
Abstract
To describe distributions of immune markers in children and young adults by sex and HIV status, and within groups, investigate associations of immune markers with bone density across Tanner stage. Using data and samples from 353 participants in a cross-sectional study in youth with perinatally acquired HIV (PHIV) and matched HIV-negative controls, distributions of inflammation and activation immune markers were described by sex and HIV status. Correlations and structural equation models (SEM) were used to explore marginal and multivariable associations of the immune markers with bone density and to assess whether patterns of association varied by sex and HIV status. Immune marker distributions did not differ by sex, but there were some differences by HIV status. Correlation patterns among bone, body composition, and immune markers were similar across the sex and HIV status groups. Conclusions from SEMs were limited by small sample sizes, but there was some indication that patterns of association between bone density and certain immune markers differed in male PHIV with more advanced Tanner stage compared to the other three groups. In conclusion, distributions of bone density, body composition, and immune markers may vary by sex and HIV status, although associations among these outcomes within sex and HIV status groups appear similar. Bone density of male PHIV appears to be more negatively affected than females, regardless of female HIV status. Larger longitudinal studies across Tanner stages are needed to further explore potential biological relationships between immune markers and bone density in youth living with HIV.
Introduction
Bone loss is a major comorbidity in individuals living with HIV on antiretroviral therapy (ART), 1 yet, basic pathogenic mechanisms that lead to bone loss remain elusive, and there are few published studies on immune markers and bone in children or adults living with HIV. 2 –5 Understanding these mechanisms is critical in children and adolescents, as bone mass accrual during this period is essential to bone health later in life. 6,7 Previous publications, including an analysis of the cohort 8 described in this article, showed that males with perinatally acquired HIV (PHIV) had lower bone mineral density (BMD) than females with PHIV and HIV-negative males, and differences increased with advancing Tanner stage.
Seminal studies on bone metabolism have demonstrated reciprocal links and cross-talk between the skeletal system, adipose tissue, endocrine organs, and the immune system. 9 –12 HIV directly perturbs the immune system, and HIV and ART affect body fat distribution and lipid metabolism, leading to chronic inflammation and possible effects on subsequent bone health. We hypothesized that beneficial effects of estrogen on factors regulating bone health, including immune activation and inflammation markers, protect female PHIV from bone loss more than male PHIV.
Pediatric AIDS Clinical Trials Group (PACTG) Protocol P1045 was a cross-sectional study that collected bone density and body composition measures in PHIV and HIV-negative children, adolescents, and young adults (youth) (clinical trial registration number NCT00069004). Selected activation and inflammation markers were assayed from stored samples. In this article, we first compared immune marker distributions by sex and HIV status. We then used structural equation models (SEM), which allow exploration of causal relationships in complex networks, to investigate possible biological relationships between immune markers and bone outcomes (whole-body bone mineral content [BMC], whole-body BMD, and lumbar spine [LS] BMD) and to evaluate whether these relationships were modified by Tanner stage within groups defined by sex and HIV status.
Materials and Methods
PACTG P1045 study design
P1045 was a cross-sectional study of 396 children and adolescents enrolled at 37 PACTG sites in the United States. 13 Youth living with PHIV aged 7 to <25 years were enrolled into one of six strata defined by protease inhibitor use (yes/no) and Tanner stage (1, 2–3 and 4–5). A comparison group of youth without HIV (HIV-negative) with a similar distribution of sex and race/ethnicity within each Tanner stratum were also enrolled. The study was approved by the Institutional Review Board at each site and informed consent was obtained from each participant older than 18 years of age, and from the parent or guardian of younger participants.
Measurements
Tanner stage classification
Tanner stage, which represents stages of progression through puberty, was determined by inspection of breasts and pubic hair for females and genitalia and pubic hair for males. 13 Since growth velocity is greatest in Tanner Stage 3 and 4, Tanner stage for this analysis was grouped as 1–2, 3–4, and 5.
Bone and body composition
Dual-energy X-ray absorptiometry (DXA) scans were analyzed centrally at the Body Composition Analysis Center at Tufts University School of Medicine. In this analysis, bone density was represented using whole-body BMC, whole-body BMD, and LS BMD. Body composition was represented by total (legs plus arms) extremity fat (EXFT), total trunk fat (TRFT), and total (legs plus arms) lean extremity mass (EXLN).
Immune markers
Eleven activation and inflammation markers were measured in participants with sufficient stored plasma/serum (pg/mL). Inflammation markers included interferon gamma (IFN-γ), tumor necrosis factor alpha (TNF-α), and adiponectin. Activation markers included five interleukin markers (IL-6, IL-17, IL-7, IL-12p40, and IL-1β), two soluble clusters of differentiation markers (sCD40L and sCD14), and soluble vascular cellular adhesion molecule (sVCAM). sCD14 concentrations were measured by Quantikine ELISA (R&D Systems, MN). The other analytes were measured by MagPix Assays (Millipore, Germany). Adiponectin and sVCAM were run in a duplex assay, and all others were run together. Samples were run in triplicate, and coefficients of variation (CVs) were calculated; all analytes with a CV >10% were rerun with or without dilution as needed. Immune markers with values below the assay limits of detection (censored) were set to the lower limit of detection and measurements log10-transformed.
Statistical methods
Distributions of bone density (BMC, BMD, LS BMD), body composition (EXFT, TRFT, EXLN), and immune markers were summarized by sex and HIV status, and compared across groups in linear regression models with main effects for sex and HIV status and their interaction. If the significance level for the interaction was <0.05, the term was maintained in the model. Pairwise Pearson correlations were calculated between the bone, body composition, and immune markers.
Because SEMs assume multivariate normality and cannot accommodate censoring, 14 the three markers with more than 30% censoring (IL-1β, IL-6, and IL-12p40) were not included in the SEM analysis. Bone, body composition (log10-transformed), and immune marker measurements (log10-transformed) were put on a similar scale of age- and sex-adjusted Z-scores based on distributions in the HIV-negative males and females separately, as previously described. 13 To avoid undue influence of outliers, only participants with Z-scores in the range of −5 to 5 on all measurements were included.
The model included two unobserved latent variables: bone (BONE) measured by BMC, BMD, and LS BMD and body composition (BODY) measured by EXFT, TRFT, and EXLN. Given the complexity of potential biological relationships and to avoid model misspecification of pathways, no latent structure was assumed for the immune markers. Tanner stage and race/ethnicity were included as independent variables (bone is known to vary by these factors) 15 as well as an interaction term between Tanner stage and immune marker to allow associations of immune factors and BONE to vary across Tanner stage.
The model is illustrated in Figure 1. Given the limited sample size and large number of factors of interest, models were fit in each of the four sex/HIV status groups separately. A final parsimonious model for each group was chosen using stepwise Akaike Information Criterion. Goodness-of-fit was assessed using the Comparative Fit Index (CFI) and root mean squared error approximation. 14 Adequate fit is generally described as having CFI >0.9 and models with CFI >0.8 were considered acceptable. We report adjusted estimates and 95% confidence intervals of the relationship between each immune marker Z-score and the BONE latent variable by Tanner stage. Estimates by Tanner stage were calculated by combining immune marker main effects and interaction terms.
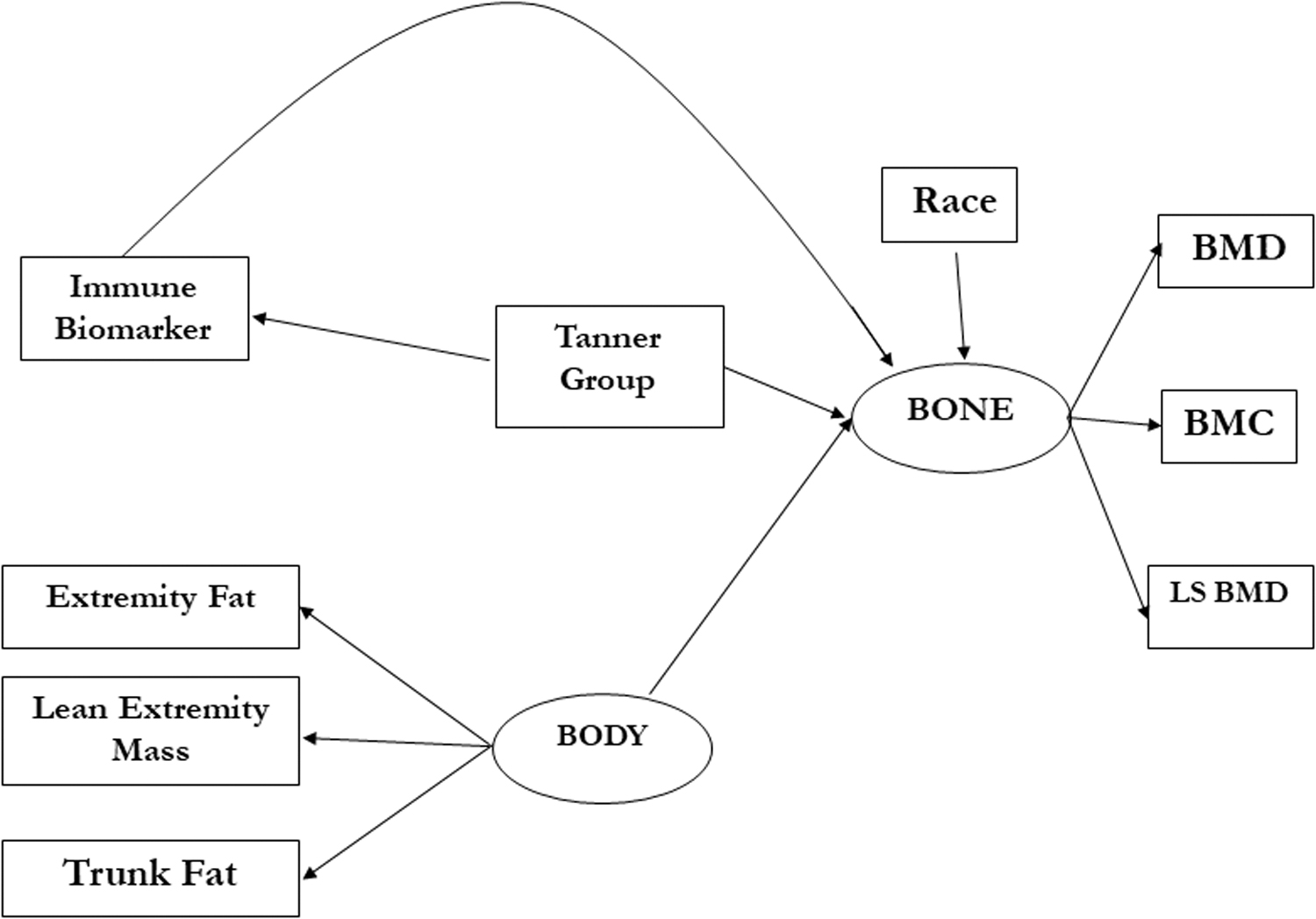
SEM model from which the best model was chosen for each sex/HIV status group. BMC, whole-body bone mineral content; BMD, whole-body bone mineral density; LS BMD, lumbar spine bone mineral density; SEM, structural equation models.
Results
Population characteristics
Of the 396 participants enrolled in P1045, 379 (96%) had DXA results, of whom 353 (93%) had sufficient samples to assay the 11 immune markers. Characteristics of these 353 participants by sex and HIV status are summarized in Table 1 (median immune marker levels by Tanner stage are listed in the Supplementary Table S1). Median age in the four groups ranged from 11.7 to 12.8 years. The majority of participants (53%) were Tanner stage 1–2.
Characteristics of Analysis Population
BMC, bone mineral content; BMD, bone mineral density; BMI, body mass index; EXFT, total extremity fat mass; EXLN, total extremity lean mass; IFN-γ, interferon gamma; IL, interleukin; LS BMD, lumbar spine bone mineral density; PHIV, perinatally acquired HIV; sCD, soluble cluster of differentiation; sVCAM, soluble vascular cellular adhesion molecule; TNF-α, tumor necrosis factor alpha; TRFT, trunk fat mass.
Linear regression models of bone, body composition, and immune markers included main effects for sex and HIV status but no interaction terms (p > .05 for all models, results not shown). Adjusted for HIV status, mean LS BMD, EXFT, and TRFT were higher and EXLN was lower in females compared to males (all p ≤ .007). Adjusted for sex, mean EXFT was lower in PHIV compared to HIV-negatives (p = .002). Boxplots of the 11 immune markers by sex and HIV status are shown in Figure 2. Based on the regression models, none of the immune markers differed by sex, and six differed by HIV status: mean TNF-α, sVCAM, and sCD14 were higher in PHIV, and IL-17, IL-12p40, and adiponectin were lower in PHIV (all p ≤ .05).

Boxplots of immune markers by sex and HIV status.
Correlations between bone, body, and immune measures
Pearson correlations within each sex and HIV status group are illustrated in Figure 3. Correlation patterns were similar in the four groups. As expected, the three bone outcomes were strongly positively correlated with each other (0.89 ≤ ρ ≤ 0.96). The bone measures were more positively correlated with lean extremity mass than with the two fat measures. The two fat measures were more strongly positively correlated with each other than with lean extremity mass. Bone outcomes were moderately negatively correlated with adiponectin in both PHIV and HIV-negative males and with IL-12p40 in HIV-negative males and females. Adiponectin, sVCAM, and sCD14 had low correlations (−0.02 ≤ ρ ≤ 0.38) with each other and the other immune markers. Other than sCD40L, the other six markers were fairly consistently positively correlated (0.20 ≤ ρ ≤ 0.78) with each other.
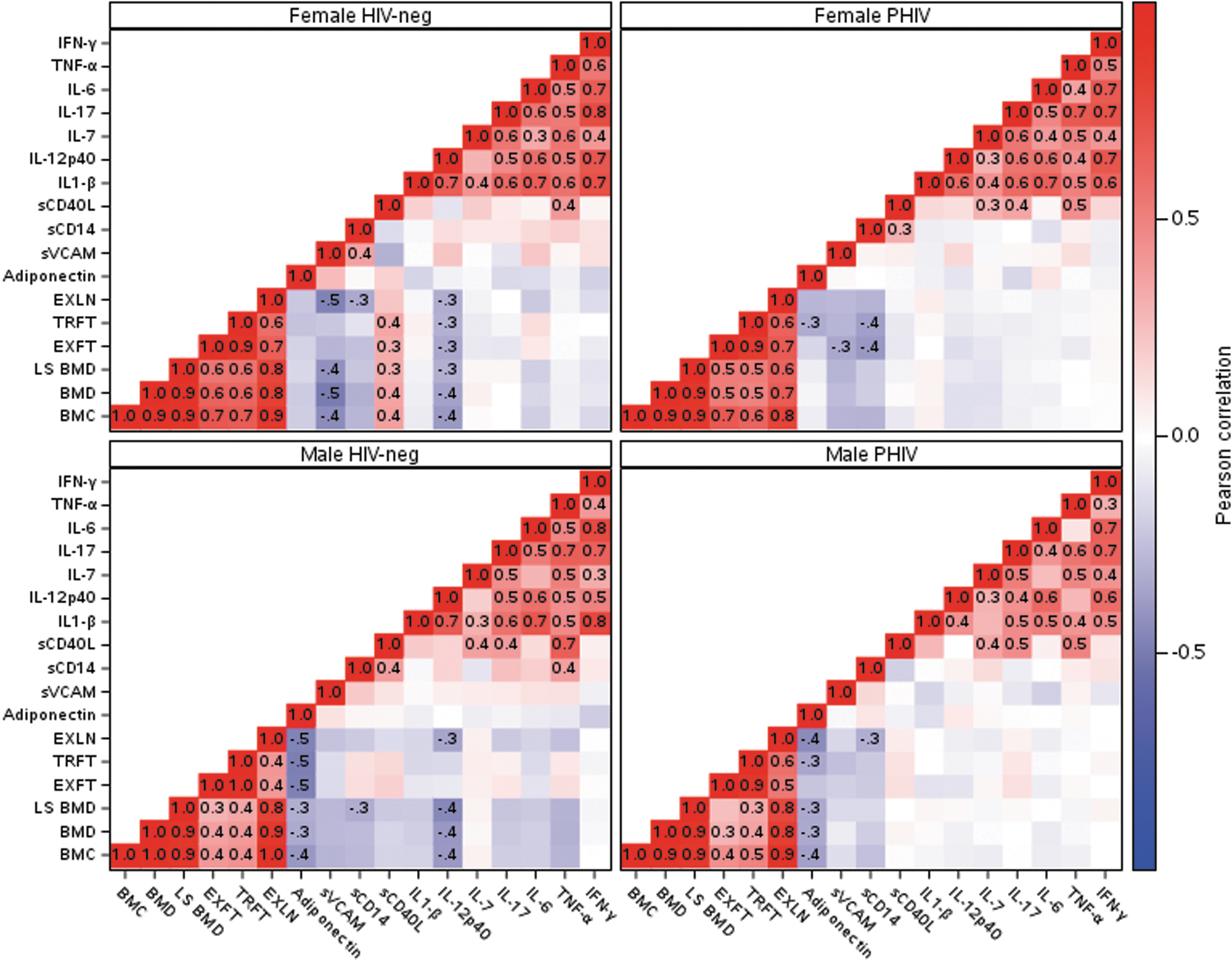
Pearson correlations (red: positive, blue: negative, degree of color saturation reflects magnitude) between standardized bone-profile variables (lower left), body composition measurements (middle), and immune markers (upper right). EXFT, total extremity fat mass; EXLN, total extremity lean mass; TRFT, trunk fat mass.
Structural equation models
The SEM analysis included 344 of the 353 participants with immune marker data (9 participants were excluded with outliers for at least one marker). Stepwise selection was used to determine if the proposed model in Figure 1 could be simplified. Race/ethnicity was not associated with BONE in any of the four groups and was dropped from the model.
Results from the final models for the association of BONE with each immune marker across Tanner stage are illustrated in Figure 4 (estimates are shown in Supplementary Table S2). Estimates are interpreted as unit increases in BONE Z-score for each unit increase in the immune marker Z-score, adjusted for the BODY latent variable. The final models had a good fit (CFI ranging from 0.82 to 0.96). As expected, the BODY latent variable was positively associated with BONE in all HIV/sex groups (associations not shown). No obvious or consistent patterns emerged across Tanner stage, sex, or HIV status. Male PHIV tended to have lower (and mostly negative) associations with bone in Tanner 3–5 compared with each of the other three groups for INF-γ, TNF-α, IL-17, and sCD14. In female PHIV, positive associations of bone with INF-γ, TNF-α, sCD40L, and adiponectin were observed in Tanner 5 and a negative association with IL-17 at Tanner 3–4.
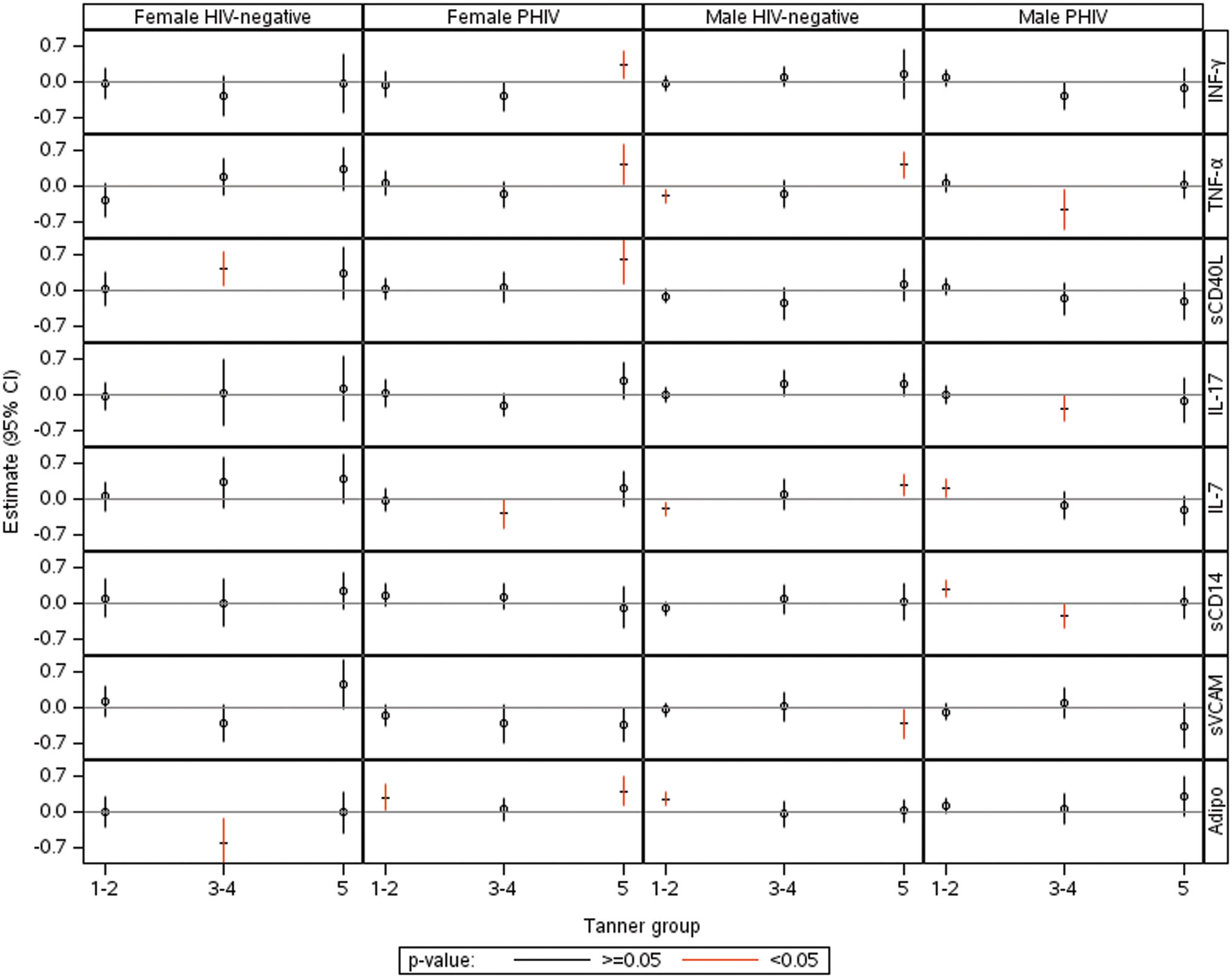
Effect estimates and 95% confidence intervals for the association of each individual marker on BONE modified by Tanner stage in each sex/HIV status group, holding the latent BODY variable constant.
Discussion
To try to better understand why male youth with PHIV and more advanced Tanner stage had more compromised bone density than females with PHIV and HIV-negative youth, we used stored samples to measure immune marker levels. Marker levels were summarized and compared by sex and HIV status, and their marginal and multivariable associations with bone density were explored.
In this cohort, PHIV had higher mean levels of TNF-α, sVCAM, and sCD14, and lower IL-17, IL-12p40, and adiponectin than in HIV-negatives. Bone outcomes were moderately negatively correlated with adiponectin in males, and with IL-12p40 in HIV-negatives. Research has shown that there is cross talk between fat and bone cells, such as through adiponectin, a fat-derived adipokine with anti-inflammatory effects whose production can be inhibited by TNF-α and IL-6. Adiponectin appears to have an independent role in bone development and metabolism in young children, 16 –18 and levels have been reported to be inversely correlated with BMD with variation by sex. 19,20 In one study of adults living with HIV, IL-7 concentrations did not differ in those with normal versus low BMD, 3 and in another cross-sectional analysis, Erlandson et al. found no association between TNF-α and bone. 4 Resino et al. found higher IL-7 concentrations in children living with HIV who had low CD4 counts compared to HIV-negative. 21 In studies of adults living with HIV, IL-7 levels are higher in adult women compared to men living with HIV 22 and higher in adults with increasing T cell depletion. 23 IL-17 concentrations are elevated in adults living with HIV, but relatively lower in those with higher BMI. 24 In a study of young PHIV in South Africa (all Tanner Stage 1 and 2), Shiau et al. found no association between sCD14 and whole-body BMC or bone turnover markers. 5 Little is known about the relationship between immune markers and bone outcomes among youth who did not acquire HIV perinatally. In one study that measured 19 immune markers in young adult males who acquired HIV through risk behaviors (not perinatally or through the blood products) and healthy controls, three markers (sCD27, sCD14, and sCD163) provided the best discrimination between the two groups. 25 Among youth who acquired HIV through risk behaviors, BMD was lower in those on ART compared to ART naive and HIV-uninfected youth. 26
The SEM models provided minimal insight into how associations of bone with the immune markers varied by sex, HIV status, and Tanner stage after controlling for body composition. These associations are complicated and the small sample sizes made effect estimates imprecise. One observation was that male PHIV tended to have more negative associations with bone in Tanner 3–5 groups for INF-γ, TNF-α, IL-17, and sCD14, not observed in the other Tanner groups. In male and female PHIV, estimates for IFN-γ, TNF-α, IL-17 (females only), IL-7, and adiponectin tended to decrease from Tanner 1–2 to 3–4 and increase to positive levels at Tanner 5. Bone velocity is highest during Tanner 3–4. 27 In healthy children, there is balance between bone resorption and bone formation during adolescence, favoring bone formation. 28 It is possible that elevated levels of cytokines have a more deleterious effect than other factors during rapid bone accrual in male PHIV. The positive associations at Tanner 5 in female PHIV suggest a decreased effect of proresorptive cytokines favoring greater bone formation. Bone resorption is decreased through activation of CD40 in B cells to produce osteoprotegerin (OPG), which blocks receptor activator of NF-jB ligand (RANKL) attachment, and through CD40L to increase the number of mesenchymal stem cells from which osteoblastic cells originate. The RANKL/OPG ratio is elevated in other inflammatory diseases, such as in juvenile idiopathic arthritis, 29 and IL-6 levels are elevated in children with inflammatory bowel disease. 30 IFN-γ can have inhibitory effects on osteoclastogenesis, leading to less bone resorption. 31 The reason for a positive association of TNF-α with bone in female PHIV at Tanner 5 is not clear.
This study contributes to our knowledge about immune marker distributions across Tanner stage by sex and HIV status as there are no reference standards for immune markers in healthy children and adolescents. We attempted to address gaps in the literature on underlying immune mechanisms of inflammation on bone, but two factors hampered this goal. First, small sample sizes (made smaller by deletion of nine participants with outlying immune marker values) limited the multivariable analyses, and we were unable to control for all factors potentially associated with immune levels and bone density, such as antiretroviral treatment history and HIV-1 RNA levels. In addition, apart from consequences from HIV, there are numerous factors that can adversely impact bone accrual, including vitamin D insufficiency, 32 delayed puberty, 33 body composition abnormalities, 34 and concomitant medications (corticosteroids, medroxyprogesterone). 35,36 Second, immune markers and DXA-derived bone outcomes were measured at a single time point, limiting our ability to determine temporal relationships. A balance between the effects of osteocytes and osteoblasts on bone formation and bone resorption, respectively, serves to maintain bone architecture in the adult skeleton. In contrast, in children, the balance of bone formation and bone resorption at different periods of development serves to contribute to bone growth and laying down new bone until peak bone mass is achieved. Thus, the role of immune markers and bone in childhood may be unique and difficult to interpret, especially given the lack of data in healthy growing children.
Deficits in bone accrual in young men who acquire HIV perinatally or during adolescence 37 have been observed, raising concerns regarding later life osteoporosis and fracture. Larger longitudinal studies with markers of bone resorption and formation and data across Tanner stages are warranted to further elucidate the relationships between immune markers and bone in youth living with HIV.
Footnotes
Acknowledgments
The authors thank the children who participated in this study, their families, and the entire Protocol P1045 team for their contributions and support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Overall support for the International Maternal Pediatric Adolescent AIDS Clinical Trials Network (IMPAACT) was provided by the National Institute of Allergy and Infectious Diseases (NIAID) with cofunding from the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD) and the National Institute of Mental Health (NIMH), all components of the National Institutes of Health (NIH), under award numbers UM1AI068632 (IMPAACT LOC), UM1AI068616 (IMPAACT SDMC), and UM1AI106716 (IMPAACT LC), and by NICHD contract number HHSN275201800001I. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.