
Abstract
It is now recognized that to fully understand the role of host genetic variation on susceptibility to HIV-1 infection, investigations must be extended to African populations. We sought to determine if genetic variation in IL10 are associated with HIV-1 infection in a West African cohort in Mali. HIV-infected and -uninfected individuals were genotyped for three common single nucleotide polymorphisms (SNPs) located at positions −592 (C/A), −819 (C/T), and −1082 (G/A) of the IL10 promoter. We found that the ATA haplotype, which has been previously associated with low IL-10 expression, was the most represented in the cohort. Although we observed a trend toward an increased frequency of ATA/ATA carriage in HIV-infected compared with -uninfected individuals, the difference was not statistically significant. Similarly, individual IL10 SNPs were not significantly enriched in the HIV-infected group, suggesting that IL10 genetic variants are not associated with HIV-1 in this West African cohort from Mali.
Introduction
Previous reports have revealed interindividual variability in susceptibility to HIV-1 infection and disease progression. A substantial part of this variability is thought to be determined by host genetic variation. However, only two genetic loci, notably the human leukocytes antigen class I (HLA) locus encoding for the amino acids in the HLA-A and HLA-B peptide binding groove and the chemokine (C-C motif) receptor gene cluster (mostly CCR5) have been clearly linked with HIV-1 disease susceptibility. 1 Indeed, most studies (candidate-gene analyses and genome-wide associations) have focused on Caucasian populations with few examining the impact of host genetic variation on HIV-1 infection in African populations. Given that the majority of new HIV infections occur in sub-Saharan Africa 2 and the recent finding of ∼3 million new genetic variants in Africans, which are not carried by individuals of non-African ancestry, 3 understanding the genetic basis of HIV-1 infection and disease outcomes in African populations should be a research priority. In this article, we sought to determine the influence of genetic variation in IL10 gene on susceptibility to HIV-1 infection in an ethnically diverse sub-Saharan West African population, specifically, in individuals living in Mali. We targeted this important immune-regulatory cytokine because of its potential role in regulating immune responses to HIV-1 and also the presence of genetic variants in IL10 gene that have been reported to influence its expression. 4,5 We concentrated our analyses on the three common proximal promoter single nucleotide polymorphisms (SNPs), namely −592 (C/A; rs1800872), −819 (C/T; rs1800871), and −1082 (G/A; rs1800896). The SNPs are known to be in tight linkage disequilibrium and form three classical haplotypes, namely ATA, ACC, and GCC haplotypes that have been associated with low, medium, and high IL-10 expression, respectively. 6 Remarkably, IL10 promoter SNPs and their haplotypes have been previously associated with differential susceptibility to HIV-1 infection in Caucasian and, also in South African populations. 7,8
Materials and Methods
We used peripheral blood mononuclear cells (PBMCs) of 159 study participants that were stored in a repository of samples in the laboratory of the University Clinical Research Center (UCRC) under an Institutional Review Board (IRB)-approved repository protocol. The UCRC repository consists of samples collected from previous prospective cohort studies aimed at understanding the pathogenesis of HIV-1 infection in Mali. Specifically, the parent studies were designed to assess longitudinal changes in immune responses during HIV-1 disease progression and the impact of opportunistic infections such as tuberculosis on key immunological markers. For the purposes of this study, the PBMCs were selected from the repository according to the following inclusion criteria: (1) HIV-1 (not HIV-2) positivity for the HIV-infected group, (2) HIV-1 and HIV-2 negativity for the HIV-uninfected group, and (3) availability of at least one vial of PBMCs stored in liquid nitrogen. The serology was performed using an ELISA assay (GenScreen™ ULTRA HIV Ag-Ab; Biorad), followed with western blot analyses (New Lav Blot I and II; Biorad). Of the 159 samples tested, 45% (71) were HIV-infected and 55% (88) were HIV-uninfected study participants. Genomic DNA was isolated from the PBMCs using DNeasy Blood & Tissue Kit from Qiagen according to the manufacturer's instructions. The extracted DNA was quantified and used as the template for genotyping by polymerase chain reaction (PCR) using Taqman SNP genotyping assay from Life Technologies (USA). Primers targeting IL10 proximal promoter SNPs were used for the PCR as follows: −592C/A (rs1800872; C_1747363_10), −819C/T (rs1800871; C_1747362_10), and −1082G/A (rs1800896; C_1747360_10). Homozygote and heterozygote genotypes were distinguished, based on the presence of one or both alleles at the same position, using the Taqman SNP genotyping software. Data were then exported for further analysis using GraphPad Prism (version 7). All statistical tests were performed using a Fisher's exact test by utilizing the R statistical software. A two-tailed p-value was calculated for each comparison. Any p-value <.05 was considered to be statistically significant.
Results
First, we sought to determine the frequency of the three IL10 SNPs in both of the HIV-infected and -uninfected groups. At position −592, C/C, C/A, and A/A were all detected with the heterozygous C/A predominating (Fig. 1A). Similarly, at position −819, C/T heterozygotes were the most represented among the genotypes (C/C, C/T, and T/T) (Fig. 1B). In addition, the G/A and A/A genotypes were more frequent than the G/G genotype at position −1082 (Fig. 1C). Despite higher frequencies of the A/A and T/T in individuals infected with HIV-1 at position −592 and −819, respectively, the p-values indicate lack of statistically significant differences between the two groups (A/A: odds ratio [OR] = 1.52; 95% confidence interval [CI] of OR = 0.69–3.39; p = .27 and T/T: OR = 1.63; 95% CI of OR = 0.73–3.68; p = .26). Our data suggest that the individual SNP genotypes of the three common IL10 proximal promoter SNPs are not associated with susceptibility to HIV-1 infection in the study population.
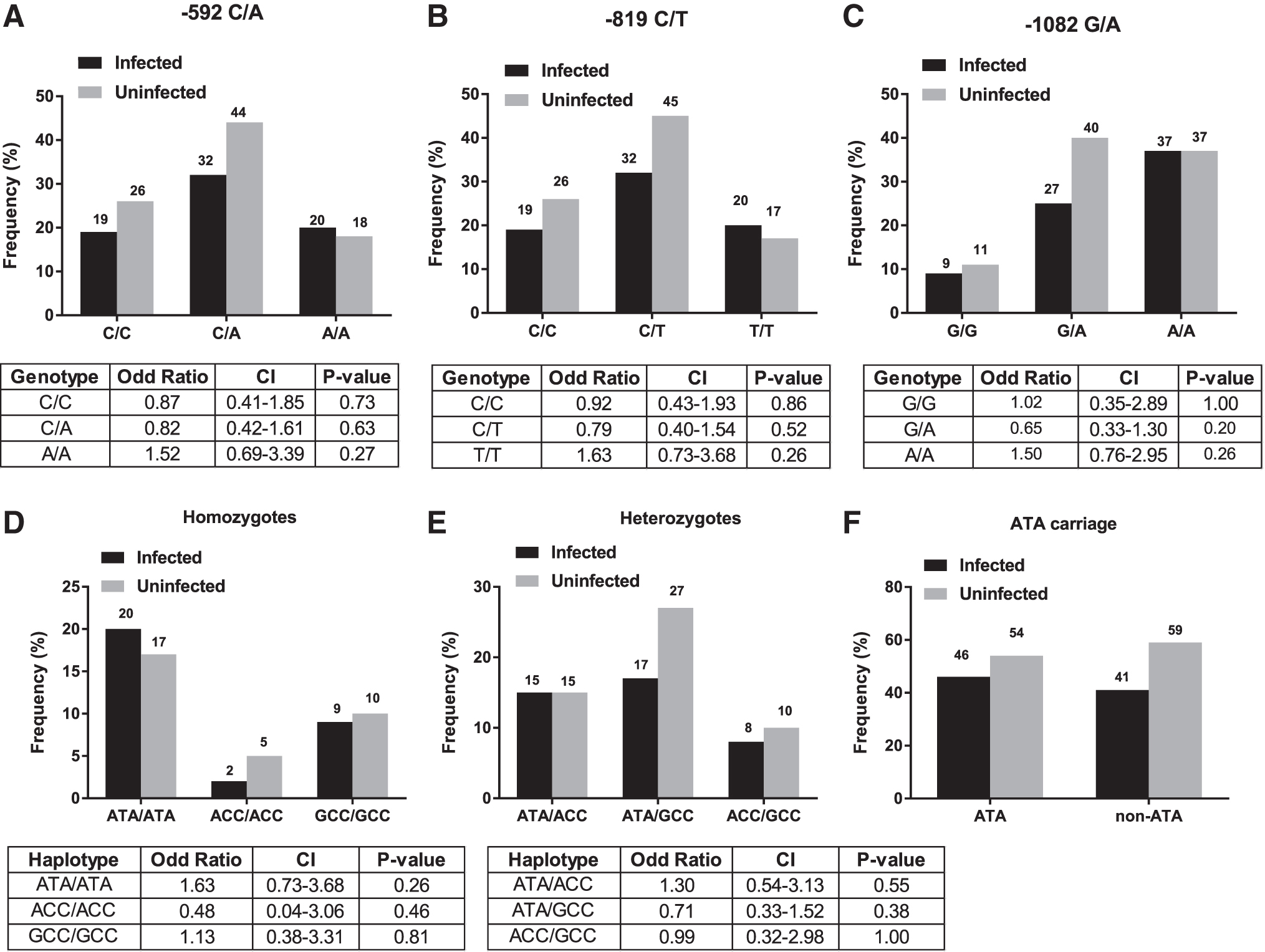
Frequency distribution of genetic variation in IL10 in our study population. Bar graphs represent the proportion of individual IL10 SNPs
Then, we aimed to determine frequencies of IL10 haplotypes, which are the different combinations of the three SNPs, and their potential association with the risk of HIV-1 infection. Of the 159 samples analyzed, we found 10 haplotypes, namely ATA/ATA, ACC/ACC, GCC/GCC, ATA/ACC, ATA/GCC, ACC/GCC, ATA/ACA, ATA/ATC, GCC/GGC, and GTA/GCC. When we categorized the haplotypes based on their homozygosity or heterozygosity, we found three homozygous haplotypes, specifically ATA/ATA, ACC/ACC, and GCC/GCC (Fig. 1D) and seven heterozygous IL10 haplotypes, namely ATA/ACC, ATA/GCC, ACC/GCC, ATA/ACA, ATA/ATC, GCC/GGC, and GTA/GCC. Among the seven heterozygous haplotypes, four haplotypes (ATA/ACA, ATA/ATC, GCC/GGC, and GTA/GCC) were found at very low frequency (≤1%), and thus are not displayed on Figure 1E. The remaining three heterozygous haplotypes (ATA/ACC, ATA/GCC, and ACC/GCC) are shown on Figure 1E. With respect to HIV-1 status, we found that the ATA/ATA homozygous haplotype was represented at a higher frequency in the HIV-infected group (20%) when compared with the HIV-uninfected group (17%), suggesting that the ATA/ATA haplotype may increase susceptibility to HIV-1 (Fig. 1D). However, the odds of being an ATA/ATA carrier in HIV-infected group was not significantly different from that of HIV-uninfected group (OR = 1.63; 95% CI of OR = 0.73–3.68; p = .26). Unlike the ATA/ATA, the ATA/GCC heterozygous haplotype was more frequent in the HIV-uninfected group (27%) relative to the HIV-infected group (17%), suggesting that this haplotype may confer resistance to HIV-1 infection (Fig. 1E). However, we found no association between the ATA/GCC and a reduced risk of HIV-1 infection (OR = 0.71; 95% CI = 0.33–1.52; p value = .38). These results suggest that the IL10 haplotypes (homozygous and heterozygous) of the study participants are not associated with HIV-1 infection in Mali.
Given the lack of association between the IL10 SNPs and their haplotypes and the risk of HIV-1 infection, we stratified our cohort, based on ATA carriage, to assess the influence of having a high proportion of ATA carriers on our analysis. We found that 71% of our study participants were ATA carriers versus 29% of non-ATA carriers (data not shown). The non-ATA carriers consisted of ACC and GCC carriers at a frequency of 16% and 13%, respectively. When we focused on HIV infection, the proportion of HIV-infected individuals was notably similar between ATA carriers and non-ATA carriers (46% versus 41%) (Fig. 1F). Thus, our data suggest that the enrichment of ATA-carriage in our study population is unlikely to explain the absence of association found between IL10 haplotypes and HIV-1 status in our study population.
Discussion
In this pilot study, we found no association between IL10 promoter SNPs or haplotypes and susceptibility to HIV-1 in a West African population from Mali. Our findings suggest a differential effect of IL10 SNPs on HIV-1 infection susceptibility, based on host ethnicity. This is because our results diverge from a prior study in Caucasians by Shin et al. in determining the impact of IL10 SNPs on the risk of HIV-1 infection. 8 Our data, however, are well aligned with a study, conducted in African Americans 9 and with a recent meta-analysis of IL10 SNPs from 42 eligible studies, consisting of 15,727 HIV-infected and -uninfected participants from diverse ethnicities, including Africans. 10 Nevertheless, our results are also not consistent with a study conducted in South Africa, which reported a higher risk of acquiring HIV-1 in −592A carriers. 7 A possible explanation of the difference between our study and the South African study could be due to genetic differences between West and South Africans or the inherent genetic diversity of HIV-1 strains that circulate within these two regions. This hypothesis is supported by the findings of the African Genome Variation Project (AGVP), which have revealed key regional differences across Africa. 11 For example, AGVP has found a substantial heterogeneity within African populations in four loci associated with susceptibility to malaria, including the IL10 locus. Thus, it is likely that similar mechanisms could be driving such differential genetic susceptibility between geographically distinct African populations in the setting of HIV infection.
Conversely, genetic diversity of the virus could be also contributing to differential disease susceptibility by region or ethnicity because different HIV-1 subtypes have been shown to express differential levels of IL-10. Notably, isolates of HIV-1 subtype C, the most common HIV-1 subtype in Southern Africa, have a common mutation (cysteine to serine) that occurs in HIV transactivating proteins (Tat) resulting in a lower level of IL-10 expression in monocytes when compared with HIV-1 subtype B predominantly found in Europe and North America. 12 Thus, similar mechanisms of differential IL-10 expression could be in place between HIV-1 subtype C and the recombinant HIV-1 strains CRF02_AG that circulate in West Africa, including Mali. 13
It is worth noting that our findings, regarding the predominance of ATA carriage in Mali, are consistent with previous studies about the distribution of IL10 haplotypes across different ethnic groups. Similar to our study, Hoffman et al. found that ATA-inclusive haplotypes were significantly more common in Africans than in Caucasians. 14 These findings were later confirmed by Oleksyk et al. who found 44% of all ATA-inclusive haplotypes in African Americans and only 21% in Caucasians. 9 These data imply that frequencies of IL10 SNPs and haplotypes are not uniform across ethnicities, which could have implications for clinical evaluation of therapeutics aimed at targeting IL-10-dependent pathways to limit HIV-associated chronic immune activation.
Limitations of our study include the small sample size and lack of clinical information regarding stage of HIV-1 infection. Specifically, we cannot exclude the possibility that we underestimated the effect size or analyzed a heterogeneous population consisting of both acute and chronic HIV infection. Since, it has been proposed that the role of IL-10 during HIV could be infection-phase dependent, 4,7,15 studying such a heterogeneous population may have diluted the effect size of the IL10 SNPs. Also, it might have been fruitful to follow high-risk HIV-uninfected subjects of known IL10 genotypes across time and regularly assess the proportion of subjects who became infected during the follow-up period, but such a study was beyond the scope of the project. Therefore, future larger-scale studies with long-term follow-up of well-characterized study participants are warranted to confirm these results and to clearly define the role of genetic variation in IL10 gene on HIV-1 susceptibility and disease outcomes.
Footnotes
Acknowledgments
The authors thank the clinical and laboratory personnel of the University Clinical Research Center (UCRC) at the University of Sciences, Techniques, and Technologies of Bamako (USTTB) in Mali. We are also thankful to the members of the Bishai and Bream Laboratories at the Johns Hopkins School of Medicine (Center for Tuberculosis Research) and the W. Henry Feinstone Department of Molecular Microbiology and Immunology at the Johns Hopkins Bloomberg School of Public Health.
Authors' Contributions
D.D., R.L.M., and S.D. designed the study, analyzed and interpreted the results, and drafted the article; B.B., D.G., and S.D. enrolled the participants of the study and reviewed the article; B.K., M.W., N.C., Y.S.S., and M.B. developed the repository, processed the samples, collected the data, and reviewed the article; M.D. and M.U. analyzed the data and reviewed the article; J.H.B. and W.R.B. provided new analytical tools and reviewed the article; J.L.H., S.L.K., C.A., M.D., and S.D. critically edited and reviewed the article.
Disclaimer
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Research reported in this publication was supported by Fogarty International Center of the National Institutes of Health, under award D43TW010350 (MPI: R.L.M and S.D.). D.D. is also independently supported by Fogarty International Center and the Office of the Director of the National Institutes of Health through the Office of Research on Women's Health, Career Development Award K43TW011426 (PI: D.D.).