
Abstract
Microbial translocation has been suggested as a major driver of chronic immune activation HIV infection. Thus, we compared the extent of microbial translocation in patients with acute HIV infection and patients followed after CD4-guided structured treatment interruption (STI) by measuring different circulating markers: (1) lipopolysaccharide (LPS)-binding protein (LBP), (2) bacterial DNA, (3) soluble CD14 (sCD14), and (4) mitochondrial DNA (mtDNA). Bacterial DNA and sCD14 levels were similar in all groups. Patients in acute phase showed higher levels of LBP and mtDNA. In STI, we found a positive correlation between the percentage of CD8+ T cells and bacterial DNA levels. Considering all patients, LBP was positively correlated with the percentage and the absolute count of CD8+ T cells, and with mtDNA stressing the importance of mitochondrial products in sustaining chronic immune activation.
Introduction
Chronic HIV infection is characterized by a persistent immune activation, even after years of antiretroviral therapy (ART), leading to the onset of cardiovascular diseases, neurocognitive disorders, and senescence. One of the main drivers of this immune activation is the translocation of microbial products from the gastrointestinal tract to the systemic circulation, after the disruption of the epithelial barrier by the virus. 1 Microbial products contain pathogen-associated molecular patterns that trigger a proinflammatory cascade that ends in persistent immune activation. Furthermore, damage-associated molecular patterns such as circulating mitochondrial DNA (mtDNA), whose main source is cell death, can be released, enter into circulation, and activate innate immune cells. 2
Microbial translocation can be directly measured by the quantification of bacterial DNA, 3 whereas other markers, such as soluble CD14 (sCD14) and lipopolysaccharide (LPS)-binding protein (LBP), are indirect measures that reflect the innate immune response. To better understand the extent of microbial translocation of various circulating markers in different disease stages, we quantified bacterial DNA, LBP, sCD14, and mtDNA in plasma from groups of patients with acute HIV infection, patients with CD4-guided structured treatment interruption (STI; at various time intervals from therapy interruption to 12 months after therapy resumption) and healthy subjects. Correlations between bacterial DNA concentration with viro-immunological parameters were also performed.
Materials and Methods
We analyzed cryopreserved plasma samples from 19 patients with HIV infection enrolled into previous studies. 4,5 Eleven patients were enrolled during acute HIV infection within 2 months after infection and had a CD4+ T cell count of 846 ± 368 cells/μL (mean ± standard deviation) and a viral load (VL) of 1.1 ± 1.3 million copies/mL. Patients with acute primary infection were naive for therapy and had hepatitis C virus (HCV) coinfections: five hepatitis B virus (HBV) coinfections and four cases of syphilis. Eight patients were scheduled for a CD4-guided STI [samples had been collected at the time of therapy interruption (T0), 2 months after interruption (T1) and 1 year after therapy resumption (T2)]. Criteria for restarting treatment were a CD4(+) T lymphocyte count <350 cells/μL on two separate occasions, a clinical manifestation of AIDS, and the patient's desire to resume therapy. The three-drug combination ART was interrupted at a mean of 77.4 ± 22.3 months with a CD4 count of 1,124 ± 473 cells/μL, and undetectable viremia. All patients had HCV and HBV coinfections, three had syphilis. The same therapy was prescribed for six of eight patients at the moment of resumption. Furthermore, we included cryopreserved plasma samples from 10 healthy, concurrent, and age-matched subjects. The study was approved by the Area Vasta Emilia Nord Ethical Committee (Protocol no. 0006721/19). Circulating bacterial DNA was analyzed by the QX200 droplet digital polymerase chain reaction (ddPCR) system (Bio-Rad, Hercules, CA). DNA was extracted from plasma by using QIAmp DNA mini kit (Qiagen, Alameda, CA) and, as control, from water samples to avoid false results due to bacterial contamination. For amplification, we used the previously described 16S_F (5′-GCAGGCCTAACACATGCAAGTC-3′) and 16S_R (5′-CTGCTGCCTCCCGTAGGAGT-3′) primers, optimized for the ddPCR primers, optimized for ddPCR. 6 For each ddPCR reaction, we performed eight negative controls and a positive control of DNA extracted from Escherichia coli, in duplicate. Bacterial DNA copies were expressed per milliliter of plasma. We quantified circulating mtDNA by quantitative real-time PCR as previously described. 7 SCD14 and LBP plasma levels were detected by Human sCD14 Quantikine ELISA Kit (R&D Systems, Minneapolis) and Human LBP ELISA Kit (Abcam, Cambridge, United Kingdom), respectively. Quantitative variables between groups were analyzed by Mann–Whitney test for unpaired samples and Wilcoxon test for paired samples. The linear regression analysis was used to verify the correlations between analyzed parameters. Statistical analyses were performed using Prism 6.0 (Graphpad Software, Inc., La Jolla, CA).
Results
The 16S rDNA and sCD14 plasma levels were similar among groups and remained similar among the three time points for the CD4-guided STI; data not shown). Conversely, LBP levels were higher in patients with acute infection than in both healthy participants (p = .008; Mann–Whitney test) and STI patients at the time of treatment interruption (p = .015). LBP levels were also higher in STI patients at T2 than STI at T0 (p = .016, Wilcoxon test), as shown in Figure 1A. As shown in Figure 1B, circulating mtDNA concentration was higher in patients with acute infection than in STI patients at all time intervals (p = .020, p = .042 and p = .007, respectively). A trend also suggested that mtDNA concentrations were lower in healthy subjects than in patients with acute infection (not statistically significant).
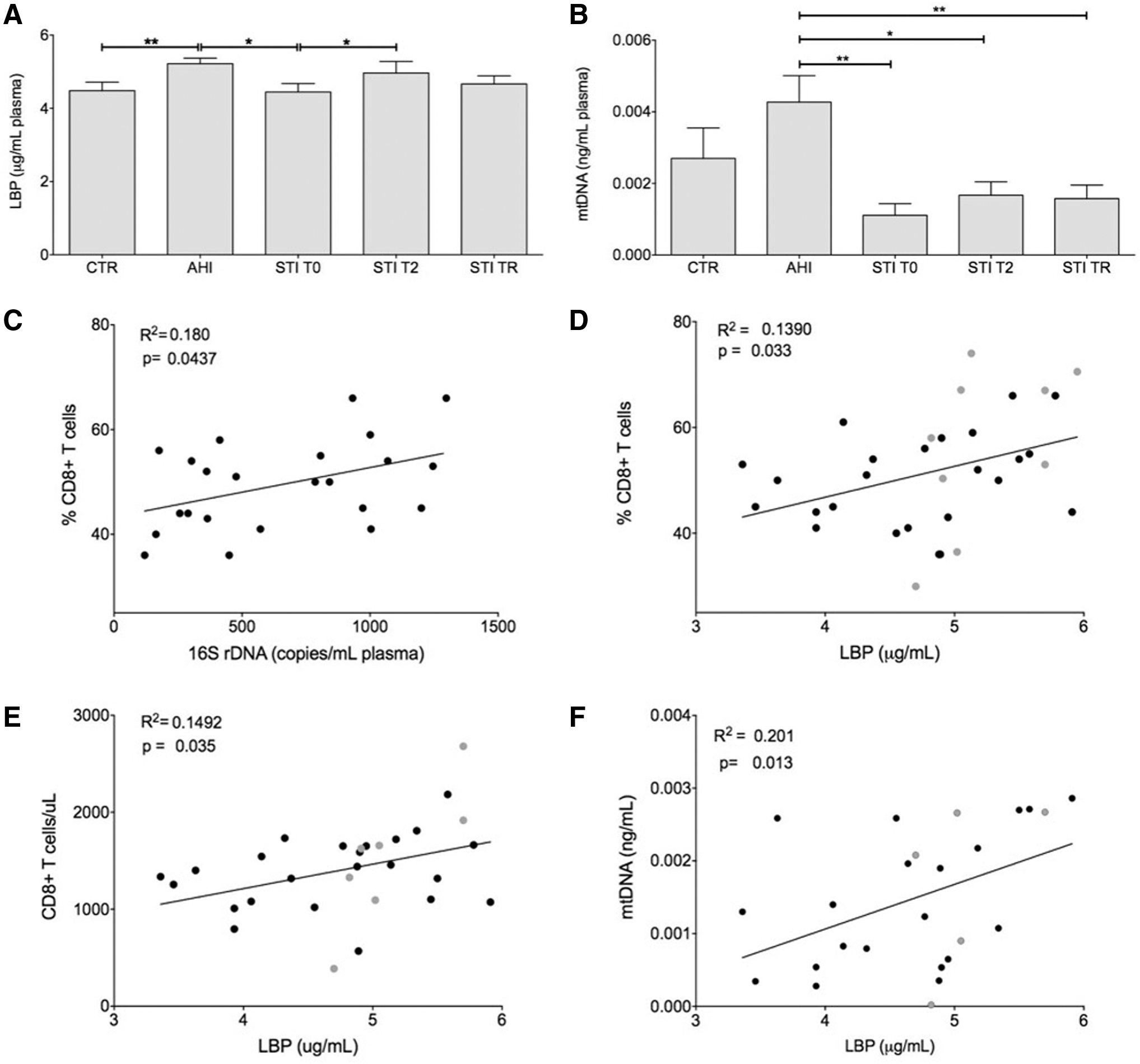
Quantification of soluble, circulating factors, and correlation between immunological and soluble factors. Column graphs representing mean and standard error of mean of LBP
Then, we analyzed the possible correlation between bacterial DNA concentration with viro-immunological parameters, including lymphocytes, CD8+ and CD4+ T cell count and percentage, CD4/CD8 ratio, and VL for HIV+ patients [acute HIV and STI patients (at T0, T1, and T2)]. The percentage of CD8+ T cells was positively correlated with bacterial DNA amount in STI patients (R 2 = 0.180, p = .044, Fig. 1C). When all HIV+ patients were considered, LBP plasma levels were positively correlated with the T cell count percentage of CD8+ T cells (R 2 = 0.139, p = .033, Fig. 1D and R 2 = 0.149, p = .035, Fig. 1E, respectively). Finally, a positive correlation was found between circulating mtDNA and LBP levels in all HIV+ patients (R 2 = 0.201, p = .013, Fig. 1F).
Discussion
Circulating markers that enable the measurement of the extent of microbial translocation are useful to better understand and monitor alterations induced by mucosal damage and their hypothetic recovery in different disease stages. Therefore, for certain approaches, the use of ddPCR, instead of real-time PCR, ensures an absolute quantification of nucleic acid with more sensitivity and accuracy, with lower background signal and without the need of a standard curve, the reason for which ddPCR has started to be widely adopted in the field of HIV infection. 8,9 In our study, bacterial DNA levels evaluated by ddPCR did not change between patients and healthy participants, or among patients in different stages of the disease, despite the presence of HIV/HBV (or HCV) coinfection, which is known to increase microbial translocation. Positive correlations with the percentage of CD8+ T cells in STI patients were observed. There has been conflicting evidence with regard to positive correlations between bacterial DNA and activated CD8+ T cells in previous studies. 3,10 Conversely, LBP directly correlates with CD8+ T cell count, CD8+ T cell percentage, and mtDNA.
Conclusions
Despite a limited sample size, our data enforce the concept that microbial translocation is a complex process and the bacterial load alone is not sufficient to differentiate between either acute or chronic disease stages or to explain persistent immune activation. The immunological response is likely to also depend upon other crucial factors that are able to trigger and drive inflammation, such as mtDNA.
Footnotes
Authors Contributions
M.N. and A.C. contributed to the conception and the design of the study. M.N., A.C., J.C., and M.P. contributed to the revision and the final approval of the article. M.N., J.C., and S.P. drafted the study. D.L.T. and S.D.B. contributed to the statistical analysis. M.N., C.M., and A.V.M. contributed to the interpretation of data. L.G. and A.D.G. provided the processing and storage of biological samples. S.P. and L.G. performed the molecular analysis. M.D. and C.M. enrolled the subjects for the study.
Acknowledgments
We thank the Italian National Agency for the Evaluation of the University System and Research (ANVUR) for the FFABR 2017 (funding for basic research activities) assigned to M.N. and the International Society for Advancement of Cytometry (ISAC) for the Marylou Ingram Scholar to S.D.B. We also acknowledge the valuable technical assistance of Dr. Lara Rossini (Bio-Rad specialist).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported FFABR 2017 (funding for basic research activities) to Milena Nasi assigned by National Agency for the Evaluation of the University System and Research (ANVUR).