
Abstract
Coccidioidomycosis is a common fungal infection in people living with HIV-1, particularly in southwest regions of the United States where the Coccidioides sp. is endemic, but rates of infection have significantly declined in the era of potent combination antiretroviral therapy (cART). Natural coccidioidomycosis also occurs in outdoor-housed macaques residing in the southwestern states that are utilized in biomedical research. Here, we report on a recrudescent case of previously treated, naturally occurring coccidioidomycosis in a pigtail macaque that was experimentally infected with simian immunodeficiency virus (SIV) and virally suppressed on cART. Coccidioides IgG antibody titer became detectable before discontinuation of cART, but symptomatic coccidioidomycosis developed subsequent to cART withdrawal. This animal was screened and treated in accordance with the guidelines for the prevention and treatment of coccidioidomycosis, suggesting that macaques with a history of coccidioidomycosis should be excluded from enrollment in HIV studies. Continual monitoring for known endemic pathogens based on the colony of origin is also recommended for animals utilized for HIV/AIDS research.
Coccidioidomycosis is caused by a soil-dwelling fungus and is highly endemic in the southwest regions of the United States. Most human coccidioidomycosis cases result in pneumonia, and account for up to 30% of community-acquired pneumonia in endemic areas. 1,2 Symptomatic coccidioidomycosis is more common in people living with HIV (PLWH) if there is progression to AIDS, productive HIV viral replication, and/or depletion of CD4 T lymphocyte counts below 250 cells/μL of blood. 2,3 However, potent combination antiretroviral therapy (cART) has significantly reduced the incidence of coccidioidomycosis in PLWH. 3
Coccidioidomycosis has also been documented in outdoor-housed macaques utilized for biomedical research in the southwestern states. 4 The Washington National Primate Research Center's (WaNPRC) breeding colony in Mesa, Arizona, which houses pigtail macaques (PTM) (Macaca nemestrina) in a mixed indoor/outdoor setting, reported a prevalence of 12% in 2017 (personal communication C.M.M.). Notably, the PTM model of HIV has gained popularity because aspects of simian immunodeficiency virus (SIV) infection in this species more closely resemble that of humans infected with HIV-1, compared with other species of macaques. 5 To date, coccidioidomycosis in SIV-infected macaques has not been described. Here, we report recurrence of previously treated, naturally occurring coccidioidomycosis in a PTM that was experimentally infected with SIV and virally suppressed on cART. Coccidioidomycosis IgG titer became detectable before discontinuation of cART, but symptomatic coccidioidomycosis developed subsequent to cART withdrawal.
A 5-year-old intact male PTM was enrolled in a study evaluating a novel hepatitis B virus vaccine for use in PLWH. All experimental procedures performed on this macaque were approved by the University of Washington IACUC. In December 2017, 2 years before study enrollment, the macaque presented with coughing while at the WaNPRC Arizona colony. A diagnosis of coccidioidomycosis was based upon positive IgM (1:4) and IgG (1:8) antibody titers (ProtaTek Reference Lab, Mesa, AZ). As per standard protocol at the WaNPRC, and in accordance with the prevention and treatment guidelines for coccidioidomycosis, 2 long-term fluconazole therapy (10 mg/kg once daily; Zydus Pharmaceuticals, Pennington, NJ) was initiated until the animal was confirmed to have at least three negative IgG/IgM antibody titers over a 1-year period. Coughing resolved within 1 month of treatment, and follow-up titers indicated decreasing IgG and IgM antibody titers that were undetectable by September 2018, with a final negative titer in September 2019 (Fig. 1A, B).
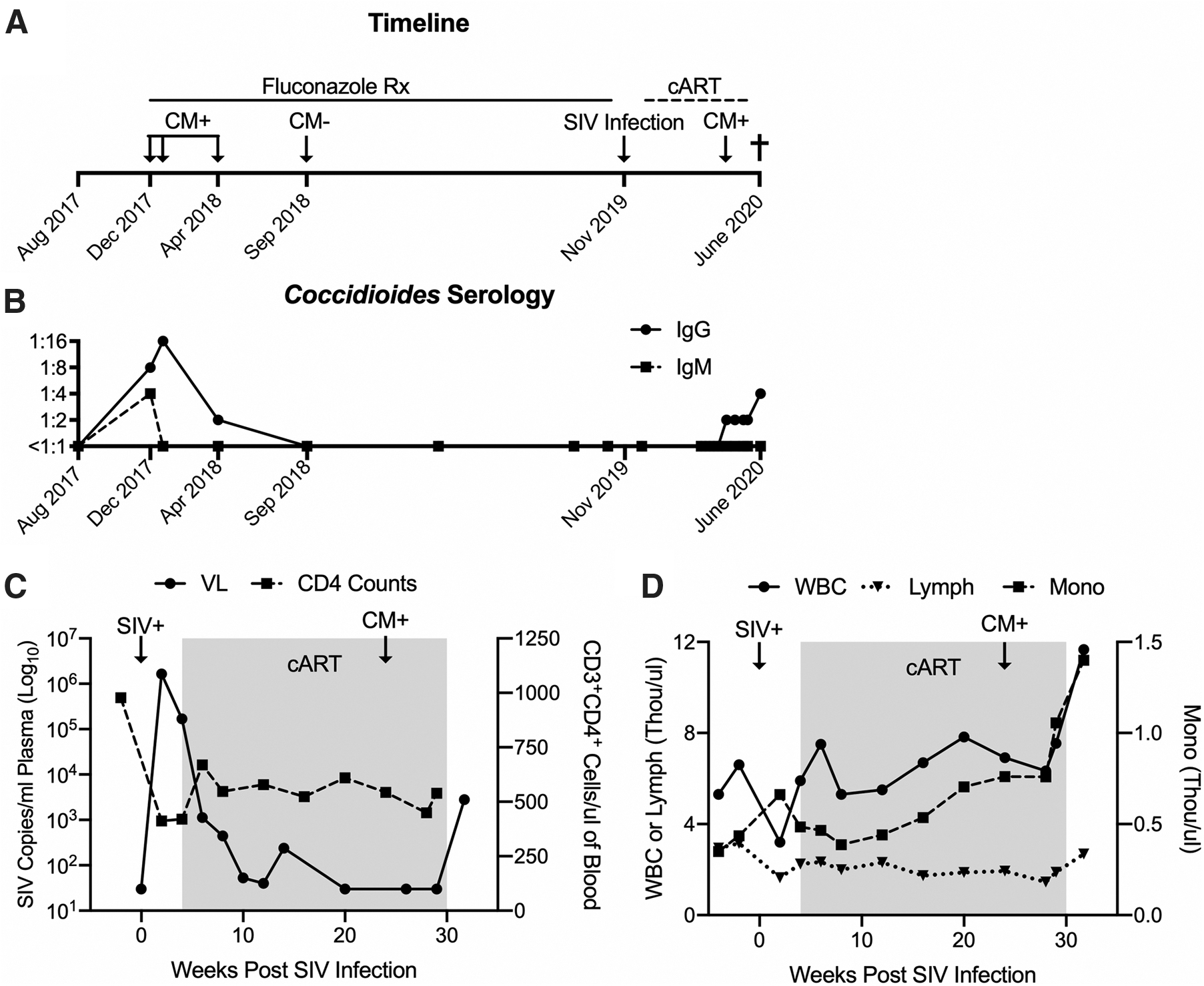
Re-emergence of naturally acquired coccidioidomycosis (CM) in an SIV virally-suppressed, immunologically intact PTM.
Three months before study enrollment, the macaque was relocated to the WaNPRC Seattle site, an AAALAC international-accredited institution, and maintained in a strictly indoor setting. The macaque was intravenously inoculated with 10,000 infectious units of SIVmac239M 6 in November 2019. Daily cART consisting of 5.1 mg/kg tenofovir disoproxil fumarate (Gilead Sciences, Foster City, CA), 2.5 mg/kg dolutegravir (ViiV Healthcare, Research Triangle Park, NC), and 30 mg/kg emtricitabine (Gilead Sciences) was administered subcutaneously from 4 to 30 weeks post-SIV infection (Fig. 1A). Blood was collected before SIV infection and every other week thereafter for complete blood count (CBC), subset analysis, biochemistry, or viral load evaluation.
During acute SIV infection, baseline CD4 counts (978 cells/μL) declined to as low as 412 cells/μL and peak SIV viremia reached 6.2 Log10 copies/mL of plasma. During the period of cART, CD4 counts remained stable (449–669 cells/μL) and plasma viremia was <30 copies/mL by 10 weeks on cART (Fig. 1C). CBC analyses were unremarkable during acute SIV and cART (Fig. 1D). Approximately 2 weeks after cART withdrawal, the macaque presented with dyspnea marked by open-mouth breathing and audible wheezing, and was sedated with intramuscular ketamine (10 mg/kg; Covetrus, Dublin, OH) for evaluation. Oxygen saturation was 99% on flow-by oxygen, but rapidly decreased to 85% on room air. The macaque was administered intravenous midazolam (0.5 mg/kg; Heritage Pharmaceuticals, Eatontown, NJ) and intubated to facilitate ventilation, but the animal's respiration could not be stabilized. Due to grave prognosis, the animal was euthanized with intravenous Nembutal (160 mg/kg; Virbac AH, Westlake, TX) and necropsied.
Necropsy revealed a 1–1.5 cm3 white and brown multinodular firm mass located in the tracheal lumen approximately 1 cm cranial to the bifurcation (Fig. 2A, B). This mass was contiguous with a 2–3 cm3 similar mass located ventral to the trachea, with extension through and direct connection to the luminal tracheal mass. On section, the lesion consisted of coalescing brown-tan nodules exuding moderate amounts of viscous material, and with white firm reticulation. Representative tissues and organs were fixed in 10% formalin, embedded in paraffin, sectioned, and then hematoxylin and eosin stained.
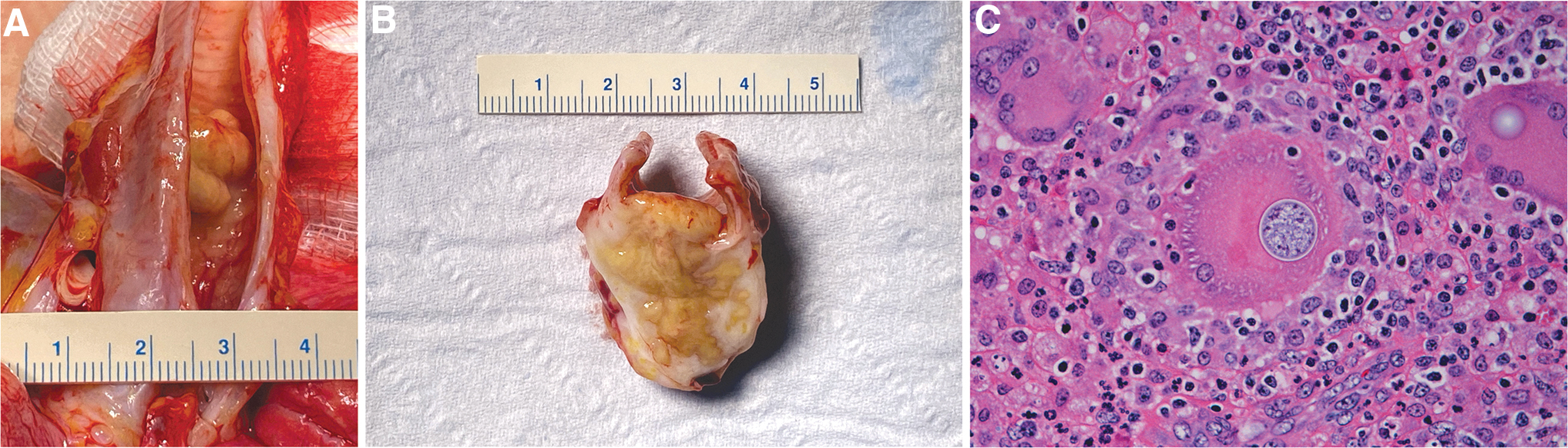
Coccidioides sp.-associated tracheal lesion gross pathology and histology.
Sections of trachea and associated structures (jugular veins, carotid arteries, paratracheal lymph nodes) had effacing inflammation apparently arising from a paratracheal lymph node that was sclerosing and pyogranulomatous with pyogranulomas, large numbers of giant cells, and moderate numbers of fungal spherules consistent with Coccidioides sp: spherules ranged from 10 to 40 μm with a thin refractile capsule and contained few to numerous 2–5 μm endospores (Fig. 2C). Transmural inflammation extended into the tracheal lumen with ulceration and partial occlusion. Where not ulcerated, the tracheal epithelium was attenuated to hyperplastic and had submucosal granulomatous and eosinophilic inflammation. There also were regional vasculitis and cellulitis. Fite's acid fast stain was negative.
Lungs had moderate multifocal areas with peribronchial granulomatous and eosinophilic inflammation with giant cells and tract formation, multiple moderately large areas of effacing, similar alveolar inflammation, and with occasional extension of inflammation extending into bronchioles. Other tissues and organs were unremarkable besides moderate inflammatory bowel disease unrelated to coccidioidomycosis. Interpretations were chronic, occlusive, recrudescent tracheal and peritracheal coccidioidomycosis arising from a quiescent residual infection of a paratracheal lymph node. The pneumonia was subacute and subsequent to aspiration of exudate and fungal organisms, and likely also had a secondary bacterial component.
Cryopreserved plasma retroactively submitted for IgM and IgG Coccidioides titers indicated a low but detectable IgG titer in April 2020, approximately 8 weeks before discontinuation of cART (Fig. 1A), which continued to increase thereafter despite stable CD4 counts (400–700 cells/μL) and undetectable SIV viral load (Fig. 1C). CD4 counts were not evaluated at euthanasia, however, CD3+CD4+ T cells in blood mononuclear cells (19.4%) by flow cytometry were consistent with levels in 5 other SIV-infected PTMs enrolled in the same study at the same time point (18.8 ± 6.05) (data not shown), suggesting that further CD4 decline did not occur in this animal after cART withdrawal. Increases in white blood cell and monocyte counts (Fig. 1D) and globulin levels (4.6 g/dL) (data not shown) corresponded to resurgence of detectable Coccidioides IgG titer, however, only after cART withdrawal (Fig. 1D).
Since the advent of potent cART, the incidence of symptomatic coccidioidomycosis in PLWH has significantly declined, 3 and case reports of symptomatic coccidioidomycosis are primarily associated with therapeutic noncompliance. 7 –9 For PLWH in regions with endemic Coccidioides, one to two serologic tests per year are recommended, and treatment is only implemented with positive serology and until CD4 counts are >250 cells/μL. 2 Similarly, this macaque was screened for coccidioidomycosis at least twice per year and CD4 counts remained above 250 cells/μL during the course of the experimental study. Recurrence of coccidioidomycosis following antifungal therapy is rare in humans and often due to exposure to other Coccidioides sp. strains. 10 However, because Coccidioides fungal spores are transmitted by inhalation from the environment, rather than between animals and humans, 1,4,11 and as this macaque was strictly housed indoors in Seattle, which is nonendemic for the fungus, exposure to another strain is unlikely. CD4 T helper cells, including Th1 and Th17 subsets, are important for Coccidioides control. 12 The SIV infection may have impaired Th1 and Th17 responses in this animal and likely reduced its ability to keep the fungal infection suppressed and contributed to recrudescence of the initial infection.
To reduce confounding variables in HIV/AIDS research, specific pathogen-free (SPF) macaque colonies that are absent of simian retrovirus type D (SRV-D), simian T lymphotropic virus-1 (STLV-1), SIV, and macacine herpesvirus-1 (McHV-1) are necessary. 13 –15 However, screening for other pathogens [e.g., rhesus rhadinovirus (RRV), cytomegalovirus (CMV), lymphocryptovirus (LCV), and simian foamy virus (SFV)] may also be recommended. Furthermore, naturally transmitted pathogens endemic to specific regions of the United States, such as Chagas disease (Trypanosoma cruzi) and histoplasmosis (Histoplasma sp.), have been reported to recur in the macaque species enrolled in HIV/AIDS research. 16 –18 Coccidioidomycosis has been documented in other nonhuman primate (NHP) species used for HIV/AIDS research, including rhesus macaques (Macaca mulatta) and sooty mangabeys (Cercocebus atys). 4,19,20 To our knowledge, this is the first case report documenting recrudescence of naturally occurring coccidioidomycosis in an NHP enrolled in HIV/AIDS-related research. This case report emphasizes the importance of coccidioidomycosis screening in all NHPs planned for enrollment into HIV/AIDS research when there is potential for fungal exposure, particularly NHPs originating from outdoor colonies in endemic areas. In addition, NHPs with a history of coccidioidomycosis, regardless of successful antifungal treatment and negative follow-up Coccidioides titers, should be excluded from enrollment in SIV/SHIV studies.
Footnotes
Acknowledgments
The authors thank Solomon Wangari, Naoto Iwayama, Chul Ahrens, and William Garrison for outstanding NHP support, and Gilead Sciences and Viiv Healthcare for providing cART. They thank Brandon Keele for providing the SIVmac239M. They also thank the University of Washington Virology & Immunology Core, Sandra Dross, Bei Ming, and Jessica Li for plasma viral load assays and CBC subset analysis, the University of Washington Department of Laboratory Medicine for serum chemistries, and Amirah Ullah and Erin Broderick for technical support. This study was supported by grants from the National Institutes of Health (NIH), the National Institute of Allergy and Infectious Diseases (R56 AI141494 to DHF), and the National Institute of Mental Health (K01 MH1235258 to MAO). The WaNPRC is supported by grant P51 OD010425 from the NIH Office of Research Infrastructure Programs.
Authors' Contributions
Methodology: K.A.G., R.D.M., T.B.L., B.B., A.B., D.A.J., C.M.M., and M.A.O. Data analysis: K.A.G., R.D.M., T.B.L., B.B., A.B., and M.A.O. Writing—original draft preparation: K.A.G. and M.A.O. Writing—review and editing: K.A.G., R.D.M., T.B.L., B.B., A.B., D.A.J., C.M.M., D.H.F., and M.A.O.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was funded by the National Institutes of Health (NIH) grants R56 AI141494 and P51 OD010425. M.A.O. was supported by NIH K01 MH1235258.