-1 infection of thymocytes in fetuses and children, and by anomalies in peripheral T cell subset distribution in HIV-1-infected children. 75 –77 Together, these findings suggest that HIV-1 has a deleterious effect upon the thymus of infected individuals, generally showing greater thymic involution than uninfected individuals of similar age, though the impact of this involution at these ages is of varying importance.
Involution of the thymus, whether by aging or HIV-1-associated destruction, is a serious structural and functional change that leads to long-lasting effects, contributing to the onset of immune senescence. The lack of a site for extensive T cell proliferation and maturation effectively acts as a bottleneck for the overall efficacy of the adaptive immune system, and has a cascading effect upon secondary lymphoid compartments. Fewer mature T lymphocytes means decreased overall immunity, resulting in an increased probability of serious infection, not to mention other housekeeping tasks, such as lymphatic and circulatory filtration and surveillance. However, it is important to also note that while this process of atrophy is currently understood to be an inevitable byproduct of aging, there exists the potential for the identification of strategies to mitigate, halt, or even reverse the involution of the thymus. It has been well documented in the young that malnutrition leads to rapid involution of the thymus, but the reintroduction of proper nutrition allows for rapid regeneration of the tissue. 78 With the use of ART, HIV-1-infected individuals may recover significant functional thymic mass after its degradation by effects of the virus. Douek et al. measured thymic output in HIV-1-infected individuals and observed a decline in thymic emigrants in aged as well as HIV-1-infected individuals; however, ART could rapidly reverse this decrease in thymic output in HIV-1-infected individuals. 79 As not all HIV-1-infected individuals recover their CD4+ T cell levels while on ART, Rb-Silva et al. developed a mathematical model to predict immune reconstitution in HIV-1-infected individuals started on ART based on thymic function. 80
However, as normal thymus tissue continues to be present and functional late in adult life, 67 the impact of aging on the thymus does not completely reflect the structure and function of the thymus in HIV-1 infection even in the era of ART.
Aging and HIV-1-Mediated Alterations of the Lymph Nodes
The general structure of the lymph node is similar to the thymus, with an inner medullary region and an outer cortical region containing hyperproliferative cells surrounded by a fibrous capsule. Between the cortical region and the capsule is a sinus that transports draining lymph, antigens, and leukocytes from the afferent lymphatics and sites of inflammation. 81,82 Lymphocytes, primarily T and B cells, along with nutrients from the blood enter the secondary lymphoid tissues through specialized high endothelial venules (HEVs). If an infection is present in the body, an incoming flow of lymphocytes and nutrients from the blood will intersect with an incoming flow of antigens and antigen presenting cells (APCs) from the afferent lymphatics. This intersection provides all the necessary components to build an adaptive immune response.
Similar to other immune compartments, the architecture of the lymph node is built through intercellular communication between lymphocytes and stromal cells. Stromal cells within the lymph node including fibroblastic reticular cells (FRCs) are critical in creating the matrix and reticulum where leukocytes interact. FRCs also produce IL-7, a prosurvival signal for naïve T cells, 83 and require lymphotoxin-β (LTβ) signaling from the infiltrating T cells, 84 emphasizing the delicate relationship between lymphocytes and stromal cells within lymphoid compartments. The cortical reticulum where B cell follicles reside is a more open and loose mesh network compared with the medullary T cell zone, which is more compact and has a higher density of fibrous networks 85 composed of ECM proteins such as collagen I and III, fibronectin, and tenascin 86 (Fig. 2).
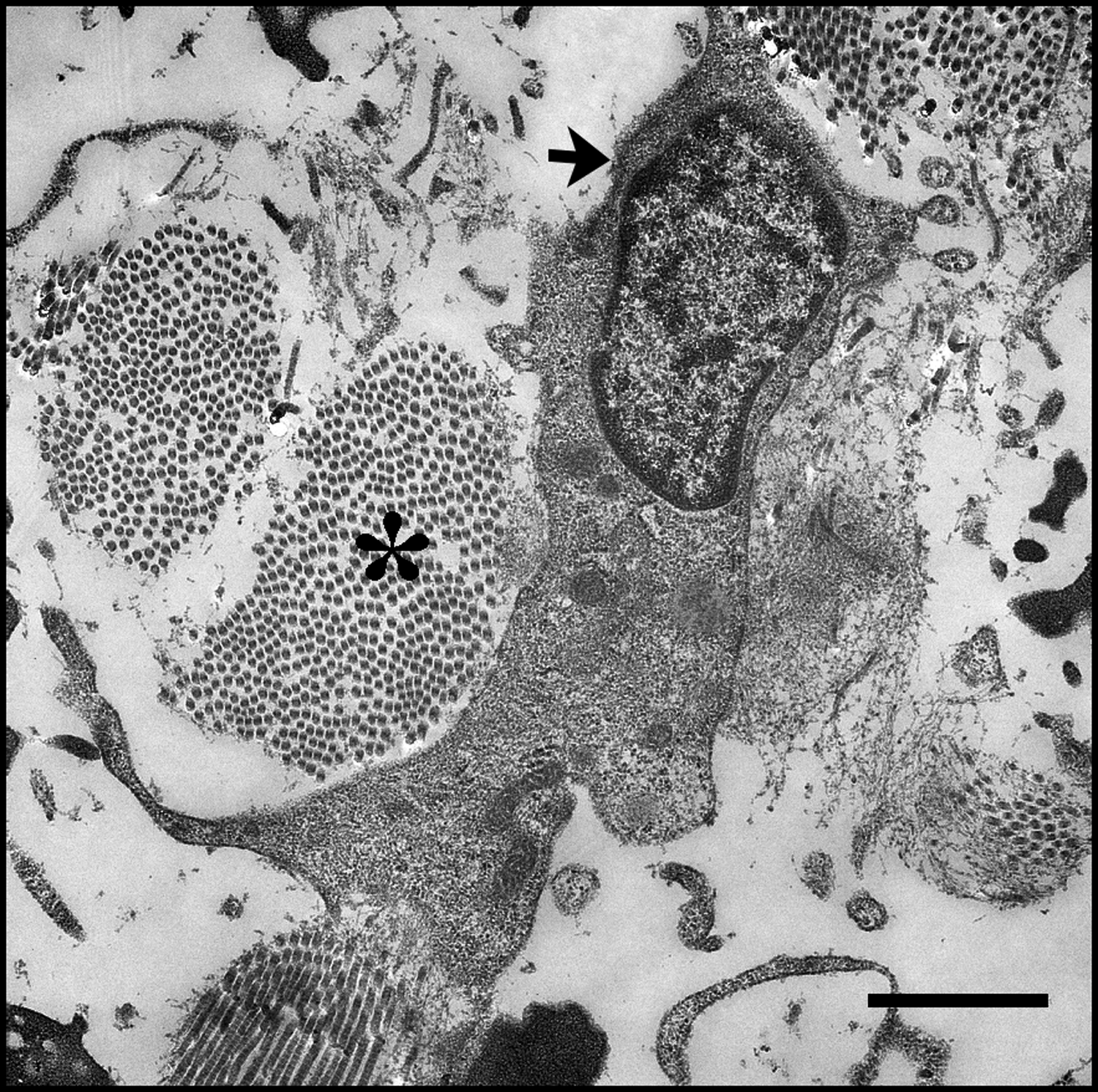
Polarized cell adherent to adjacent collagen bundles in lymphoid tissue. This migrating lymphocyte (arrow) makes several contacts to the ECM proteins like the collagen bundles seen here (asterisk). Proper density and placement of these architectural components within lymphoid tissues are critical for leukocyte migration and function. Matrix degeneration, fibrosis, and lipomatosis of lymphoid tissues have previously been associated with various diseases, including aging and progressive HIV-1 disease. Scale bar = 2 μm.
The age-related decline in ECM-producing stromal cells contributes to changes in lymph node morphology and impairment of adaptive immunity in the elderly. Thompson et al. provide a concise summary of how these stromal cells deteriorate with age. 83 Lymphatic endothelial cells become more permeable and result in decreased antigen flow from the tissues into the lymph node. Furthermore, aged HEVs become less permeable and hinder lymphocyte entry into the lymph nodes. Decreased flow of antigens, APCs, and lymphocytes into the lymph node dampens adaptive immune responses and levels of lymphocyte-derived stromal cell growth factors, causing further degeneration of lymph node histoarchitecture. The follicular dendritic cells (FDCs) that present antigen and immune complexes within the cortical B cell follicles and the FRCs that produce the medullary reticulum and provide a network for the interaction of T cells and dendritic cells also decrease with age.
Several studies have looked at lymph node architecture and deterioration throughout the aging process. In 1947, Denz described age-associated morphological differences in >300 lymph nodes from 150 autopsies. 85 Hadamitzky et al. studied age-related histoarchitectural changes in 41 superficial inguinal lymph nodes from individuals ranging in age from 17 to 98 years. 87 Much of this early work was recapitulated in later studies, including Tsakraklides et al. in their 2,250 axillary lymph node study and Luscieti et al. who also found age-related and location-specific structural changes in human lymph nodes. 88 The general consensus is that lymph node histoarchitecture varies based on underlying disease, lymph node location in the body, and exposure to antigens.
The most pronounced age-related morphological change found by Luscieti et al. and others was the decrease in germinal center size and number. 88,89 Cortical germinal centers do not develop until 2–6 months after birth, 85 concomitant with foreign antigen exposure, 88,89 and maximum lymph node size is reached in children by 7–10 years of age. 85 Development of germinal centers is more prominent in spherical cervical (CLN), mesenteric (MLN), and lumbar lymph nodes that are found deep within the body and peripheral to the mucosa as opposed to flattened superficial axillary, popliteal, or cubital lymph nodes. 85,88 The discrepancy in germinal center number and size is thought to be associated with amount of antigen exposure between mucosal and superficial locations. After puberty, the superficial lymph nodes begin to show signs of degeneration, including increased fibrosis, fatty tissue accumulation (lipomatosis), and decreased cellularity (Fig. 3). Contrary to superficial lymph nodes, deep nodes such as CLN or MLN in individuals ≥80 years contained germinal centers, although their activity and number appear to decrease with age. 85
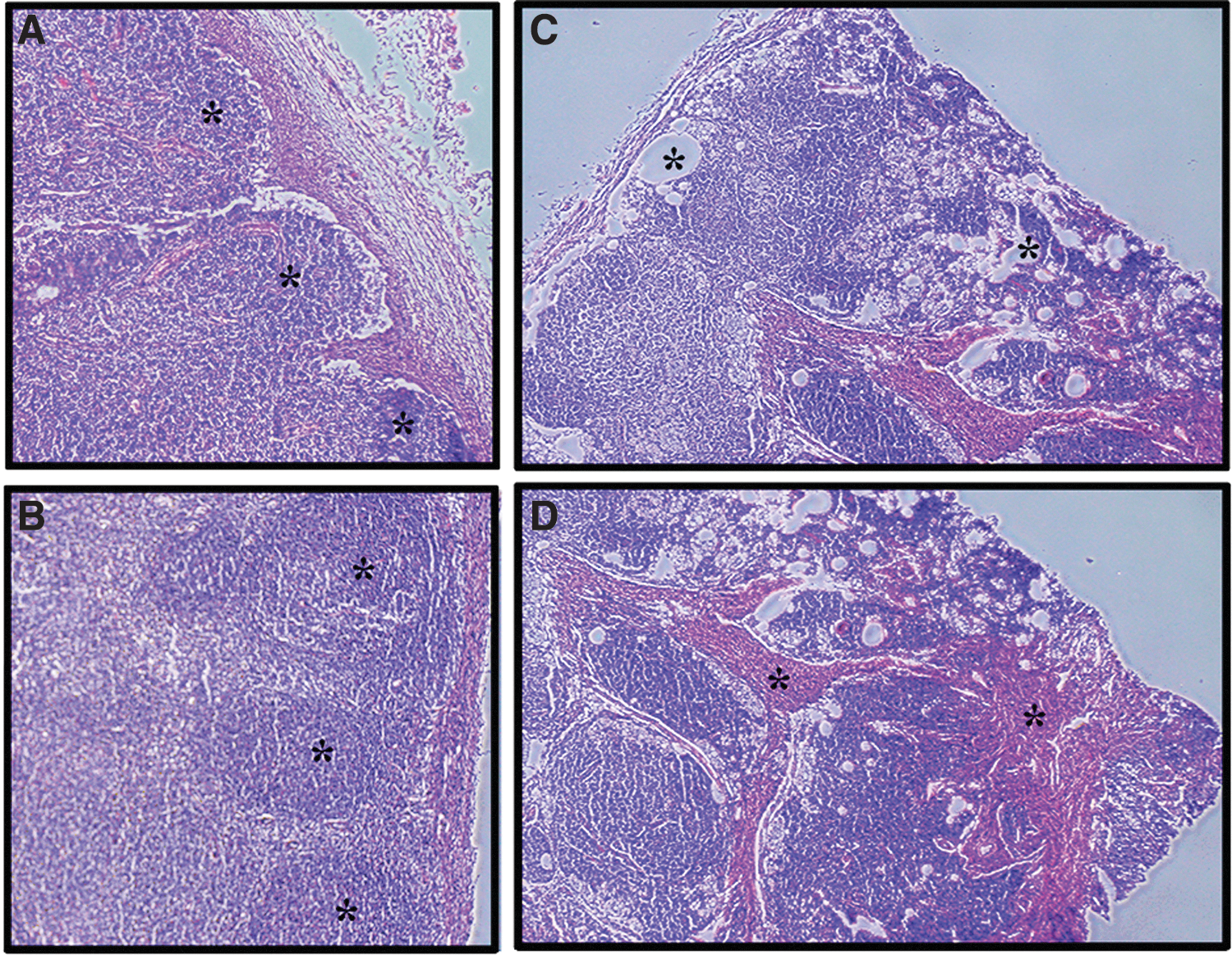
Age and active HIV-1 replication affect lymph node histoarchitecture. Hematoxylin and eosin staining of sectioned lymph nodes from HIV-1-infected individuals show how antigen exposure and age may affect tissue structure.
Several of the studies indicate that lipomatosis and fibrosis occur independently. Fat accumulation was rarely seen in lymph nodes from the youngest groups but was consistently found in those from the older groups. 87 Lipomatosis was not often seen in mesenteric lymph nodes with continuous antigen exposure but could readily be seen in superficial axillary and inguinal nodes. 85 Although fibrosis was more severe and prominent in the older age groups, mild fibrosis was still seen in the youngest cohort, indicating that fibrosis is not solely dependent on age. 87 Denz reported that beginning at age 30, the medullary reticulum begins to decrease in cellularity and increase in collagenous material with limited if any lipomatosis. 85,89,90 Fibrosis only occurred after medullary lymphocyte decline and subsequent compaction of reticulum fibers, yet the fibrosis did not impair medullary cell proliferation during active inflammation.
Similar to aging, HIV-1 infection causes substantial deterioration of lymph node architecture, including germinal center alterations and fibrosis. Several studies have investigated HIV-1-mediated effects on lymph node structure and function. Paiva et al. evaluated cervical lymph node biopsies of 31 adult HIV-1-infected individuals with or without AIDS before ART. 91 Schacker et al. compared inguinal lymph node histology of 33 individuals, 24 of which were HIV-1 positive and ART naïve, while 9 were from HIV-1-negative individuals. 92 In another study, Schacker et al. analyzed inguinal lymph node fibrosis of 11 HIV-1-positive individuals before and after ART administration. 93
A general theme emerges from these studies. Chronic antigen exposure leads to prominent follicular development and lymphadenopathy early in infection. Elevated concentrations of HIV-1 antigens can be found in the lymph nodes at concentrations >300 pg/mL and are thought to drive the follicular hyperplasia present during chronic infection. 91,94,95 As the disease progresses, follicular lysis and involution occur in the germinal centers. Atrophy of cortical germinal centers may be due to disruption of intercellular communication between FDC, CD4+ T follicular helper cells (TFH), and B cells. A similar reduction in the FRC network, IL-7 production, and naïve T cell infiltration of the medulla is also seen during HIV-1 infection. 81,84
Decreased stromal cell survival and function may also be due to decreased lymphocyte entry into the lymph node. 91,92 In biopsies from AIDS patients, increased fibrosis was seen in the medulla and the basement membrane surrounding HEVs 92 (Fig. 4). Increased collagen-I and fibronectin fibrosis is also seen in the lymph nodes of nonhuman primate animal models infected with simian immunodeficiency virus (SIV). 81,96 Like the fibrosis seen in other tissues, lymph node fibrosis seen in HIV-1 and SIV infection may be induced by TGF-β1 signaling initiated by surrounding Tregs, 81,84,96 which can stimulate myofibroblastic production of ECM components. 97

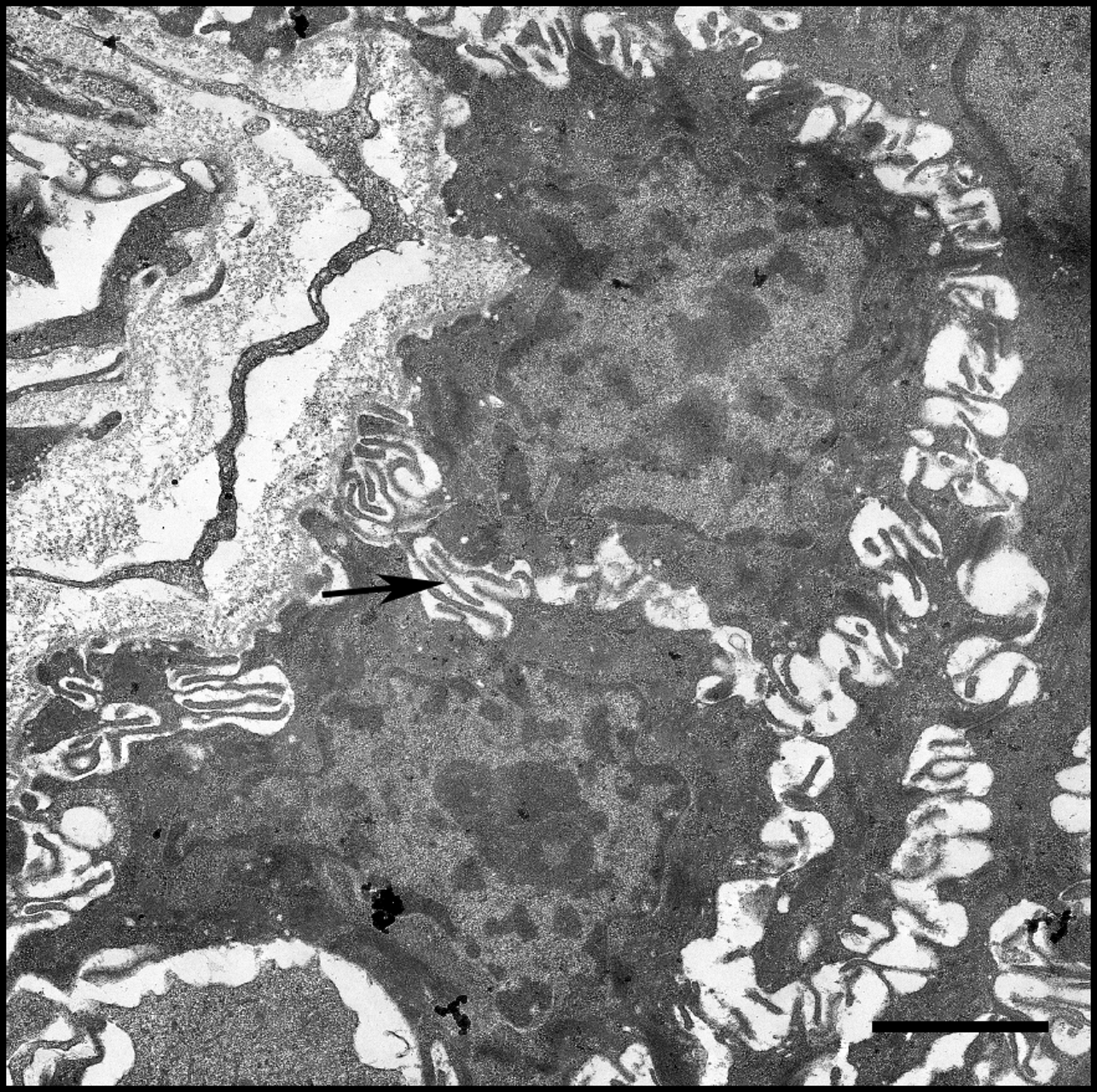
The edge of this blood vessel found within human lymphoid tissue shows several erythrocytes (asterisk) in the lumen of the blood vessel. In the bottom of the micrograph, ECM can be seen below the endothelial cell layer (arrow). Increased fibrosis within the basement membrane of high endothelial venules is thought to decrease lymphocyte homing to lymphoid tissues in both progressive HIV-1 disease and aging. Scale bar = 2 μm.
Aging and HIV-1 disease, through different mechanisms, both lead to deterioration of lymph node structure and function and subsequent impairment of adaptive immunity. 81 After sufficient tissue deterioration and loss of lymphocytes in both aging and HIV-1, lymph nodes become effectively nonfunctional, aside from passing lymph to other nodes. 87
Aging and HIV-1-Mediated Alterations of MALTs
Mucosal associated lymphoid tissues (MALTs) are critical in developing appropriate adaptive immune responses against mucosal pathogens. Along the mucosa, antigen can be transported from the mucosal lumen through M cells to specialized lymphoid inductive sites that generally contain follicles, interfollicular areas, subepithelial dome regions, and overlying epithelium. 98 These regions have distinct T and B cell zones, similar to other secondary lymphoid organs. Although T cells receive inductive signals at these locations, they are thought to migrate to the mesenteric lymph nodes for further differentiation. The differentiated T cells that reside within the inductive sites are effector cells called lamina propria lymphocytes (LPLs). 99,100 Age-related alterations of these dynamically organized mucosal sites of immune effector induction are highly studied in detail in animal models, but the corresponding data in humans are comparatively scarce. 101
MALT effector sites are distributed throughout the mucosa and are the sites of IgA secretion into the mucosal lumen. 98 These tissues can be found throughout the mucosal areas, and include tonsils and adenoids (Waldeyer's ring), Bronchial ALT (BALT), Larynx ALT (LALT), Nose ALT (NALT), and Gut ALT (GALT). GALT includes Peyer's patches, lamina propria of the small intestines, and follicular aggregates of the stomach and large intestines. The majority of these mucosal sites are not typically developed in the fetus, and only become populated by lymphocytes after exposure to antigen and microbes after birth. 98,101,102 Debertin et al. reported that 40% of children studied had developed BALT and NALT, while 80% developed LALT all within the first year of life. 101,102 In adulthood, healthy humans typically lose BALT while 56% of healthy adults still maintain LALT, while NALT presence is still unknown. 101 Beharka et al. investigated how age affects MALT by phenotyping intraepithelial lymphocytes, LPL, and PBMC in addition to measuring immunoglobulin A (IgA) levels in the saliva and serum of young (20–40 years old) versus older (≥65 years) individuals (n = 17). They did not find a significant change in salivary IgA, but reported an increase in serum IgA with age. 103
Although there are limited data on age-related changes of MALT in humans, several reports on Peyer's patches and GALT in aging rodents mirror what happens in human lymph nodes. With age, there is a decrease in naïve CD4+ T cells and FDCs within GALT, which leads to decreased germinal center development and altered gut IgA levels. 104,105 The decrease in lymphocytes leads to a decrease in LTβ signaling, stromal cell survival, and organization of MALT, once again emphasizing the interdependent relationship between lymphocytes and stromal cells in developing these secondary lymphoid tissues. 106
Chronic antigen stimulation and proinflammatory cytokines found in the mucosa provide an ideal niche for HIV-1 replication. Deterioration of the MALT in progressive HIV-1 disease has been associated with chronic inflammation. Several groups have reported the rapid depletion of CD4+ T cells in GALT during acute HIV-1 infection. 96,100,107 Estes et al. examined peripheral blood, inguinal lymph nodes, and terminal ileum biopsies from 35 HIV-1-infected and 11 HIV-1-uninfected individuals to see if morphological changes within these tissues correlate with peripheral CD4+ T cell counts and disease status before and after 6 months of ART. 107 They reported that fibrosis within the GALT prevents optimal reconstitution of naïve and central memory CD4+ T cells in the Peyer's patches, even after treatment. 107 SIV-infected rhesus macaques also have significant loss of Th17 CD4+ T cells in the GALT, 108,109 which has been linked to increased microbial translocation and chronic inflammation in SIV and HIV-1 infections. 110
Although additional human studies are needed to determine the effects of aging and HIV-1 infection on MALT structure and function, both conditions may impair this immune compartment in similar ways to peripheral lymph nodes by disrupting communication between lymphocytes and surrounding stromal cells (Fig. 5).
Cell communication in lymphoid tissues is critical in maintaining tissue structure and function. In this micrograph of human tonsils, several cells can be seen projecting membrane protrusions to communicate with each other (arrow). Signaling between leukocytes and stromal cells is critical for both lymphoid tissue architecture and leukocyte function and survival. Depletion of leukocyte subsets or stromal cells in aging and HIV-1 infection disrupts this interdependent relationship, and alters the structure and function of lymphoid tissues. Scale bar = 2 μm.
Conclusions
How do tissue microenvironments change in immune compartments in an aging individual and how can this be compared with alterations of immune microenvironments seen during HIV-1 disease? Although both conditions lead to impairment of innate and adaptive immunity in the face of chronic inflammation, there are distinct differences between the two. In the peripheral blood and bone marrow of aging individuals, there is often a myeloid skewing due to loss of naïve lymphocytes and drastic reduction in peripheral B cells. On the contrary, HIV-1 infection depletes CD4+ T cells in addition to causing a variety of hematological abnormalities in the peripheral blood and lymphoid tissues that impact both the myeloid and lymphoid lineages.
Gradual deterioration of other lymphoid structures including the thymus, lymph nodes, and MALT is seen in aging. Destruction of these cellular microenvironments is even more prominent in advanced HIV-1 disease. Increased fibrosis, lipomatosis, and subsequent disorganization of secondary lymphoid tissues ensue. 111 –114 Changes in lymphoid tissue structure during aging and HIV-1 infection may be caused by a disruption of a delicate relationship between lymphocytes and the surrounding stromal cells. The interdependent relationship between lymphocytes and the surrounding stromal cells, including FRCs and FDCs in the lymph node, adventitial reticular cells in the bone marrow, and epithelial cells within the thymus deteriorates with age and is exacerbated in HIV-1 infection.
Footnotes
Acknowledgments
The authors thank Dr. Mauricio González-Navarro, Dr. Maria Fernanda Torres-Ruiz, and Dr. Yara Andrea Luna-Villalobos in obtaining the cervical and inguinal lymph nodes as well as Dr. Thomas Troost for supplying tonsillar tissue samples for these studies. They also gratefully acknowledge Dr. Beth Jamieson for her valuable comments and thoughtful review. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health (NIH).
Author Disclosure Statement
The authors have no disclosures or conflicts of interest to declare.
Funding Information
This publication resulted in part from research supported by National Institute of Allergy and Infectious Diseases (NIAID) award AI126617, cofunded by National Institute on Drug Abuse (NIDA), National Institute of Mental Health (NIMH), and National Institute of Neurological Disorders and Stroke (NINDS).