
Abstract
Although previous studies have analyzed cross-level CRF01_AE viral genomic data in populations, less is known about intrapatient viral evolutionary dynamics during antiretroviral therapy (ART) failure. We longitudinally sampled plasma and peripheral blood mononuclear cells (PBMC) at different time points from one human immunodeficiency virus type 1 infected patient. The evolution of viral quasispecies was inferred from viral phylogenies. Before treatment, no drug-resistant mutations were found in this patient's plasma, and all viruses had C–C chemokine receptor type 5 (CCR5) tropism. Two months after treatment, the majority of the virus population in plasma and PBMC were drug resistant and X4-tropic. By 5 months after treatment, the viral load increased significantly, and viruses reversed tropism from X4 to R5 in plasma and PBMC. During treatment failure, the effective population of the pol DNA reservoir in PBMC remained stable, whereas the env DNA reservoir increased. The effective population of the R5 tropism virus increased more rapidly than that of the X4 tropism virus. The ratio of non-synonymous to synonymous substitutions in the env gene of R5 tropism virus (0.43) was lower than X4 tropism (0.52). However, four env positive selection sites were identified in R5 tropism viruses (HXB2: 364, 398, 399, and 400) but none were identified in X4 tropism viruses. Our data demonstrated the different intrapatient evolutionary dynamics patterns of env and pol genes in an individual who experienced periods of ART failure. Our findings also suggest the importance of the R5 tropism virus in the DNA reservoir during ART failure.
Background
Because of the high replication rate of human immunodeficiency virus type 1 (HIV-1), the high frequency of recombination among viral genomes, and the lack of proofreading activity of the viral reverse transcriptase (RT), 1 the HIV-1 in infected individuals is present as a swarm of highly related variants (referred to as a quasispecies 2 ). When the selective pressure of antiretroviral therapy (ART) is exerted on such a population, drug-resistant mutants may emerge, which can lead to virological failure. The patterns of emergence and polymorphism of mutations are some of the viral evolutional phenomena still confounding our understanding of HIV-1 drug resistance and persistence in the DNA reservoir. In the pol gene segment, single mutations in RT can result in high-level drug resistance to HIV-1 RT inhibitors such as lamivudine (3TC) and nevirapine (NVP). 3 However, viruses highly resistant to the protease (PR) inhibitor lopinavir require multiple PR mutations. 4
The env gene region of HIV-1 is important, because it is responsible for the capacity of HIV-1 to use coreceptors for entry into host cells. 5 The primary genetic determinant for the HIV coreceptor usage is the third variable region (V3) of the HIV gp120 envelope glycoprotein, encompassing ∼35 residues with a conserved disulfide. This region is subject to high selection pressure. Viruses, especially those using the CCR5 receptor to enter the target cells (R5 viruses), are generally predominant at early stages of HIV-1 infection, whereas the emergence of C–X–C Motif Chemokine Receptor 4 (CXCR4)-using viruses (X4 viruses) generally occurs at later stages. 6 The evolution of R5 to X4 tropism following a host-specific mutational pathway could be influenced by the immune pressure that promotes a rapid host-specific adaptation.
CRF01_AE was introduced from Southeast Asia in the 1990s and has expanded rapidly in China. 7 It has been reported that untreated patients infected with HIV-1 CRF01_AE subtype have a shorter median survival than patients of subtype B8 and that the subtype is associated with X4 rather than R5 tropism, ultimately leading to a more rapid disease progression. 9 Although previous studies have analyzed CRF01_AE viral genomic data for patients who have experienced drug resistance and virological failure during antiretroviral therapy (ART), less is known about the intrapatient evolutionary dynamics of CRF01_AE on viral tropism and drug resistance before and during treatment and virological failure.
To learn more about the evolutionary patterns during ART failure, we longitudinally sampled plasma, peripheral blood mononuclear cells (PBMC) at different time points from a CRF01_AE HIV-1-infected patient who experienced therapy failure. The distribution of resistance mutations and evolution of viral quasispecies in the RT, PR, and env genes were determined by single-genome sequencing and inference from viral phylogenies. The demographic history of the HIV DNA reservoir in PBMC was reconstructed by applying the phylodynamics approach.
Materials and Methods
Patient and ethical statement
A 38-year-old patient was diagnosed to be HIV positive in July 2014. Blood and cerebrospinal fluid (CSF) samples were collected at different time points from July 2014 to December 2016. Written informed consent was obtained from the subject before sample collection.
Samples
We longitudinally sampled plasma, PBMC, and CSF at different time points from an HIV-1-infected patient who had periods of therapy failure. Plasma was separated and stored at −80°C until use. Plasma HIV-1 RNA was extracted by using a QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany). RNA extract was subjected to reverse transcription by using the Superscript II (Invitrogen). PBMC were separated in a Ficoll-Hypaque gradient, and two million PBMC were stored at −80°C until use. Genomic DNA was extracted by using a QIAamp DNA Mini Kit (Qiagen) according to the manufacturer's recommendations.
Single genome sequencing
In-depth pol and env sequencing of RNA and DNA was done by using limiting dilution amplification. The DNA/RNA extract was diluted until no more than 30% of the nested PCR reactions were positive. Primers and PCR conditions were the same as previously published. 10 Fragment sequencing was carried out by using the Big Dye Terminator V.3.0 Cycle Sequencing Ready Reaction kit (ABI). Chromatograms were base called by using the Sequencher (v5.0) program (GeneCodes). Sequences exhibiting nucleotide mixtures, defects, or hypermutations (identified by using the Hypermut program 11 ) were excluded, as were sequences exhibiting evidence of within-host recombination (identified by using the RDP4 program 12 ).
Phylogenetic analysis
MUSCLE software (v3.8.31) 13 was used to align all env and pol sequences from plasma viral RNA and cellular DNA collected at the different collection points—before ART initiation and during the different ART regimens. Alignments were manually edited and trimmed to 326 nucleotides for env (HBX2:7080-7406) and 1,125 nucleotides for pol (HBX2: 2247-3371) by using BioEdit software (v7.0.9). Shorter sequences and sequences with stop codons or gaps larger than a nucleotide triplet were removed from the alignments. The best-fitting nucleotide substitution model was selected with jModeltest software (v2.1.7), 14 using the Akaike Information Criterion (AIC). Phylogenetic trees were inferred by using PhyML software (v3.0). 15 Bootstrap analysis was performed on 1,000 replicates.
Demographic reconstructions
The demographic history of the HIV reservoir in PBMC was estimated by using the BEAST software 16 and implemented in the Bayesian Markov chain Monte Carlo (MCMC) method. The Bayesian skyline model 17 and relaxed molecular clock model with uncorrelated lognormal distribution 18 were incorporated in the MCMC method. Multiple independent MCMC runs were performed and assessed for consistency. Convergence of relevant parameters and Bayesian skyline results were assessed by effective sample sizes over 200 in Tracer v1.6.
Coreceptor tropism analysis
Viral coreceptor tropism was determined by using env sequences and the geno2pheno (coreceptor) algorithm, which have been developed by using training sets with different subtypes. 19 As recommended by the European Consensus Group on clinical management of HIV-1 tropism testing, a false positive rate cutoff of 10% was used for classifying a virus as R5 or X4 tropic. 20
Drug resistance genotyping
Drug-resistance-associated mutations in the PR and RT sequences were sought, and genotyping results were interpreted, by using the drug resistance interpretation beta test (HIV RT and Protease Sequence Database; Stanford University). 21
Selection pressure analysis
Selection pressure was analyzed at the codon level for the env gene in cells infected with the different viral tropisms. The CODEML program in the PAML 4.4 software package was used to apply site-specific models for detecting positive selection. 22 Two selective models that allow for positive selection (2a and 8; ω > 1) were compared with two null models (1a and 7, respectively) that do not allow for positive selection. The likelihood ratio test was used to determine whether there were significant differences between the null model and the alternative model by calculating twice the log-likelihood difference following a χ 2 distribution, with the number of degrees of freedom. 23
Nucleotide sequence accession numbers
All the HIV-1 sequences related to this work have been submitted to GenBank, and they were assigned accession numbers MT575772-MT575993.
Results
Patient treatment
PBMC and plasma samples were collected at different time points from July 2014 to December 2016 (Fig. 1). The patient was diagnosed as HIV positive in July 2014 (−3 months in Table 1), at which point his CD4+ T cell count was 80 cells/uL and plasma HIV viral load 7.9 × 104 copies/mL. Antiretroviral therapy with 3TC+tenofovir (TDF)+NVP was initiated in October 2014 (0 month). Because of a liver adverse reaction, the ART regimen was switched to 3TC+TDF+efavirenz (EFV) 1 month after the first treatment. Before treatment, no drug-resistant mutations were found in the plasma, and all viruses had R5 tropism. One to two months after the first treatment, viral load only decreased to about 2 × 103 copies/mL. Due to virological failure, the ART regimen was again changed to 3TC+TDF+lopinavir and ritonavir (LPV/r), after month 15. In response to the new ART regimen, the viral load dropped to under the detection limit (50 copies/mL).

Schematic representing the treatment and sampling protocols used in this study. This patient initiated antiretroviral therapy with 3TC+TDF+NVP in October 2014 and switched to 3TC+TDF+EFV in November 2014. Then, the ART regimen was changed to 3TC+TDF+LPV/r on February 2016. The month on the initiation of first treatment is regard as month 0. Samples used in the study were collected at different time points shown on top of the schematic. Rectangle represents plasma, and circle represents PBMC. 3TC, lamivudine; EFV, efavirenz; LPV/r, lopinavir and ritonavir; NVP, nevirapine; PBMC, peripheral blood mononuclear cells; TDF, tenofovir.
Characteristics of Sequences Isolated from Plasma
CCR5, C–C chemokine receptor type 5; LOD, limit of detection (50 copies/mL).
Drug resistance genotype
Drug resistance mutation sequencing and analysis was only completed after the entire study period, so they were not used for clinical decisions. The sequences of the pol gene of samples taken before the first 3TC+TDF+NVP initiation showed no drug resistance mutations. Two months after treatment began, the viral load moderately decreased; however, M184V, K103N, and Y181C mutations were already present. M184V mutations confer high-level resistance to 3TC, whereas K103N and Y181C mutations are associated with high-level resistance to NVP as well as to EFV, which has not yet been prescribed. Four months after the second treatment, 3TC+TDF+EFV, viral load increased, with M184V, K103N, Y181C, and G190A mutations associated with high-level resistance (Tables 1 and 2).
Characteristics of Sequences Isolated from Peripheral Blood Mononuclear Cells
PBMC, peripheral blood mononuclear cells.
Tropism switch
Before antiretroviral therapy initiation, the HIV tropism geno2pheno [coreceptor] algorithm predicted that the plasma and CSF contained HIV virus with a predominance of R5 tropism (100%). Over subsequent plasma sample test points, distinguishable dynamics of the HIV tropism were observed. Two months after the first treatment, the majority of the virus population in plasma and PBMC were drug resistant and X4-tropic. Subsequently, the effective population of the R5 tropism virus increased more rapidly than that of the X4 tropism virus. By 5 months after the first treatment, the viral load had increased significantly, and viruses reversed tropism from X4 to R5 in plasma and PBMC. After the third ART regimen, the viral load was suppressed to under the detection limit (50 copies/mL) and the percentage of R5 tropism virus decreased in PBMC (Tables 1, 2 and Supplementary Table S1).
Phylogenetic inferences
Sequences of the pol segment in plasma and PBMC phylogenic tree displayed the development of new clusters, but sequences with no drug resistance mutations still persisted in the HIV DNA (Fig. 2). Unlike the pol sequences, the env phylogenetic tree displayed no consistent clustering of sequences, but rather commingling from different sampling times, indicating the existence of various viral lineages that persisted with the infection. The sequences of env in PBMC DNA 3 months after the first treatment were the same as those 20 months after the first treatment, which indicated the persistence of env in the HIV DNA. The env sequences of DNA 2 months after the first treatment were identical to those of RNA in the plasma 3 months after the first treatment, which indicated that the HIV sequences in plasma may originate from the HIV DNA in PBMC (Fig. 3).
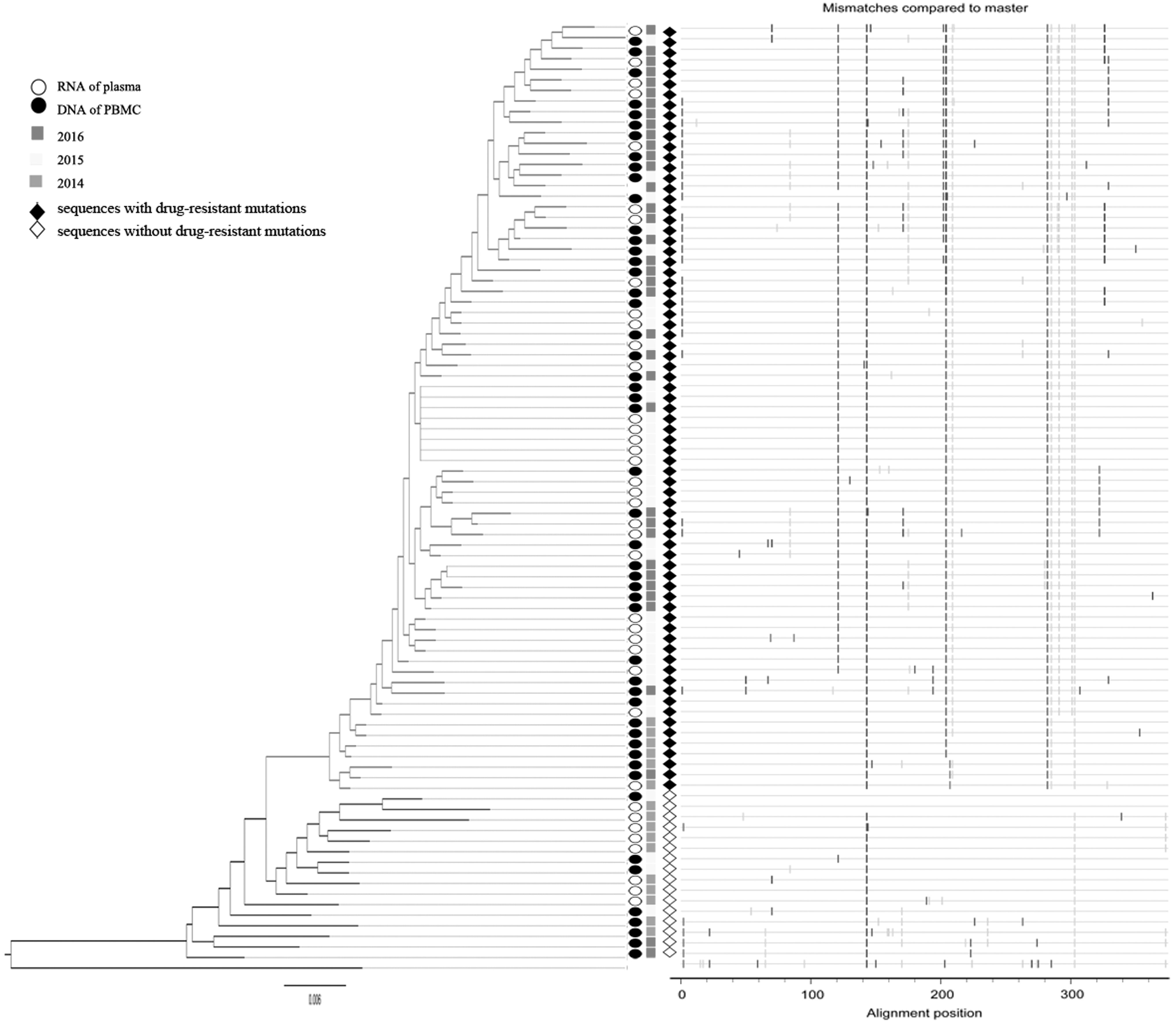
Intrapatient HIV pol diversity in plasma and PBMC. Phylogenies, inferred from nucleotide sequence alignments (HIV pol), are rooted by CM240 (accession: U54771), with scales denoting the estimated number of substitutions per nucleotide site. Matched highlighter plots, made from amino acid sequence alignments, show substitutions relative to the master sequence (one of sequences isolated in 3 months before treatment initiation). Symbols denote sampling location: DNA from PBMC (filled circles) and RNA of plasma (open circles). Different gray shading of squares represent different time of sampling. The filled diamond markers for the sequences with drug-resistant mutations. The open diamond markers for the sequences without drug-resistant mutations.

Intrapatient HIV env diversity in plasma and PBMC. Phylogenies, inferred from nucleotide sequence alignments (HIV env), are rooted by CM240 (accession: U54771), with scales denoting the estimated number of substitutions per nucleotide site. Matched highlighter plots, made from nucleotide sequence alignments, show substitutions relative to the master sequence (one of sequences isolated in DNA of PBMC 1 month after treatment initiation). Symbols denote sampling location: DNA from PBMC (filled circles), RNA of plasma (open circles), and RNA of cerebrospinal fluid (open triangle). The open diamond markers for the CCR5-tropism sequences. The filled diamond markers for the CXCR4-tropism sequences. CCR5, C–C chemokine receptor type 5; CXCR4, C–X–C Motif Chemokine Receptor 4.
Detecting evolution over treatment time points
To infer the dynamic of the size of the HIV reservoir in PBMC, Bayesian skyline plots were reconstructed. The effective population was determined to be the parameter representing the size of the HIV reservoir. The effective population of pol segments was shown to be stable. However, the effective population of env segments underwent a significant increase in genetic variation during the early (2014–2015) period of treatment failure; the X4-tropism virus was responsible for the initiation of that increase in 2014, whereas the R5-tropism virus was responsible for the later increase. This is consistent with the concurrent viral load and tropism in the plasma. The overall effective population of R5 tropism viruses increased more rapidly than X4 tropism viruses (Fig. 4). This finding, combined with the viral history of this patient, suggests that the viral rebound in the plasma could be the driver of the growth in the env effective population in the HIV reservoir in PBMC.

Demographic history of base on pol and env sequences in DNA from PBMC. Bayesian skyline plots showing the viral diversity in the env sequences
Selection pressure analysis
To compare the evolutionary difference between X4 and R5 tropism viruses, we conducted a selection pressure analyses for the env co-receptor gene during the treatment failure. The ratio of non-synonymous to synonymous substitutions in the env gene of R5 tropism virus (0.43) was lower than for X4 tropism (0.52). Four positive env selection sites were identified in R5 tropism viruses, primarily in the V4 loop that is associated with the attachment and incorporation of the virus into the cells (HXB2: 364, 398, 399, and 400) but none were identified in X4 tropism viruses (Table 3).
Non-synonymous/Synonymous Ratios of X4 and R5 Tropism Quasispecies in Peripheral Blood Mononuclear Cells
P-value was calculated by conducting the LRT.
Codons with probability (p > .95) that supports the likelihood of a site having a dN/dS >1.
CXCR4, C–X–C Motif Chemokine Receptor 4; Ln L, log-likelihood; LRT, likelihood ratio test.
Discussion
In this study, viral sequences were isolated over a 29-month period from the plasma and PBMC of an HIV-infected patient who experienced several periods of therapy failure. There was no drug-resistant mutation and no X4-tropism virus found in the plasma before treatment. One month after the beginning of treatment, the majority of the virus exhibited both drug-resistant mutations and X4-tropism. This finding supports previous studies of naive patients who have shown that before treatment there are already many minorities with preexisting mutations in the quasispecies swarm of highly related viral variants, and that this entire viral population can shift from a susceptible to a resistant phenotype in as short duration as 2 to 4 weeks. 3 Previous studies have also shown that the tropism switch is not an “on-off” phenomenon, but it may result from a profound re-shaping of viral quasispecies, even under suppressive ART. 2,24 These facts suggest that the patient's viral load should be determined as early as 1 month after the initiation of treatment.
In this study, distinguishable dynamics of HIV tropism were observed over subsequent test point plasma samples. Because of the differences in coreceptor expression of different CD4 T cell subtypes, the infection of R5 and X4 viruses exhibited different efficiencies. The expression of CXCR4 is usually highest on naive CD4 T cells, followed by central memory, transitional memory cells, and effector memory cells. 25 These cells are also the main source of the HIV DNA reservoir. Due to ART suppression of viremia, the majority of the HIV-infected R5 CD4 T cells will be killed. The ideal target of the HIV is the active R5 CD4 T cell in which the virus can replicate dramatically, and the study found that during the periods of highest viral load in the plasma, almost 100% of viruses were R5 tropic. X4-using variants then became more prevalent after the suppression of viremia. With respect to the evolutionary dynamics of the HIV DNA reservoir in PBMC, this study found significant differences in the pol and env genes' overall effective population in PBMC. Before and during virological failure, the pol effective population variability in the PBMC reservoir remained stable, indicating the replacement of the old drug-resistant variants by new ones. 26 However, the env effective population variability in the DNA reservoir increased during virological failure, indicating that the increased HIV viral load in plasma contributed to the increased HIV DNA reservoir. These findings support previous suggestions that episodes of virological failure seem to prevent the reduction of proviral DNA and to accelerate viral evolution. 24
During treatment failure, the overall effective population of the R5 tropism virus increased more rapidly than X4 tropism virus in PBMC. Selective pressure is less on R5 than on X4 viruses, which is consistent with ultra-deep sequencing results in patients on suppressive antiretroviral therapy. 27 The selective pressure on X4 viruses may enhance env entry efficiency and HIV-1 viral fitness.
Conclusions
Our data demonstrated the different intrapatient evolutionary dynamics patterns of env and pol in an individual infected with HIV-1 CRF01_AE who experienced periods of ART failure. The period of ART failure witnessed not only the emergence of drug resistant mutants but also the switch of tropism. Our findings suggest the importance of R5 tropism virus in the DNA reservoir during ART failure. To fully understand the role of host viral evolutionary dynamics of CRF01_AE viruses during ART, further studies using a larger cohort are required.
Footnotes
Acknowledgment
The authors are grateful to Susan Joyce Herzog for assistance in editing their article.
Authors' Contributions
X.P. performed the analysis and drafted the article. Y.X. performed the experiment. Y.H. conducted epidemiology investigations and collected samples. B.Z. supervised and administered this project. All authors read and approved the final article.
Ethics Approval and Consent to Participate
The study was approved by the institutional review boards of the First Affiliated Hospital, School of Medicine, Zhejiang University (reference number: 2020265). All subjects signed an informed consent form before collecting samples.
Availability of Data and Materials
The datasets used in this study are available from the corresponding author on reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the Medical Health Science and Technology Project of Zhejiang Provincial Health Commission (no. 2020KY137) and the Independent Task of State Key Laboratory for Diagnosis and Treatment of Infectious Diseases (no. 2019ZZ18).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.