
Abstract
HIV-1-induced cytopathicity of thymocytes is a major cause of reduced peripheral T cells and rapid disease progression observed in HIV-1-infected infants. Understanding the virulence factors responsible for thymocyte depletion has paramount importance in addressing the pathogenesis of disease progression in children. In this study, thymocyte depletion was analyzed following infection with two primary CXCR4-tropic HIV-1 pediatric isolates (PI), PI-2 and PI-2.1, which were serially derived from an in utero-infected infant. Although highly similar to each other, PI-2 showed markedly decreased thymocyte depletion in vitro compared with PI-2.1. Further analysis showed a novel deletion in the Nef protein (NefΔK7S) of PI-2, which was absent in PI-2.1. This deletion inhibited Nef-mediated major histocompatibility complex class I (MHC-I) downregulation in infected thymocytes in vitro and in vivo; in contrast, the mutated Nef continued to downregulate CD4 surface expression in vitro. These results suggest that HIV-1 Nef contributes to thymic damage in infants through selective functions.
Introduction
The thymus is particularly active during early life as the immune system develops in infants. Thymocyte infection with HIV-1 leads to severe thymic involution and dysfunction, resulting in declined thymopoiesis and subsequent depletion of peripheral T lymphocytes. 1 Although direct killing of thymocytes has been suggested as a major mechanism responsible for thymic dysfunction, 2,3 HIV-1 also causes nonspecific activation and proliferation of intrathymic progenitor cells through secretion of proinflammatory cytokines in the thymic microenvironment that result in elevated viral infection, exhaustion of precursor cells, and destruction of thymic architecture. 4,5 These conditions within the thymus have been observed in children infected with HIV-1 and are suggested to be a leading cause of rapid disease progression in perinatally infected children. 5,6 Studies to understand the underlying pathological and molecular aspects of thymus dysfunction in HIV-1 infection suggest that the Nef protein of HIV-1 plays a key role in impairing thymocyte development in addition to interfering with peripheral T cell function. 7 –10
Nef is a phosphorylated and myristoylated 27 kDa protein that is unique to HIV/SIV and is relatively conserved with a 75%–85% similarity within clade B of HIV-1. 11 Nef is a multifunctional protein that is suggested to play multiple key roles in HIV-1 and SIV pathogenesis. 8,12 Deletion of Nef markedly attenuates SIV pathogenicity in the rhesus macaque model of AIDS, 8,13 and dysfunctional Nef has been associated with delayed disease progression in humans. 14 –16 Nef has also been shown to impair T cell development 10,17 and enhance viral replication and infectivity. 18,19 Its expression begins early in the viral life cycle and localizes to the plasma membrane, cytoplasm, and within the budding virions. Nef promotes viral infectivity, immune evasion, and replication capacity by altering the activation state of infected cells. It does so by modulating cell signal transduction pathways and altering cell surface expression of receptors. Nef regulates various cell signaling molecules, such as Hck, Lck, Fyn, Lyn, Vav, Src, PAK1/2 (NAK), MAPK (Erk1), c-Raf1 kinase, PI3K, and Ask1. 20,21 Nef downregulates cell surface receptors such as CD4, 22 –24 major histocompatibility complex class I (MHC-I), 23 –25 MHC-II, and MHCII-associated invariant chain (Ii, CD74), 26 CD28, 27 and host restriction factor SERINC5. 28
Nef-mediated downregulation of surface expression of MHC-I and CD4 has been extensively studied in vitro. The downregulation of MHC-I A and B molecules protects infected cells from CTL-mediated killing. 29 –31 Previously, we described two closely related CXCR4-tropic HIV-1 primary isolates [pediatric isolates (PI)-2.1 and PI-2] serially derived from an in utero-infected infant, and their dissimilar abilities to cause thymocyte depletion. 32 PI-2.1 showed aggressive thymocyte depletion in vitro, while isolate PI-2 showed diminished cytopathicity. Given the central role of Nef in HIV-1 disease progression, we performed a detailed analysis of Nef sequences of these two pediatric isolates and their corresponding abilities to downregulate MHC-I and CD4.
Materials and Methods
HIV-1 pediatric isolates and virus stocks
HIV-1 primary isolates and plasma samples for virus isolation were obtained as close as possible to birth from 10 of 34 children born to 204 HIV-1-seropositive mothers monitored prospectively in the Los Angeles Pediatric AIDS Consortium between June 1988 and May 1995. 32 –34 The isolates were characterized as syncytium inducing (SI) or nonsyncytium inducing (NSI) by using the MT-2 assay. 35 The characteristics of some of the children providing the viral isolates were previously described: PI-2 is an SI HIV-1 isolate obtained 1 week after birth from an infant infected in utero who developed AIDS and died within 6 months. 34 PI-2.1 was isolated from plasma 1 month after birth from the same infant. PI-2 and PI-2.1 isolation and propagation were done by directly adding serum from an infected infant to a pool of purified activated allogeneic CD4+ T cells, prepared as described previously. 36,37 Briefly, allogeneic CD4+ T cells from three normal donors were individually purified by capture in CD4 monoclonal antibody-coated tissue culture flasks (Applied Immune Sciences, Santa Clara, CA) and activated by stimulation with 200 ng/mL anti-human CD3 antibody (OKT3; Ortho Biotech, Inc., Raritan, NJ) plus 5,000 U/mL recombinant human interleukin-2 (IL-2) for 5 days. Cells from three donors were combined, cryopreserved in liquid nitrogen, and then thawed and cultured in medium with IL-2 for 2–3 days before infection. The makeup by phenotype of the CD4+ T cell pool at the time of infection by the pediatric isolates was as follows: 95% CD4+ CD8−, 5% CD4+ CD8+, <1% CD8+ CD4−, 60% CD27+, and 50% HLA-DR+. In addition, 94% of the cells from the stimulated CD4+ T cell pool expressed CXCR4 and 30%–40% expressed CCR5 (data not shown). 32 The same CD4+ T cell pool was used for isolating PI-2 and PI-2.1 virus from plasma (passage 1) and for preparing virus stocks (passage 2) and for preparing the control viruses including CCR5-tropic virus HIV-1 NF-NSX (NF-NSX) and the CXCR4-tropic virus HIV-1 NL4-3 (NL4-3). 38,39 The infectivity of the pediatric isolates and of the molecular-clone-derived laboratory viruses was determined by limiting dilution assays in PBMC. The CCR5-tropic pediatric isolate PI-8 was prepared similarly to PI-2 and PI-2.1 and used as control for the Western blot studies. 32
Cloning and sequencing of the nef gene in pediatric HIV-1 isolates
RNA was isolated from plasma using the TRIzol reagent (Invitrogen, Carlsbad, CA) and was reverse transcribed into complementary DNA (cDNA) using the High-Capacity cDNA Reverse Transcription Kits (Applied Biosystems, Foster City, CA). Reverse transcription was primed with a random primer (Applied Biosystems) and following nested polymerase chain reaction (PCR), the cDNA of full-length nef was amplified. First and second rounds of PCR were carried out using Nef 8687F (5-GTAGCTGAAGGGACAGATAGGGTTAT-3) and Nef 9589R (5-TAGTTAGCCAGAGAGCTCCCA-3) as the outer primer set and Nef 8787 XbaIF (5-GCTCTAGAATGGGTGGCAAGTGCTCAA-3) and Nef 9495R (5-TTATATGCAGCATCTGAGGGC-3) as the inner primer set, respectively. Numbering of primers is based on the NL4-3 sequence. High Fidelity Advantage® HD Polymerase Mix (Clontech, Mountain View, CA) was used for PCR, according to the manufacturer's instructions. After cloning of PCR products into the Topo TA cloning vector (Invitrogen), plasmid DNAs were isolated from at least six colonies for sequencing with the ABI 3130 Genetic Analyzer (Applied Biosystems).
Production of single-round infectious, pseudotyped HIV-1 reporter virus
The reporter HIV-1 construct AA1305#18 plasmid containing an NL4-3 backbone with reporter antigens heat shock antigen (HSA) or mouse CD24 (mCD24) and deletions in Vpu, Env, and Vpr was used to produce VSV-G pseudotyped virus as described previously. 40 Insertion of isolated nef genes was done using PCR products that were gel purified with Quick Spin Column (Qiagen, Valencia, CA) and digested with XbaI and BspEI restriction enzymes (New England Biolabs, Ipswich, MA). Single-round infectious pseudovirus was produced by cotransfecting 293T cells with the whole-genome plasmid AA1305#18 containing nef alleles and a plasmid encoding the envelope glycoprotein from VSV-G using BioT transfection reagent (Bioland, Paramount, CA).
Western blot
HIV-1 nef genes from the pediatric isolates (PI) were cloned into a plasmid containing the NL4-3 genome (pNL4-3) to make infectious virus as described previously. 41 To construct NL4-3/PI-nef, the nef gene of NL4-3 was replaced with either individual nef obtained from Topo cloning or quasispecies or nef obtained from the nested PCR as described above. T1 cells were infected with NL4-3 virus containing nef alleles from either wild-type NL4-3 or alleles from the pediatric isolates (multiplicity of infection [MOI] = 1). On day 5, lysates harvested from the infected cells were run onto 10% polyacrylamide gel electrophoresis (PAGE) for protein separation. Following transfer to a PVDF membrane (Millipore, Billerica, MA), HIV-1 proteins were detected by an anti-Nef antibody (Nef#2949; NIH AIDS Reagent Program) or an anti-Gag antibody (Gag#6457; NIH AIDS Reagent Program) followed by detection using the SuperSignal West Pico Chemiluminescent Substrate (Thermo Scientific, Rockford, IL).
Immunophenotyping
Surface expression of MHC-I and CD4 in HIV-1-infected and uninfected cells was done using flow cytometry. Study of MHC-I downregulation by Nef on infected thymocytes 40,42 with FITC- and PE-conjugated monoclonal antibodies was done as previously described. 42 –45 PE-conjugated or FITC-conjugated KC57 mAb (Beckman Coulter, Brea, CA) was used to determine intracellular HIV-1Gag protein expression in distinct thymocyte subsets 43,46 and was performed following surface immunophenotyping. 44,45 Study of CD4 downregulation by Nef on infected cells was described previously. 40 Briefly, 2x106 T1 cells were infected with 100 μL (10 ng p24) of viral stock. Three days postinfection, surface expression of CD4 and mCD24 was detected with PE-conjugated human anti-CD4 (BioLegend, San Diego, CA) and FITC-conjugated mouse anti-CD24 (BioLegend). Multiparameter data acquisition was done on a dual-laser FACS Calibur flow cytometer (BDIS, San Jose, CA) or an MACSQuant VYB, and data analysis was performed with FCS Express 6.0.
In vitro infection of postnatal thymocytes
Normal human postnatal thymus specimens were obtained from infants and children undergoing corrective cardiac surgery. The tissue was cut into small pieces, red blood cells were removed by NH4Cl/Tris lysis buffer, and the mechanically disrupted tissue was passed over a cell strainer to generate a single-cell suspension of thymocytes as previously described. 39,44,46 Thymocytes were infected in vitro with CXCR4-tropic pediatric isolates (PI-2 and PI-2.1), the CXCR4-tropic molecular clone NL4-3, the CCR5-tropic molecular clone NF-NSX, or mock infected with the supernatant from the same CD4+ T cell pool used for virus stock preparation. The thymocytes were cultured in serum-free medium for 14 days with IL-2 and IL-4 as previously described. 32 All thymocyte infections with pediatric isolates were standardized by using equal numbers of virus infectious units/107 cells. HIV-1 virion production was measured by HIV-1 Gag p24 enzyme-linked immunosorbent assay as previously described. 44
In vivo infection of SCID-hu mice
C.B-17 SCID mice were bred at UCLA and implanted with human fetal thymus and liver graft (thy/liv) under the murine kidney capsule. 47 Four to six months postimplantation, the thy/liv grafts of SCID-hu mice implanted with thy/liv grafts from the same donor were infected with different HIV-1 isolates (PI-2, PI-2.1, NL4-3, or NFN-SX) by injecting 10–30 ng of p24 in a 50-μL volume directly into the graft as previously described. 44 Mock-infected implants, used as controls in the experiments, were prepared by injecting implants with 50 μL of appropriate control supernatants. Two different experiments consisting of 10–12 SCID-hu mice per 3-week time point were conducted. MHC-I and CD4 phenotyping in HIV-1-infected cells was done at 3 weeks postinfection as previously described. 45
Results
PI-2 is less cytopathic to thymocytes in vitro than PI-2.1 or NL4-3
HIV-1 strains isolated from in utero and perinatally infected children vary in rates of disease progression and exhibit differential effects on thymopoiesis during in vitro culture. 6,32 Beyond the observation that CXCR4-tropic HIV isolates cause more thymocyte depletion than CCR5-tropic HIV isolates, 48 it is unclear what molecular mechanisms determine differential effects on thymocyte depletion and thymic development by HIV-1. Two CXCR4-tropic HIV-1 isolates (PI-2 and PI-2.1) were isolated from an in utero-infected infant and were found to infect CD4+ thymocytes differently. 32 Primary human thymocytes were infected in vitro with PI-2, PI-2.1, or NL4-3 viruses in parallel with a mock-infection control and cultured in serum-free medium supplemented with IL-2 and IL-4 for 14 days. Flow cytometric analysis of surface CD4 and CD8 expression was done on the thymocytes to measure subset depletion. Apoptotic and dead cells were excluded using 7-AAD, and quadrants were set using isotype controls. Following in vitro infection of human thymocytes, PI-2 was less efficient in depleting CD4+ thymocytes (9% CD4+ depletion) compared with PI-2.1 (40% CD4+ depletion) or the CXCR4-tropic molecular clone NL4-3 (90% CD4+ depletion) as related to the percentage of CD4+ cells in mock-infected thymocytes (Fig. 1A). Despite initial infection with equal infectious units, PI-2 in vitro replication was also lower than PI-2.1 as measured by p24 levels in the culture supernatants (Fig. 1B).

Thymocytes infected with the CXCR4-tropic HIV isolate PI-2 in vitro show less CD4 depletion and lower HIV-Gag p24 levels than thymocytes infected with PI-2.1 or NL4-3. Thymocytes were infected in vitro with CXCR4-tropic pediatric isolates (PI-2 and PI-2.1), the CXCR4-tropic NL4-3 molecular clone NL4-3, or mock infected with the supernatant from the same cells (CD4+ T cell pool).
PI-2 contains amino acid mutations in the Nef protein
The Nef protein was previously shown to affect the pathogenicity of HIV-1 in cultured thymocytes 17 and the thy/liv implant in the SCID-hu mouse model. 49 Sequencing of the nef alleles found in PI-2 and PI-2.1 was done to examine a possible cause of their discrepancy in thymic cytopathicity. Analysis of the two nef sequences revealed that the amino acids (aa) of these two isolates were highly similar, and the majority of reported functional motifs were conserved in both isolates (Fig. 2A). The myristoylation residue (G2) required for membrane localization, the oligomerization domain needed for all functions, and motifs such as WL58, LL165, DD175, and EE155, which are required for CD4 binding and lysosomal degradation function, were found to be preserved in both isolates. However, a deletion of seven amino acids (NefΔK7S) from amino acid positions 82 to 88 (82KGALDLS88) was present in PI-2 but not in PI-2.1. This sequence falls within the core domain of Nef (aa 72–203), which is a relatively conserved and structured type II polyproline helix (aa 72–77) followed by two α-helices (aa 81–120). 50 This novel deletion is also 4 amino acids downstream of a polyproline helix (72PXXPXR/K77) binding site for Src-homology domain 3 (SH3) that mediates cellular activation and MHC-I downregulation. 15,29 In addition to NefΔK7S, a single amino acid mutation at position 71 from K to R (K71R), adjacent to the SH3 binding site, was observed in PI-2.
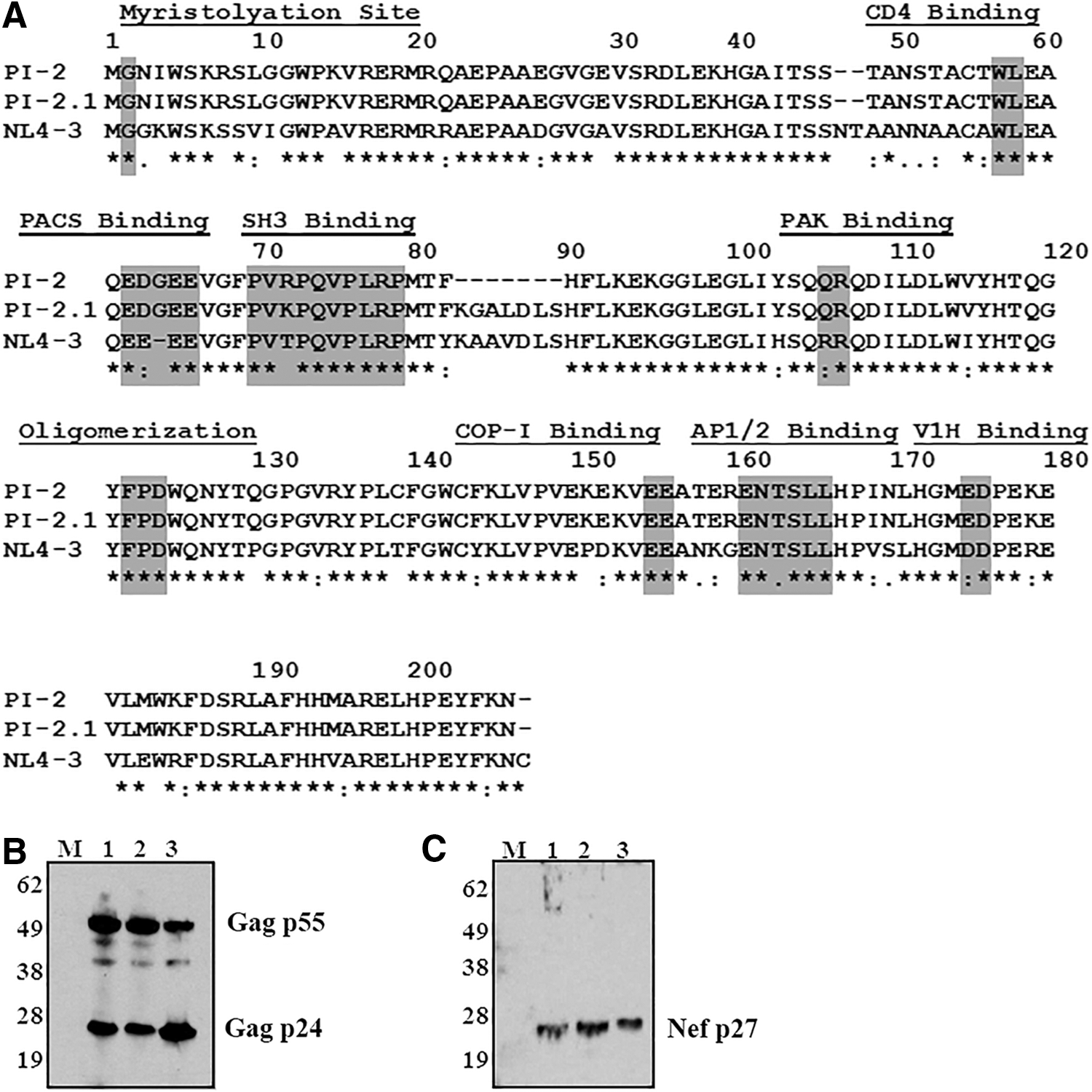
Nef mutations from HIV isolate PI-2 do not inhibit the expression of structural proteins.
Nef mutations in PI-2 do not inhibit expression of structural proteins
To assess if the identified mutations in the nef gene of PI-2 inhibited Nef protein expression, Western blot analysis was done on cells infected with CXCR4-tropic molecular clone NL4-3 (lane 1), CCR5-tropic pediatric isolate PI-832 (lane 2), and CXCR4-tropic isolate PI-2 (lane 3). The results show that PI-2-infected cells expressed similar levels of HIV-1 Gag (Fig. 2B) and Nef (Fig. 2C) as cells infected with PI-8 or NL4-3 and used as controls. Accordingly, PI-2-infected cells express HIV-1 Nef despite mutations.
PI-2 is impaired in MHC-I downregulation in vitro and in vivo
While a myriad of cellular effects have been shown for Nef, two of its most clearly defined functions are to downregulate cell surface expression of MHC-I and CD4 molecules. These functions were assessed in thymocyte cultures infected with PI-2 and PI-2.1 and compared with cultures infected with the HIV-1 molecular clones NL4-3 or NFN-SX (NL4-3 with a substituted CCR5-tropic Env from strain JR-FL). Mock-infected cultures were prepared by adding supernatants of uninfected activated allogeneic CD4+ T cell pools. Thymocytes infected in vitro (Fig. 3) were analyzed for surface expression of MHC-I. Following infection, MHC-I downregulation was assessed by the mean fluorescence intensity (MFI) of MHC-I in HIV-1 Gag+ cells (KC57+). The cells were surface stained with antibodies to MHC-I (HLA-ABC-FITC) and CD3-APC followed with intracellular staining for HIV-1 Gag proteins using the KC57-PE antibody. Isotype controls were used to set the quadrants. MHC-I expression on mock-infected thymocytes is shown in Figure 3A. Cells productively infected (KC57+) with CXCR4-tropic NL4-3 containing a functional Nef (Fig. 3B) had decreased expression of MHC-I (MFI = 263) compared with uninfected (KC57−) cells (MFI = 745). Cells infected with the CCR5-tropic virus NF-NSX containing a functional Nef (Fig. 3C) had a decreased expression of MHC-I (MFI = 217) compared with uninfected cells (MFI = 551). Cells infected with the pediatric isolate PI-2.1 containing a functional Nef (Fig. 3D) also had decreased MHC-I expression (MFI = 220) compared with uninfected cells (MFI = 639). However, cells infected with the pediatric isolate PI-2 containing the Nef mutation (Fig. 3E) showed a smaller decrease in MHC-I surface expression (MFI = 417) compared with uninfected cells (MFI = 573).

Pediatric HIV isolate 2 (PI-2) is impaired in MHC-I downregulation in vitro. Thymocytes were infected in vitro with CXCR4-tropic pediatric isolates (PI-2 and PI-2.1), the CXCR4-tropic NL4-3 and CCR5-tropic NF-NSX molecularly cloned HIV-1 isolates, or mock infected with the supernatant from the same cells (CD4+ T cell pool).
The ability of the pediatric isolate Nef proteins to downregulate MHC-I was also tested in vivo using an SCID-hu mouse model implanted with human fetal thy/liv tissue that forms a thymus-like organoid as previously described. 44,45 SCID-hu mice were infected with NL4-3, PI-2, PI-2.1, or mock-negative control supernatant. MHC-I downregulation in productively infected (KC57+) thymocytes was measured ex vivo at 3 weeks postinfection by measuring the HLA-ABC MFI. As shown in Figure 4A, cells infected with NL4-3 (n = 5 mice) and PI-2.1 (n = 5 mice) showed decreased mean surface expression of HLA-ABC compared with cells infected with PI-2 (n = 6 mice). PI-2 expressing thymocytes had similar levels of MHC-I expression as mock infection (n = 5 mice). Statistical analysis was done using two-tailed unpaired t tests. Similar to the in vitro experiments, infections of thy/liv implants with PI-2, PI-2.1, or NL4-3 again demonstrated diminished capabilities of PI-2 to downregulate MHC-I following infection, whereas PI-2.1 exhibited comparable MHC-I downregulation with NL4-3 (Fig. 4A). The percentages of CD4+ expression on thymocytes in thy/liv implants following infection with PI-2 were the same as in mock-infected implants, contrary to our earlier in vitro experiments. However, PI-2.1-infected implants showed a similar decrease in the percentage of CD4+ thymocytes as NL4-3-infected implants (Fig. 4B).
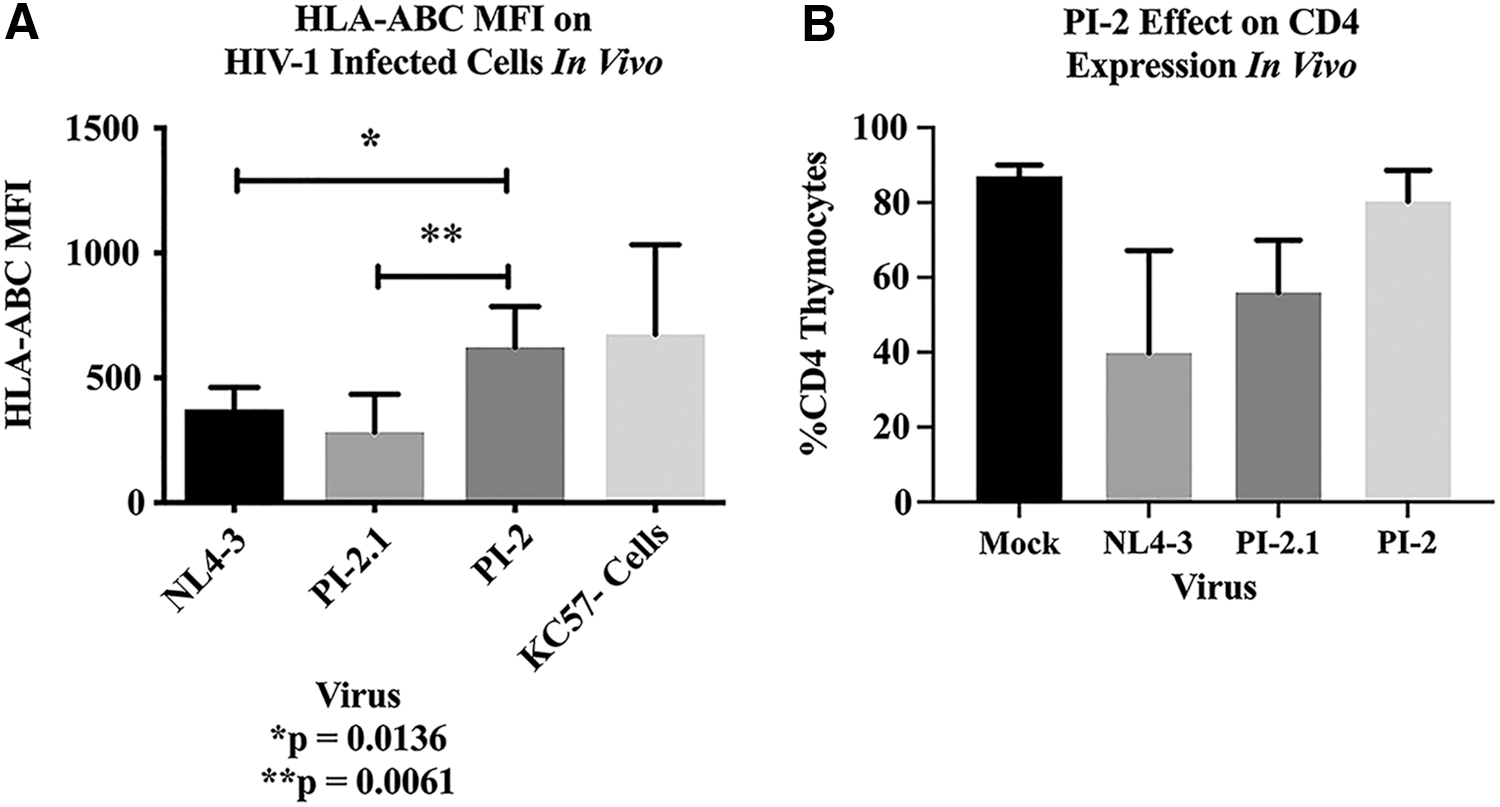
Pediatric HIV isolate 2 (PI-2) is impaired in MHC-I downregulation in vivo, but is less cytopathic than PI-2.1. SCID mice implanted with human fetal thy/liv (SCID-hu mice) were infected with NL4-3 or pediatric isolates PI-2.1 and PI-2.
HIV-1 PI-2 Nef downregulates CD4 but not MHC-I expression in the absence of Vpu and Env in vitro
To investigate the small discrepancy in CD4 downregulation between our in vitro and in vivo experiments, we analyzed the Nef proteins separately from other HIV-1 proteins, Env and Vpu, which have previously been reported to contribute to CD4 surface downregulation. 51,52 To test the effect of Nef separately from Env and Vpu while maintaining their physiologic expression levels, PI-2 and PI-2.1 nef genes were inserted into an NL4-3-based genome containing deletions of env and vpu. 40 Single-cycle infection was done using VSV-G pseudotyped viruses expressing a CD24 tag and the various nef genes. In agreement with our earlier in vitro data obtained in thymocytes infected with the original pediatric isolates, PI-2 Nef was unable to downregulate MHC-I in infected cells as much as Nef from PI-2.1 and NL4-3 (Fig. 5A–E). Confirming our previous experiment, PI-2 Nef was still able to downregulate CD4 surface expression in vitro, even in the absence of Vpu and Env (Fig. 5F–J).
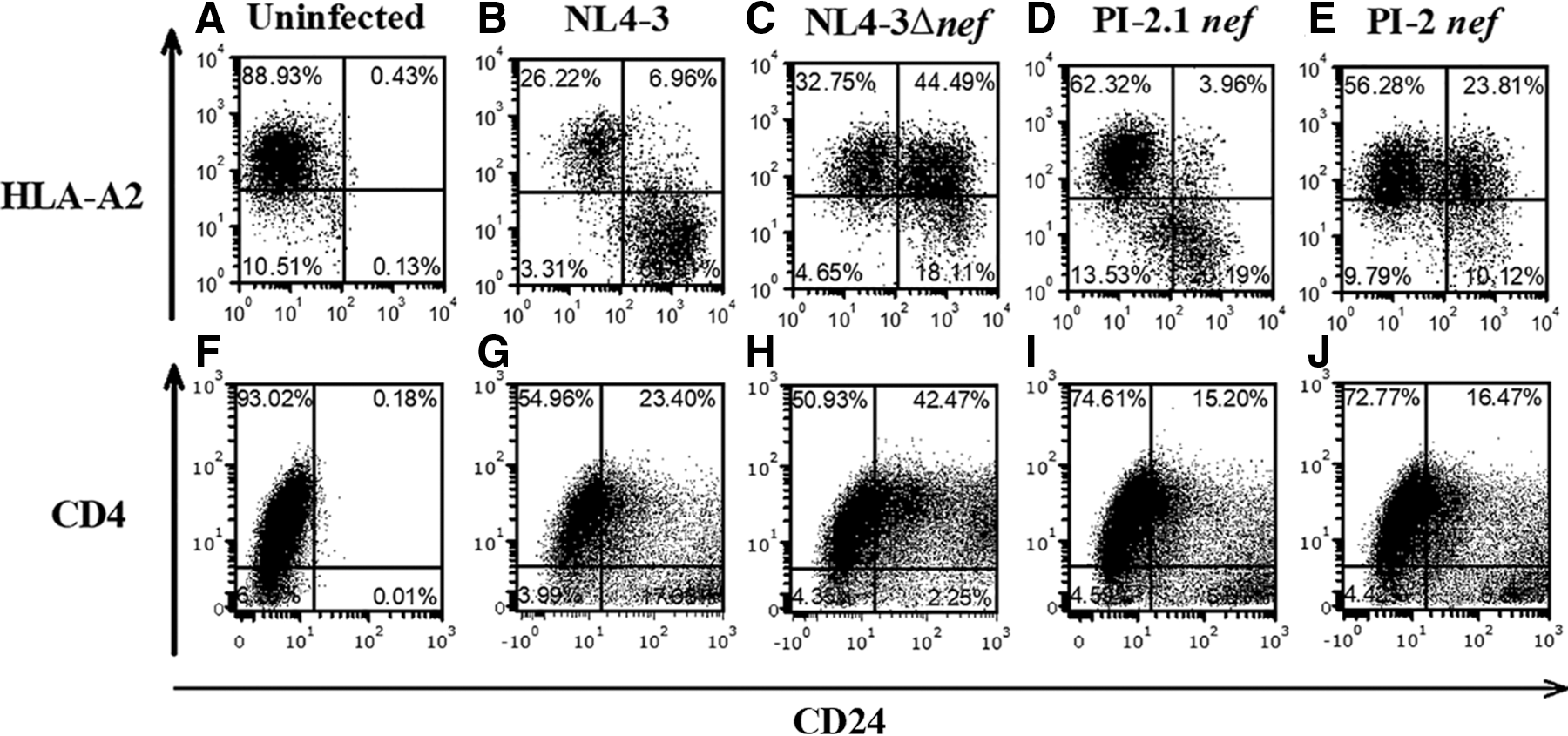
HIV PI-2 Nef downregulates CD4 but not MHC-I expression in the absence of Vpu, Env, and Vpr. To assess the MHC-class I downregulation of the nef genes in pediatric isolates PI-2.1 and PI-2 in the absence of Vpu, Env, and Vpr proteins, the nef genes were cloned out of the PI-2.1, PI-2, and NL4-3 isolates and placed in an NL4-3 vector lacking Vpu, Env, and Vpr. To determine the effect of PI-2.1 and PI-2 Nef proteins on MHC-I and CD4 downregulation, VSV-G pseudotyped virus, including a CD24 tag and the various nef genes, was used to infect T1 cells. HLA-A2 and CD4 surface expression in T1 cells infected with
Discussion
Following comparison of two primary HIV-1 pediatric isolates from the same in utero-infected infant, PI-2 and PI-2.1, we identified a unique in-frame deletion NefΔK7S in PI-2, which attenuates the depletion of thymocytes in vitro. Nef proteins bearing this mutation are defective for MHC-I downregulation, but this effect is not due to reduced Nef expression or global Nef dysfunction. Nef proteins bearing this mutation were still able to downregulate CD4 surface expression in vitro. This novel ΔK7S in-frame deletion is in a location that is not known to contain any specific motifs required for previously defined Nef functions. However, it is adjacent to the SH3 binding site that is critical for multiple Nef functions, including cellular activation and MHC-I downregulation. 53,54 It is possible that ΔK7S alters the tertiary structure of Nef and thus interferes with interactions with host factors. While there is a K versus R polymorphism that falls within the PXXP motif between residues 72–75 when comparing PI-2.1 with PI-2, this is unlikely to contribute to impaired MHC-I downregulation by PI-2. Although the NL4-3 Nef sequence has T71 (Fig. 2A), the subtype B consensus sequence is actually 71R, and thus, PI-2 actually matches “wild-type” Nef more closely than PI-2.1 or NL4-3 at position 71. Moreover, K71R is a conservative substitution. Although highly unlikely, however, we cannot definitively rule out that 71R somehow contributed to the impaired MHC-I downregulation by PI-2.
Previous data show that various mutations in Nef affect oligomerization as well as ternary complex formation with cell molecules such as AP-1 and MHC-I, 50,55 –57 and a survey of in vivo Nef sequences demonstrated frequent examples of Nef with impaired MHC-I downregulation despite no identifiable mutations in the defined motifs involved in this function. 58 Similarly, the significance of the K71R substitution mutation in PI-2 Nef is unclear. This mutation is also located in proximity to the canonical sequence required for SH3 binding. Mutational studies have demonstrated that residues flanking the motif play critical role in determining binding specificity to SH3 domain. 59
Others have reported that Nef-mediated CD4 downregulation correlates with increased viral pathogenesis in thymocytes, and MHC-I downregulation has diminished influence in cytopathicity. 60 In contrast, our findings indicate that PI-2-mutated nef containing virus was less cytotoxic in thymocytes than PI-2.1, despite having a similar ability to downregulate surface expression of CD4 in vitro. PI-2-infected cells did not downregulate MHC-I but did decrease CD4 surface expression in vitro, indicating that the novel identified mutations in PI-2 Nef may not have affected binding to several host factors reported to be involved in CD4 downregulation. 61,62 Point mutations in the nef gene (WL57AA, LL164AA, and DD174AA) of the NL4-3 molecular clone abrogate CD4 downregulation and decrease Nef-mediated pathogenesis. 60 These mutations were not present in PI-2. Van Nuffel et al. have previously reported that Nef:PAK2 and an intact VGF motif within Nef are required for reduced thymic output. 63 Despite having intact VGF and PAK2 binding motifs and representative CD4 downregulation in vitro, PI-2 still lacked the virulence and thymic depletion seen in thymocytes infected with PI-2.1 or molecular clones, indicating a correlation between MHC-I downregulation and Nef's ability to deplete thymocytes.
Given the importance of MHC-I molecules in T cell development, it is unclear whether the defect in MHC-I downregulation by HIV isolate PI-2 is directly related to the attenuated thymic cytopathicity, or if the novel identified mutations disrupt another pathway that potentiates the negative effect of Nef in the thymopoiesis. Further unraveling this phenomenon may provide greater insight into the particularly rapid disease progression seen in HIV-1-infected children, and may offer therapeutic targets for this particularly vulnerable population.
Sequence Data
The nucleotide sequences for PI-2 and PI-2.1 nef genes have been deposited in GenBank under accession numbers KC702883 and KC702882, respectively.
Footnotes
Acknowledgments
The following reagents were obtained from the NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH: anti-Nef antibody (Nef#2949), anti-Gag antibody (Gag#6457), and the HIV-1 NL4-3 infectious molecular clone (pNL4-3) from Dr. Malcolm Martin (cat. no. 114). 38 The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Authors' Contributions
A.A., L.P.-M., R.L.F., O.O.Y., and C.H.U. conceived and designed the experiments; A.A., L.P.-M., A.D.C., and D.A.-P. performed the experiments; A.A., L.P.-M., R.L.F., A.D.C., D.A.-P., and C.H.U. analyzed the data; Y.B. and O.O.Y. contributed reagents/materials/analysis tools and advice; A.A., R.L.F., O.O.Y., and C.H.U. wrote the article.
Author Disclosure Statement
The authors do not have any financial or nonfinancial competing interests.
Funding Information
This work was supported by the National Institutes of Health, grant numbers: HD29341, AI054286, AI32440, and CA016042. This work was also supported by the UCLA AIDS Institute, UCLA Center for AIDS Research (CFAR) NIH AI28697, James B. Pendleton Charitable Trust, and the McCarthy Family Foundation.