
Abstract
Although antiretroviral therapy (ART) effectively suppresses HIV replication, the latent reservoir remains the barrier to HIV eradication. It remains unknown whether long-term ART impacts levels of inducible replication-competent provirus. To address this knowledge gap, we assessed the proviral reservoir in HIV-1 perinatally infected adolescents having received ART for >13 years. We recruited 15 vertically infected adolescents living with HIV in Botswana. Historical viral load, CD4+ T cell count, and treatment data were retrieved from their outpatient medical records. Inducible replication-competent proviruses from cryopreserved peripheral blood mononuclear cells were quantified using a TZM-bl based assay (TZA). Total proviral DNA copies were quantified using droplet digital PCR. The mean age of study participants was 16 years (standard deviation = 0.7) and median CD4+ T cell count at enrollment was 784 [interquartile range (IQR) = 728.8–1,288] cells/mm3. Median age at ART initiation was 8 (IQR = 6–12) months. Fourteen (93%) participants had HIV-1 RNA <400 copies/mL at the time of enrollment in the study. A median of 19 (IQR = 18–27) HIV-1 RNA measurements were available per participant. Six (40%) participants displayed viral suppression at all clinic visits since initiating ART, whereas the remaining 9 (60%) had one or more clinic visits with detectable HIV-1 RNA. The median inducible replication-competent provirus count was 7.4 infectious units per million cells (IQR = 6.7–19.2), and did not differ significantly by either complete or incomplete viral suppression (7.2 vs. 7.4, p = .86), or by age at ART initiation (7.4 if <12 months, 11.2 if >12 months, p = .85). The median total HIV DNA count was 129.1 copies per million cells (IQR = 18.9–212.3). Our data suggest that long-term ART initiated within the 1st year in perinatally infected infants did not eliminate proviral DNA or inducible replication-competent proviruses.
Introduction
Recent findings suggest that early initiation of antiretroviral therapy (ART) limits the size of the reservoir by restricting HIV expansion into CD4+ T cells, and prolongs time to viral rebound to >100 days after ART interruption. 1,2 Although the majority of inducible reservoir assessment studies have been conducted on HIV-1-infected treatment experienced adults, no studies have quantified the inducible reservoir in HIV-1-infected pediatric patients receiving long-term ART since infancy. In Botswana, like many other high burden countries, a significant number of perinatally infected patients have received long-term ART and are virologically suppressed. Yet it is not known whether early and long-term viral suppression impacts the size or activation potential of the viral reservoir. Furthermore, most studies of the HIV reservoir have been conducted in HIV-1 subtype B- (HIV-1B) infected individuals 3 and there is need to assess the inducible replication-competent reservoir in HIV-1 non-B subtype infections, which reflects a substantial burden of HIV infection globally.
ART leads to successful viral suppression and prolongs life expectancy of people living with HIV, but it does not eliminate the virus from the body. Viral rebound can occur if ART is interrupted or terminated as a result of the activation or induction of the HIV reservoir. 4 –8 Upon induction, latently infected cells could produce replication-competent viruses. Reservoir size is presumed to be a major determinant for the extent of HIV replication after the cessation of ART. Although latent proviruses are present in CD4+ T cells, they are predominantly found in resting CD4+ (rCD4+) T cells, which account for only 1%–5% of the HIV-infected CD4+ T cell population in untreated patients. 9 –11 The low frequency of latently infected cells presents a challenge in determining the true size of the reservoir.
At present, a number of assays are used to measure the size of the reservoir. Some assays, such as droplet digital PCR (ddPCR), quantify total HIV DNA copies; these assays tend to overestimate the size of the reservoir because they do not distinguish between intact and defective viruses. 12 –15 Measuring the number of intact and/or replication-competent proviruses is a more accurate way of quantifying the reservoir. A recently developed reporter cell-based assay, the TZM-bl-based assay (TZA), measures inducible replication-competent proviral reservoir, and is sensitive, precise, and inexpensive. 14,16
Botswana is one of the countries in sub-Saharan Africa that has experienced high burden of HIV-1 subtype C (HIV-1C) infection. HIV-infected adolescents in Botswana are enrolled in the National ART program and are routinely monitored by viral load testing. It is unknown whether long-term viral suppression in adolescents who received ART since infancy can result in lower levels of inducible replication-competent proviruses, and whether their reservoir sizes differ from adults. It remains unknown whether receiving prophylaxis through the prevention of mother-to-child transmission might affect the reservoir size in perinatally infected infants.
We sought to evaluate the impact of long-term ART and sustained viral suppression on inducible replication-competent proviruses in adolescents perinatally infected with HIV-1C in Botswana. We used the reporter cell-based TZA assay to quantify inducible replication-competent proviruses in HIV-1C-infected adolescents, 14,16 in combination with ddPCR (Bio-Rad) to quantify total HIV DNA.
Materials and Methods
Study population
Fifteen perinatally infected adolescents with HIV-1C and receiving ART who had formerly participated in the “Mashi Study” (
In 2018, Mashi Study mothers or guardians were recontacted for potential enrollment of the adolescents in this study. At enrollment, HIV antibodies were tested by Determine HIV-1/2 (Abbott Laboratories, Abbott Park, IL) and UniGold HIV (Trinity Biotech, Wicklow, Ireland). Viral loads were measured by the Roche Amplicor HIV-1 Monitor Test v1.5 (Roche Molecular Systems, Branchburg, NJ), and CD4+ T cell counts by BD TruCount™ (Becton Dickinson Biosciences, San Jose, CA), HIV diagnosis dates, and ART initiation dates, treatment history were retrieved from Mashi Study records and Botswana clinical records.
Ethical considerations
This study was approved by the Institutional Review Board of the Harvard T.H. Chan School of Public Health (Boston, MA) and the Health Research Development Committee of the Botswana Ministry of Health and Wellness (Gaborone, Botswana). Written informed consent and assent were obtained from parents/guardians and participants, respectively.
Isolation and purification of resting CD4+ T cells
Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood and cryopreserved in heat-inactivated fetal calf serum (FCS) (Gibco, NY) containing 10% dimethyl sulfoxide (Sigma-Aldrich, Inc., Allentown, PA). PBMCs were thawed in R10 medium [RPMI-1640 medium (Sigma-Aldrich, Inc.) supplemented with 10% FCS (Gibco), 1%
Activation of rCD4+ T cells
For activation, the rCD4+ T cells were cultured with anti-human CD3/CD28 Ab-coated microbeads (Dynabeads, Life Technologies, Carlsbad, CA) according to manufacturer's instructions in RPMI-1640 medium (Sigma-Aldrich, Inc.) with 10% FCS (Gibco) in the presence of 300 nM efavirenz (Sigma-Aldrich, Inc.) and recombinant human interleukin (IL)-2 and IL-7 Abs (Miltenyi Biotech). After 6 days in culture, the rCD4+ T cells were washed thoroughly and used in the TZA assay. Aliquots of activated rCD4+ T cells were used for quantification of HIV proviral DNA.
TZM-bl-based assay
The activated rCD4+ T cells were serially diluted fourfold (250–125,000 cells per well, each dilution had 8 replicates) and cocultured with 30,000 TZM-bl cells per well in a 96-well plate, as described by Sanyal et al. 14 and Gupta et al. 16 A control 96-well plate with uninfected rCD4+ T cells was used in parallel with the test plate. The cells were suspended in RPMI-1640 (Sigma-Aldrich, Inc.) containing 10% FCS (Gibco) and cultured at 37°C with 5% CO2 for 72 h. The wells were washed twice with phosphate-buffered saline (Life Technologies), and Beta-Glo reagent (Promega, Madison, WI) was added to the monolayer of TZM-bl cells before incubation for 45 min to 1 h at room temperature in the dark. An illuminometer was used to measure chemiluminescence activity (β-galactosidase expression). In the test plate containing HIV-1-infected cells, a well was considered positive if the chemiluminescence signal exceeded the mean plus two times the standard deviation (mean +2*SD) of the signal observed in the control sample wells.
Calculations of infectious units per million cells
The infectious units per million cells (IUPM) values were quantified by the Siliciano Lab method (Johns Hopkins, Baltimore, MD;
HIV-1 DNA quantification by ddPCR
DNA was extracted from whole PBMCs, rCD4+ T cells before activation, and rCD4+ T cells after activation using the Qiagen Puregene Cell and Tissue kit (Qiagen Sciences, LLC, Louisville, KY) according to the manufacturer's instructions. HIV-1 was amplified using primers and probes, as previously described and listed hereunder
20
: LTRgag primers/probe [1] LTRgagF: 5′-TCTCGACGCAGGACTCG LTRgagR: 5′-TACTGACGCTCTCGCACC LTRgagP: 5′-/56-FAM/CTCTCTCCT/ZEN/TCTAGCCTC/31ABkFQ/ RPP30 primers/probe [2] RPP30F: 5′-GATTTGGACCTGCGAGCG RPP30R: 5′-GCGGCTGTCTCCACAAGT RPP30P: 5′-/56-FAM/CTGACCTGA/ZEN/AGGCTCT/31ABkFQ/
PCR running conditions were as follows: 95°C for 10 min, 45 cycles of 94°C for 30 s, and 60°C for 1 min, followed by 98°C for 10 min, and hold at 4°C. To quantify total HIV copies in each sample cell type, ddPCR expression analysis was performed and the QuantaSoft™ software (Bio-Rad, Hercules, CA) was used to analyze the ddPCR data to determine the total HIV DNA copies per million cells.
Calculation of the reservoir size
The size of the reservoir is indicated by the fraction of provirus that can be stimulated to form replication-competent virus, or the fraction of proviral expression (fPVE). The fPVE was calculated by dividing the IUPM values (determined by TZA) by the number of total HIV DNA copies per million cells (determined by ddPCR) and multiplying by 100, as described previously. 14,16
Results
Of 15 perinatally infected study participants, 64% were of male and 36% female gender, and the age at enrollment ranged from 14 to 17 years (Table 1). All participants had tested positive for HIV-1 within the Mashi Study and initiated ART at a median of 8 [interquartile range (IQR) = 6–12) months. At study enrollment, all participants had been on ART for 13–16 years. Fourteen (93%) were virally suppressed at their latest evaluation before enrollment, whereas 1 had viral load of 2,033 HIV-1 RNA copies/mL. Median CD4+ T cell count was 784 (IQR = 729–1,288) cells/mm3. A median of 19 (IQR = 18–27) HIV-1 RNA measurements per participant over a median of 15 (IQR = 13–15) years were performed. Six (40%) participants were virally suppressed at all visits for >10 years (Supplementary Fig. S1A), whereas 9 (60%) participants whose viral suppression did not exceed 10 years (n = 7 experienced from 1 to 13 incidents of detectable viral load over time, n = 2 had missing data) (Supplementary Fig. S1B). No information on participants' adherence was available.
Clinical Characteristics of Study Participants
3TC, lamivudine; ABC, abacavir; ART, antiretroviral therapy; AZT, zidovudine; d4T, stavudine; DTG, dolutegravir; EFV, efavirenz; FTC, emtricitabine; IQR, interquartile range; LPV, lopinavir; LPVr, lopinavir/ritonavir; N/A, not available; NVP, nevirapine; SD, standard deviation; TDF, tenofovir; VL, viral load.
Use of the TZA assay on cryopreserved PBMCs to assess the reservoir in HIV-1C infection
To assess the reservoir, we applied the TZA assay to cryopreserved PBMCs obtained from HIV-1C-infected adolescents and successfully quantified the levels of inducible replication-competent viruses. The purity of the isolated rCD4+ T cells was >98% (Supplementary Fig. S2). After 6 days of activation in cell culture, the mean fold change in rCD4+ T cell count was 1.5 ± 10.8, which is consistent with previous studies 14 (Table 2).
Detection of Inducible Replication Competent Proviruses
IUPM, infectious units per million cells.
Inducible replication-competent virus detected in HIV-1C-infected adolescents
Despite the study participants having been on ART for 15 years, inducible replication-competent viruses were detected in all 15 subjects using the TZA assay. The median IUPM was 7.4 (IQR = 6.7–19.2) (Table 2). Further analysis showed no significant differences in the number of inducible replication-competent proviruses between the suppressed and unsuppressed individuals (Fig. 1A). In addition, no statistical difference in the levels of inducible replication reservoir was found between adolescents who initiated ART early (<12 months) and late (>12 months of age; Fig. 1B). In addition, we stratified the results according to when the adolescents had an HIV PCR-positive result (at 0, 1, and 2 months after birth). We did not find any statistically significant differences in the levels of inducible replication-competent viruses between the three groups (data not shown).

Small reservoir size observed in adolescents on long-term ART
The median total HIV-1 DNA copies per million cells were 83.3 (IQR = 18.2–172.3) for whole PBMCs, 53.5 (IQR = 15.9–116.9) for rCD4+ T cells before activation, and 129.1 (IQR = 18.9–212.3) for rCD4+ T cells after activation (Table 3). We found no statistically significant difference in the levels of total HIV-1 DNA copies per million cells across the three compartments (Fig. 2A). We observed a strong negative correlation between the size of the reservoir determined by calculating the fPVE and total HIV-1 DNA copies per million cells (p = .007, r = −0.81 Spearman rank test) (Fig. 2B). Similar to the number of inducible replication-competent proviruses analysis, we observed no significant differences in the number of total HIV-1 DNA copies between virally suppressed and unsuppressed participants (Fig. 3A). We observed no significant correlation between age at ART initiation and the size of the reservoir (IUPMs), or total HIV DNA copies per million cells (Fig. 3 B, C). However, we observed a trend indicating that adolescents who initiated ART later had higher levels of total HIV-1 DNA copies per million cells (p = .066, r = 0.506; Spearman test). No significant difference in the size of the reservoir was found between adolescents who were breast-fed as infants and those who received formula (data not shown).
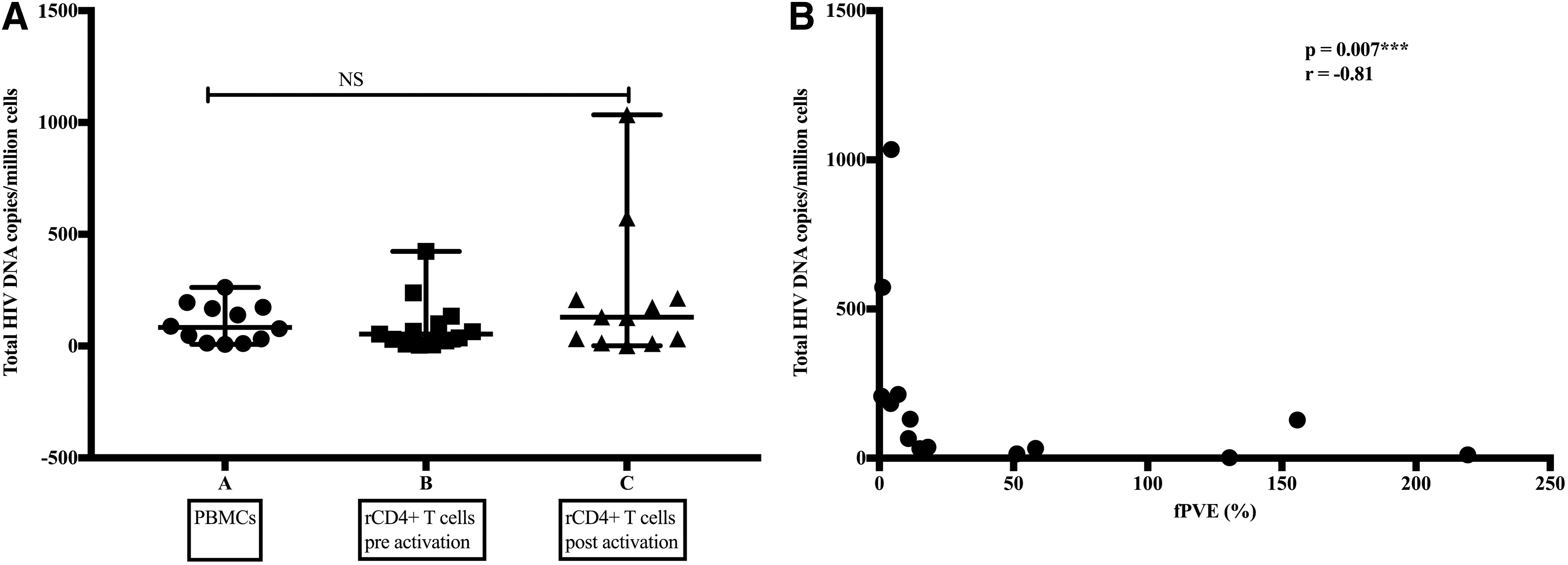
The relationship between total HIV DNA copy number and the size of the reservoir.
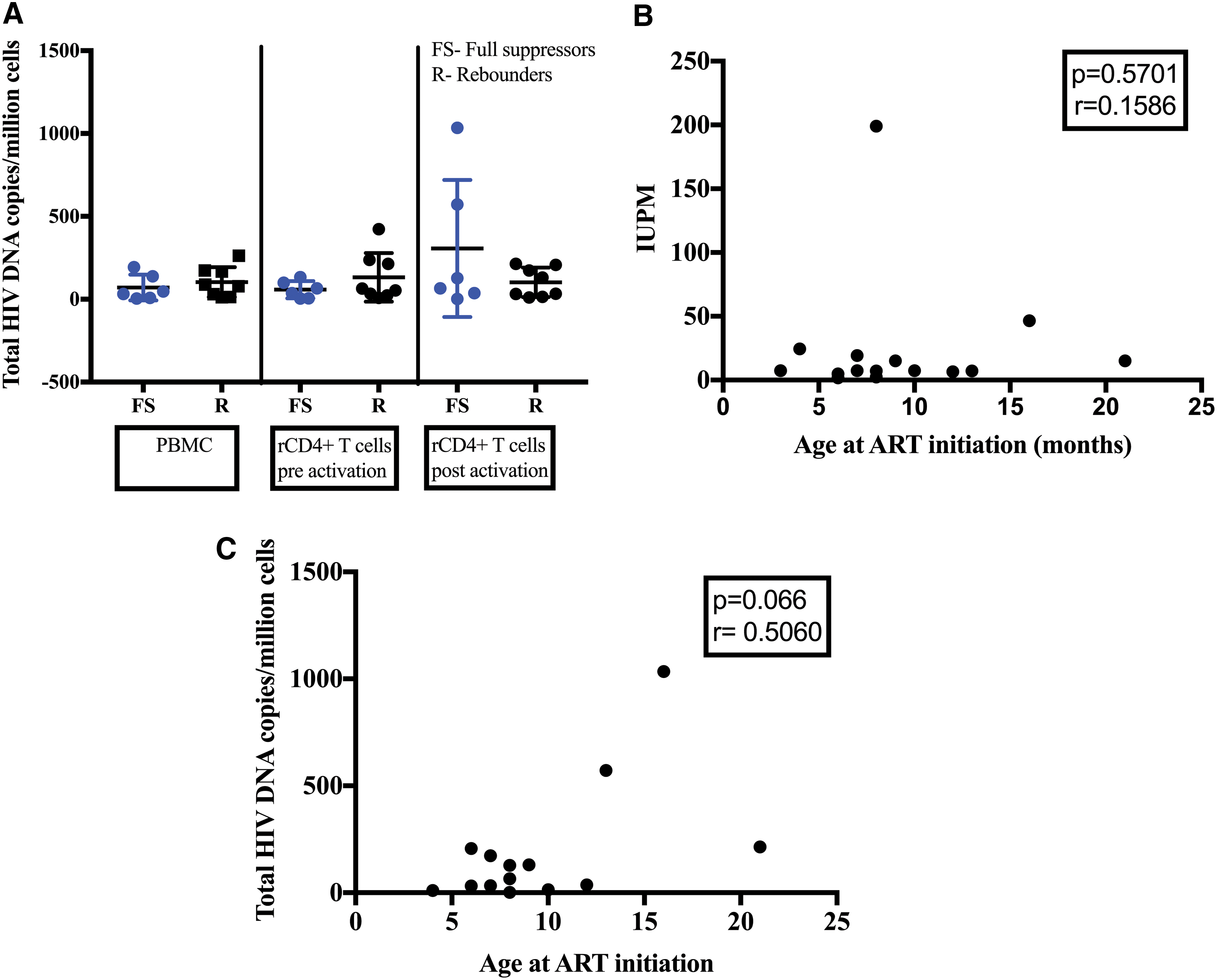
Small reservoir size among adolescents.
Quantification of Total HIV DNA by Droplet Digital PCR in the Three Compartments
PBMCs, peripheral blood mononuclear cells.
Discussion
This is the first study to assess the impact of long-term ART on the levels of inducible replication-competent provirus among adolescents perinatally infected with HIV-1C. A reporter cell-based assay termed TZA was used to assess and quantify inducible replication-competent proviruses in HIV-1C-infected adolescents from Botswana on long-term ART. To the best of our knowledge, this is the first study to use the TZA to quantify inducible replication-competent proviruses in a cohort of adolescents infected with HIV-1C. We detected inducible replication-competent reservoir in participants having been on ART for >13 years, which suggests that long-term ART and viral suppression did not eliminate the reservoir. Only 1 of the 15 participants had a viral load >2,000 HIV copies/mL at the time of enrollment in the study. Because the reservoir is established very early in HIV-1 infection, the observed increase in viral load might not correlate with the levels of the reservoir. The levels of inducible replication-competent proviruses were not affected by the infants age at initiating ART. No difference in the reservoir size was found between suppressed and unsuppressed participants. Our results are in line with previous animal model studies showing that ART initiation at 3 days post-simian immunodeficiency virus (SIV) infection did not prevent the establishment of a viral reservoir in tissues. 21
The TZA assay has been previously used to study the reservoir in HIV-1B infection. 14,16 We compared our results obtained in HIV-1C-infected adolescents with the cohort of 15 adults (mean age 55 years) on long-term ART (mean duration of 13.2 years) infected with HIV-1B and enrolled at a Pittsburgh clinical site in the Multicenter AIDS Cohort Study, 14 and found no significant difference in the median IUPM values between two studies (data not shown). We found no evidence that levels of inducible replication-competent proviruses differ between HIV-1 subtypes. It is important to note that TZA detects proviruses using a single round of infection, which is a known limitation of the assay. Therefore, it is unclear whether proviruses detected by this assay are capable of supporting multiple rounds of productive HIV-1 infection. Hence, it is possible that the TZA assay might be overestimating the replication-competent reservoir.
We used ddPCR to quantify total HIV-1 DNA copies in whole PBMCs, resting CD4+ T cells pre- and postactivation and found no differences in total HIV-1 DNA copies between the cell types. Total DNA copies detected in our cohort are higher than those observed in other adolescent cohorts. 22,23 It is important to note that children who initiated ART in the first 2 weeks of life have much lower HIV DNA copies. 22,23 In our cohort, adolescents initiated ART at a median of 8 months and there was a trend indicating that starting ART late might be associated with high total HIV-1 DNA copies. A recent study conducted in Botswana observed that treating infants shortly after birth can limit the size of the reservoir significantly, 2 indicating the clinical benefits of initiating ART immediately after birth. Initiating ART at 8 months may have been too late to maintain low reservoir size. Furthermore, we observed a strong negative correlation between the size of the reservoir determined by calculating the fPVE and total HIV-1 DNA copies per million cells. In some participants (n = 3), the fPVE percentage was >100%, which was puzzling. We speculate that it is possible that in those participants, there was excessive induction of replication-competent proviruses. However, the reason behind this observation remains unknown. Therefore, additional assessment of the reservoir is needed.
PCR-based assays detect ∼90% of defective proviruses and overestimate the size of the reservoir because they cannot distinguish between intact and defective provirus. Therefore, total HIV-1 DNA quantification may not provide a true representation of the replication-competent reservoir size. 23 It might be beneficial to use PCR-based assays in combination with other sensitive techniques to determine the true size of the reservoir. Single genome amplification might provide better information about whether the proviruses are defective or intact. We could not conduct this additional analysis owing to limited availability of T cells. A follow-up study should examine proviral sequences across the entire HIV-1 genome and assess integrity of the provirus. Current studies suggest that quantifying the proportion of intact proviruses might provide a more accurate assessment of the reservoir and the host factors that might influence reservoir size.
Conclusions
Our study demonstrated that long-term ART and sustained viral suppression did not eliminate reservoirs among perinatally infected adolescents with HIV-1C who initiated ART between 3 and 21 months of life. Because the reservoir is established very early in HIV-1 infection, it is possible that initiation of ART during the 1st year of life could not limit the reservoir. We found that TZA was a useful tool to quantify inducible replication-competent proviruses in HIV-1C infection. A follow-up study is needed to analyze the proviral landscape of the reservoir and determine whether quantifying the intact viruses may be better for monitoring the reservoir. Further research toward understanding the reservoir requires improvement of current assays and/or development of alternate high-throughput, easy-to-use assays able to assess, quantify, and monitor the proviral reservoir in HIV-1 infection.
Footnotes
Acknowledgments
The authors acknowledge G. Lee for technical advice on setting up the ddPCR assay. We would like to thank all the study participants. We would like to thank Lendsey Melton & Tumalano Sekoto for all their assistance.
Author Disclosure Statement
There are no conflicts of interest.
Funding Information
C.K.K. was supported by the Fogarty International Center (FIC) of the U.S. National Institutes of Health (D43 TW009610). C.K.K. and S.M. were supported by the Sub-Saharan African Network for TB/HIV Research Excellence (SANTHE), a DELTAS Africa Initiative (DEL-15-006). The DELTAS Africa Initiative is an independent funding scheme of the African Academy of Sciences (AAS)'s Alliance for Accelerating Excellence in Science in Africa (AESA) and is supported by the New Partnership for Africa's Development Planning and Coordinating Agency (NEPAD Agency) with funding from the Wellcome Trust (107752/Z/15/Z) and the U.K. government. The views expressed in this publication are those of the authors and not necessarily those of FIC, AAS, NEPAD Agency, Wellcome Trust or the U.K. government. ddPCR expression analysis was performed in the Boston Children's Hospital Intellectual and Developmental Disabilities Research Centre (BCH IDDRC) Molecular Genetic Core, supported by National Institutes of Health award grant no. U54 HDO9O255.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.