
Abstract
In this study we report on the identification of a simian immunodeficiency virus (SIV) infecting a Chlorocebus tantalus from Cameroon. The isolate, SIVagmTAN-CA1, was molecularly characterized by sequencing partial genome (∼4,000 bp) using the conventional Sanger method and the Oxford Nanopore Technology (ONT). In pol and gp41/nef SIVagmTAN-CA1 clusters with SIVagmSAB infecting Chlorocebus sabaeus from West Africa, whereas in env-gp120 it clusters with SIVagmTAN infecting C. tantalus from Central Africa. This mosaic structure is similar to that of a previously reported isolate infecting another tantalus monkey from Cameroon and confirms that the evolution of SIVagm is complex. Our data show that ONT sequencing gives results comparable with conventional Sanger sequencing on SIV and could help in distinguishing recombination and coinfection.
At least 45 non-human primate (NHP) species are naturally infected by simian immunodeficiency viruses (SIVs) in Africa. 1 SIVs infecting chimpanzees, gorillas, and sooty mangabeys are known to be the precursors of human immune deficiency viruses (HIV-1 and HIV-2). 2 –4 The genetic diversity and evolution of SIVs are complex. This diversity provides examples of coevolution between hosts and viruses, recombination between different SIV lineages, interspecies transmission and cocirculation of different viruses in the same species. 5 However, in a majority of cases, each NHP species is infected by a species-specific SIV lineage.
African green monkeys (AGMs) belong to the genus Chlorocebus, which is divided into six species distributed throughout equatorial Africa. 6 They are the most numerous and widespread monkeys in Africa, living mainly in wooded savannah close to rivers or lakes. The six species are geographically separated, with possible contacts in border areas. SIVs have been detected in five of them: SIVagmSAB in Chlorocebus sabaeus, from Senegal to the Volta River, SIVagmTAN in Chlorocebus tantalus from Ghana through Cameroon to Sudan, SIVagmGRI in Chlorocebus aethiops from East Africa, SIVagmVER in Chlorocebus pygerythrus from Southern and East Africa, and SIVagmMAL in Chlorocebus cynosuros from Zambia and Namibia. To our knowledge, there is no evidence of SIV infection for Chlorocebus djamdjamensis that inhabits in Ethiopia, but only few exemplars have been tested.
SIV serological testing shows that ∼50% of wild AGM population are infected. AGMs thus represent a large SIV reservoir. 7 Previous findings suggested virus/host coevolution between AGMs and SIVagm. 8 However, this hypothesis has been challenged by the identification of a recombinant SIVagm (SIVagmTAN-CM545) in a tantalus monkey from Cameroon that underscores that interspecies transmission and recombination also probably drive SIVagm evolution. 9 For instance, a large part of SIVagmTAN-CM545 (gag, pol, vif, nef) resembles SIVagmSAB, infecting C. sabaeus from West Africa (Senegal), and the remaining part of the genome resembles SIVagmTAN, infecting a tantalus monkey from Uganda. To our knowledge, there is no information on the extent of recombination and distribution of recombinant in monkeys from Cameroon, and it is unclear if recombinants actually resemble ancestral SIVs. To help fill these gaps, we analyzed another SIV-positive tantalus monkey from Cameroon. Our data show a novel mosaic SIVagmTAN with parts resembling SIVagmSAB and parts resembling SIVagmTAN.
During field surveys on SIV diversity and the origins of HIV-1, we collected AGM samples from bushmeat (n = 14) and from pet monkeys (n = 4) in Cameroon. Polymerase chain reaction (PCR) amplification and sequencing of a 1,200 bp mitochondrial DNA fragment coding for cytochrome-b showed that 17 were C. tantalus and one was from a Cercopithecus species. Serological SIV testing with INNO-LIA showed that 2 (S95 and CA1) of the 17 AGM had HIV cross-reactive antibodies. CA1 was further confirmed by a SIV peptide-based Luminex assay, 10 with highest reactivity observed against SIVagm, SIVsmm, and HIV-2. AGM-CA1 was an adult male sampled in 1998 in southwest of Cameroon, in Limbe, where he was kept as pet. For the sample S95, there was no plasma left for SIV-Luminex assay testing.
To confirm the presence of SIV virus, we extracted viral RNA from CA1 plasma using Biomérieux MiniMag Kit and generated cDNA using GoScript RT (Promega, France). Diagnostic PCR using universal HIV/SIV primers targeting a 400 bp fragment in the integrase region of pol confirmed the SIV positive status of CA1.
We next amplified a 1,800 bp fragment in pol (position relative to SIVmac239: 3,233 → 5,072) and 2,300 bp (1,200, 700, and 400 bp fragments) in env/nef using three overlapping nested PCRs (position relative to SIVmac239: 6,840 → 9,194). All the amplified fragments were sequenced using cycle sequencing and dye terminator methodologies (ABI PRISM Big Dye Terminator Cycle sequencing Ready Reaction kit with amplitaq FS DNA polymerase) on an automated sequencer (ABI 3500; Applied Biosystems, Courtaboeuf, France). Because Sanger sequencing enables the detection of variants present only in at least 20% of the viral population, we also sequenced three PCR amplicons (pol 1,800 bp, env/gp120 1,200 bp, and gp41/nef 400 bp) with a MinION (Oxford Nanopore Technology [ONT], UK) using Native Barcoding kit SQK-LSK109 to detect eventual minority SIV variants. Basecalling and corrections were performed with SeqManPro (DNASTAR, Madison, WI) for Sanger sequences, and with an updated in-house pipeline for ONT reads as described previously. 11 Corrected consensus sequences were phylogenetically analyzed using PhyML3.1, whereas recombination analysis was performed with SimPlot3.5.1 and RDP4 software on concatenated sequences.
Structural analysis of the translated generated sequences showed no stop codon. A total of 15 out of 18 highly conserved cysteine residues in 3′ were observed in env. The first three cysteines are probably present in the unamplified sequence in env/gp120, between pol and env genes. The V3 loop in CA1, like in CM545 or the other SIVagm lineages known today, was 34 amino acids long and the M-A-G triplet was found on the crown. A total of 19 potential N-glycosylation sites (N-X-S/T) were found, 12 of which are common to all known SIVagm lineages.
Phylogenetic analysis of partial fragments obtained by both Sanger and ONT sequences in pol (1,800 bp), env-gp120 (1,200 bp), and env-gp41/nef (400 bp) showed differential clustering of SIVagmTAN-CA1 with other SIVagm lineages. In pol, SIVagmTAN-CA1 clustered with SIVagmSAB infecting West African sabaeus green monkeys (Fig. 1A). In env/gp120, SIVagmTAN-CA1 clustered with SIVagmTAN infecting tantalus monkey from Uganda (Fig. 1B) and in env-gp41/nef, SIVagmTAN-CA1 again clustered with SIVagmSAB (Fig. 1C). Consensus sequences obtained by both sequencing methods showed similarities >99.5% (translated amino acid sequences were identical). Since distinct SIV lineages have been identified in some NHP's species (e.g., SIVmand1 and SIVmand2 in Mandrillus leucophaeus or SIVmus1, SIVmus2, and SIVmus3 in Cercopithecus cephus), we wanted to exclude the possibility of coinfection by two distinct lineages of SIVagm in this tantalus monkey. The MinION sequencing performed on three critical genomic regions here excluded this possibility of coinfection and rather, confirmed the mosaic nature of this novel SIVagm-CA1.

Phylogeny of SIVagmTAN-CA1 sequences obtained by ONT and Sanger sequencing technologies. The figure shows phylogenetic trees of SIVagmTAN-CA1 together with other SIV lineages. The phylogenies were inferred by maximum likelihood, using the PhyML software and the GTR+G+I substitution model.
We concatenated the different fragments obtained by Sanger sequencing to perform a recombination analysis of this 4,100 bp long construct. This analysis confirmed the mosaic structure of SIVagmTAN-CA1, very similar to that of SIVagmTAN-CM545 (Fig. 2). The pattern observed with the SimPlot software was confirmed with RDP4 software (not shown).
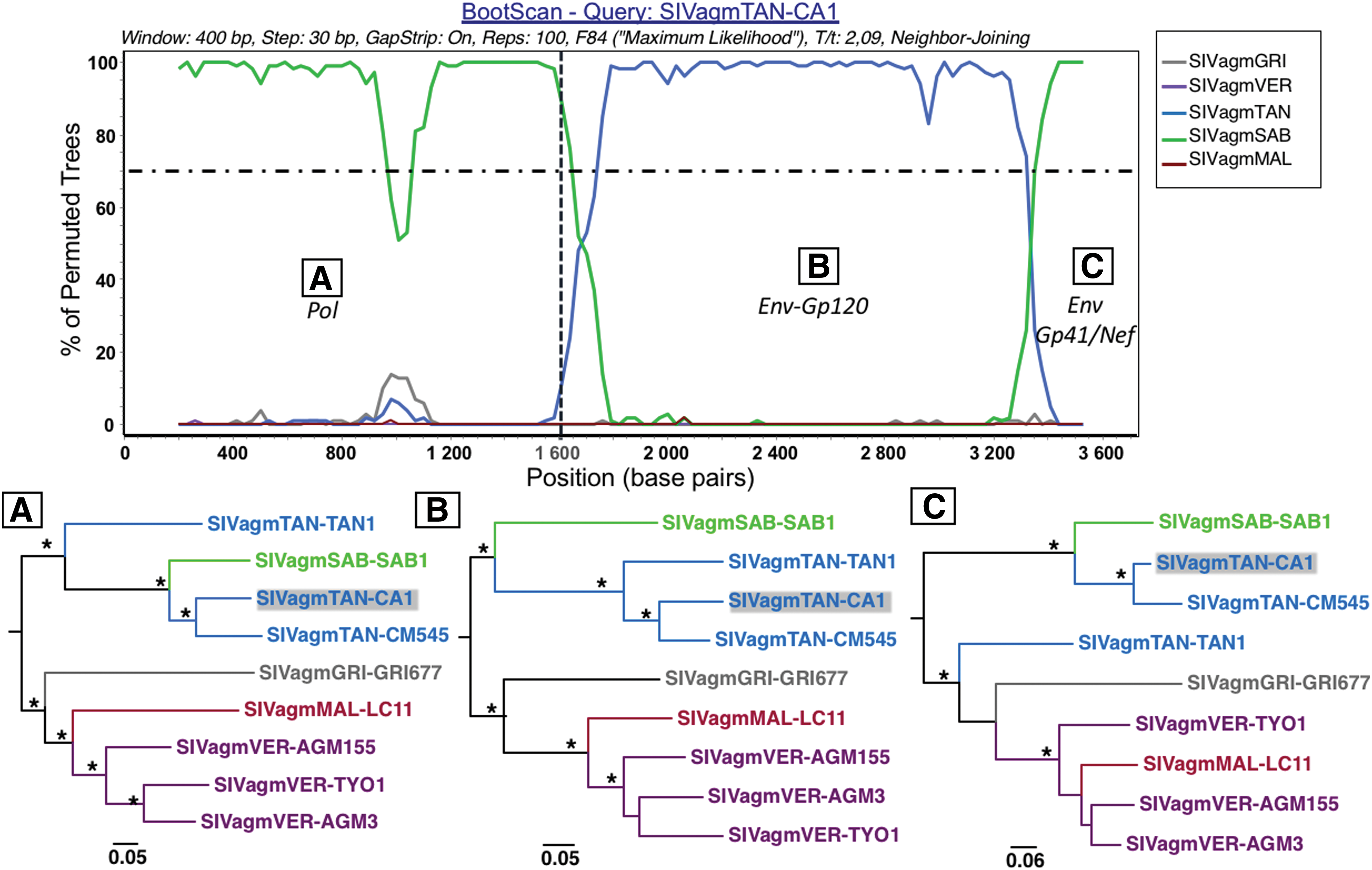
Mosaic structure of concatenated sequence of SIVagmTAN-CA1. Recombination pattern in SIVagmTAN-CA1 was searched by concatenating sequences obtained by the Sanger method and representing a ∼4,100 bp fragment. Bootscan analysis using the SimPlot software was conducted on alignment comprising the five lineages of SIVagm in addition to the novel SIVagmTAN-CA1. The pol region covers the positions 1–1,600 bp (vertical dotted line) and then env and nef regions thereafter. In pol and env-gp41/nef, the closest lineage is SIVagmSAB (green) and in env-gp120, SIVagmTAN (blue). This clustering is confirmed by the phylogeny derived from corresponding alignment slices
This study reports on a partial molecular characterization of an SIV infecting a tantalus monkey from Cameroon. The new SIV isolate, SIVagmTAN-CA1, presents a mosaic structure very similar to that of SIVagmTAN-CM545 infecting another tantalus monkey also from Cameroon. 9 These findings underscore that the dynamics and evolution of SIVagm is more complex than postulated before. Thorough investigations on genetics of Central African AGMs and of SIV infecting them, as well as from more individuals of the other AGM species and their SIVs are thus necessary. In this study, we also showed for the first time on SIV that ONT sequencing gives very comparable results to conventional Sanger sequencing and helped distinguishing recombination from potential coinfection with distinct SIVagm viruses.
Accession Numbers
The sequences generated in this study were deposited in the GenBank under the accession nos. MN315553–MN315557.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by IRD and by Agence Nationale de Recherches sur le SIDA et les Hépatites virales (ANRS) (grant nos. ANRS-12125, ANRS-12182, and ANRS-12325).