
Abstract
In the pilot NCT01935089 trial, we tested whether pegylated interferon alpha2b (Peg-IFN-α2b) with antiretroviral therapy (ART) was safe and could impact HIV and immune measures in blood and in gut-associated lymphoid tissue (GALT). Twenty HIV-1+ ART-suppressed individuals received 1 μg/kg/week Peg-IFN-α2b with ART for 20 weeks, with intermediate 4-week analytical ART interruption (ATI). Safety, immune activation, HIV viral load and integrated HIV DNA in blood, and HIV RNA and DNA in gut biopsies were measured. A total of 7/20 participants experienced grade 3–4 adverse events, while 17/20 participants completed the study. Of the 17 participants who completed the study, 8 remained suppressed during ATI, while all 17 were suppressed at end of treatment (EoT). As expected, treatment increased activation of T and natural killer (NK) cells and IFN-stimulated molecule expression on monocytes in periphery. While circulating CD4+ T cells showed a trend for a decrease in integrated HIV DNA, GALT showed a significant decrease in HIV-1 RNA+ cells as measured by in situ hybridization along with a reduction in total HIV DNA and cell-associated RNA by EoT. The observed decrease in HIV-1 RNA+ cells in GALT was positively associated with the decrease in activated NK cells and macrophages. This study documents for the first time that 20 weeks of immunotherapy with Peg-IFN-α2b+ART (inclusive of a 4-week ATI) is safe and results in an increase in blood and GALT immune activation and in a significant decrease in HIV-1 RNA+ cells in GALT in association with changes in innate cell activation.
Introduction
Viral reservoirs are maintained in HIV infected individuals despite antiretroviral therapy (ART) creating an almost insurmountable challenge to HIV eradication and cure. 1,2 Reservoirs exist in many cell types, but those in CD4+ T cells are the best characterized. 3 –5 In addition to lymph nodes and tonsils, the gastrointestinal mucosa is a major site of tissue-associated lymphocytes where persistent HIV RNA expressing cells have been described despite undetectable viral levels in blood. 6 –8 Thus, HIV eradication and/or functional cure approaches require strategies aimed at reducing or eliminating latent HIV reservoirs located in all body compartments.
ART does not alter viral rebound kinetics 9 supporting the need to develop immunologic interventions to achieve ART-free virologic remission. Type-I interferons (IFNs) have been proposed as a potential immunotherapy against persistent HIV on ART due to their ability to activate antiviral programs in vivo and to enhance natural killer (NK) and T cell antiviral responses. 10 Our previous data support that the frequency of IFN-α producing plasmacytoid dendritic cells (PDCs) on ART is associated with set-point plasma HIV viral load (VL) after treatment interruption, 11 viral control, 12 and in vitro NK cell lysis of autologous HIV-1 infected CD4+ primary T cells through a mechanism dependent on IFN-α/β receptor (IFNAR) activity. 13 Importantly, cell-meditated responses may require a level of viral reactivation [e.g. using analytical ART interruption (ATI)] to trigger the recognition of latently infected cells.
The antiviral activity of type-I IFNs has been well documented to mediate antiviral effects against HIV-1. 14 –17 Our previous clinical trial data in ART-suppressed individuals (NCT00594880) also support that in vivo administration of pegylated IFN-α2a (Peg-IFN-α2a, Pegasys) in the presence of fully suppressive ART, followed by ATI, 18 can mediate in a subset of individuals a stable viral suppression and a reduction in integrated proviral HIV DNA in periphery in association with greater IFN-induced NK cell cytotoxicity. 19 Additional studies on Peg-IFN-α2a immunotherapy in hepatitis C virus (HCV)/HIV coinfected individuals have confirmed the observation of a reduction in proviral HIV DNA levels in association with NK cell activation. 20 –22
The use of IFN-α2b as HIV immunotherapy and its effects on immune activation and tissue viral measures on ART have not been tested. In this study, we report the findings of a pilot study focused on the safety and the immune and antiviral effects of 20 weeks of dosing with Peg-IFN-α2b as immunotherapy added to ART.
Materials and Methods
Clinical trial design
The NCT01935089 pilot study (
Twenty-two HIV-1+ individuals were screened. Eligible participants were 18 to 65 years old, with weight between 125 and 300 lbs, on suppressive ART for >1 year and at screening, with plasma HIV-1 RNA (VL) <50 copies/mL for ≥1 year and at screening, CD4+ T cell count ≥450 cells/mm3 at screening, and a normal electrocardiogram (ECG) or a Framingham score >15% (men) or 10% (women). Exclusion criteria included HCV coinfection, active hepatitis B virus infection, history of major depression or ongoing moderate depression determined by PHQ-9, pregnancy, and history of autoimmune diseases, retinopathy or clinically significant ophthalmologic disease, or pathological conditions that would contraindicate the administration of type-I IFNs. Figure 1A shows a schematic representation of the study design. A narrative description of schema and sample allocation per study methods is included in Supplementary Methods.

Study schema and disposition.
Phenotypic characterization of peripheral blood mononuclear cells
Immunophenotypic characterization by flow cytometry of innate and adaptive cell subsets was performed on cryopreserved leukapheresis-derived peripheral blood mononuclear cells (PBMC) collected at baseline (BL) and end of treatment (EoT) using seven-color staining as previously described. 11,23
Antibody staining assessed markers of activation/inhibition (CD69, HLA-DR, CD38, killer cell immunoglobulin like receptors, killer activation receptors) on NK cell subsets. 24,25 Expression of IFN-αR subunit 2 (IFN-αR2) was also assessed in NK cell subsets based on our previous data showing a reduction in integrated proviral HIV DNA in periphery in association with greater IFN-induced NK cell cytotoxicity, 19 as well as a role of IFN-α/β receptor 2 (IFNAR2) activity in the in vitro NK cell lysis of autologous HIV-1 infected CD4+ primary T cells. 13 Furthermore, antibody staining assessed immunomodulatory molecules [programmed cell death (PD) ligand 1 (PD-L1), PD-L2] and IFN-stimulated proteins (Tetherin, CD169) on dendritic cell (DC) subsets, 26 monocytes, and B cells. Finally, IFN-stimulated proteins (Tetherin) and markers of activation (e.g., CD38, HLA-DR, PD-1) were assessed on T cells. Results were expressed as mean fluorescence intensity (MFI) and percent positive (%). Details are included in Supplementary Methods.
Assessment of soluble markers of immune activation
Plasma levels of IL-6, CD14, CD163, D-dimer, and C-reactive protein (CRP) were measured at BL and EoT by enzyme-linked immunosorbent assay (ELISA) as per manufacturer's specifications (IL-6, CD14, and CRP: R&D Systems, Inc., Minneapolis, MN; CD163: IQ Products, Groningen, NL: D-dimer: Life Technologies). All measurements were based on the average of duplicate undiluted samples. Lower limits of detection for IL-6, CD14, CD163, D-dimer, and CRP were 3.13 pg/mL, 250 pg/mL, 0.39 ng/mL, 0.08 pg/mL, and 0.78 ng/mL, respectively.
Gut-associated lymphoid tissue specimen collection, processing, and phenotypic characterization
A total of 12–16 rectal tissue biopsies were collected at BL and EoT by Fiber optically guided flexible sigmoidoscopy, with 2 biopsies being placed in 4% neutral-buffered, fresh paraformaldehyde for preparation of paraffin-embedded tissue and 10–14 biopsies being used for mononuclear cell isolation for same day flow cytometry and cryopreservation. A 3 and a 15-color staining panel were used to measure (1) activation or resting status of T cells based on the coexpression or not of CD25, CD69, and HLA-DR and (2) T cell memory and NK cell subsets, and their activation, and functional status (CD69, HLA-DR, perforin), as well as activation (CD69, HLA-DR) and immunomodulatory molecules (PDL1) on macrophages. Results were expressed as MFI and % positive. Details are included in Supplementary Methods.
Integrated HIV DNA in PBMC
Integrated HIV DNA was assessed during follow-up by Alu-gag-based polymerase chain reaction (Alu-gag-PCR) performed on CD4+ T cells isolated from whole blood-derived PBMC by negative magnetic selection as previously described. 27,28 Results were expressed as copies of integrated HIV DNA per 106 CD4+ T cells.
Total HIV RNA in paraffin-embedded tissue sections
Rectal biopsies collected at BL and EoT (two biopsy specimens per time point) were used for preparation of paraffin embedded tissue blocks. Adjacent sections of these blocks were used to assess the location of CD4+ T cells and CD68+ macrophages by immunohistochemistry and to detect HIV RNA with HIV-specific Sulfur-35 labeled antisense probes, according to a previously reported in situ hybridization technique. 29,30 Results were expressed as HIV RNA+ cells/mm2.
Total HIV DNA and cell-associated HIV RNA in rectal mucosa-derived cells
Cryopreserved mononuclear cells derived from rectal mucosal biopsies collected at BL and EoT were evaluated for total HIV DNA and cell-associated RNA using a qPCR TaqMan assay as previously described. 31 Specimens were assayed in replicate reaction wells and the copy number determined by extrapolation against a seven-point standard curve (1–10,000 cps) performed in triplicate. Details are included in Supplementary Methods.
Statistical analysis
Data are described as (1) medians and interquartile ranges (IQRs) and (2) mean and standard deviation (STDEV). For analysis and graphing purposes, plasma HIV-1 RNA <50 copies/mL was considered as equal to 50 copies/mL (threshold of detection). Differences between BL and EoT were tested, using Wilcoxon Signed-Rank or paired t-tests depending on data distribution assessed by the Anderson-Darling normality test. Due to the pilot nature of the study p values were unadjusted, with p values <0.05 considered as significant. Associations were tested by Spearman correlation test. Statistics was performed with JMP Pro11 (SAS Institute, Cary, NC).
Results
Study disposition, safety data, and suppression of HIV-1 RNA after a 4-week ATI
As shown in Figure 1A, the period of 20 weeks of Peg-IFN-α2b immunotherapy included a 4-week ATI to reflect a partial comparative design to the previous NCT00594880 clinical trial design, 18 where a decrease in integrated HIV DNA was detected after 17 weeks of immunotherapy with Peg-IFN-α2a inclusive of a longer (4–12 weeks) ATI. Study disposition for the pilot study is summarized in the consolidated standards of reporting trials (CONSORT) flow diagram shown in Figure 1B. Of the 22 HIV-1+ screened individuals, 2 did not fulfill eligibility criteria. Table 1 describes BL characteristics of the enrolled 20 participants.
Study Participants Baseline Characteristics
ANC, absolute neutrophil count; ALT, alanine aminotransferase; AST, aspartate aminotransferase; WBCC, white blood cell count.
None of the enrolled participants was lost to follow-up, and no suspected unexpected serious adverse reaction (SUSAR) was reported. As shown in Figure 2, changes in CD4+ T cell levels after the ATI remained stable. Illustrating expected biological effects of immunotherapy, 1/20 participants experienced a grade 2 adverse event (i.e., depression), and 7/20 participants experienced grade 3 and higher serious adverse events (SAEs), [i.e. decrease in neutrophil counts (neutropenia)] after start of IFN-α2b immunotherapy. In total three participants discontinued prematurely the study drug (grade 4 neutropenia, n = 2; grade 2 depression, n = 1; Fig. 1B). Finally, 47.1% (8/17) of participants who completed 20 weeks of treatment remained suppressed during the 4-week ATI while receiving Peg-IFN-α2b, while all 9/17 participants who experienced plasma HIV-1 RNA ≥50 copies/mL [median (IQR): 163 (20–2,380.5) copies/mL] returned to HIV-1 RNA <50 copies/mL upon ART reinitiation while continuing Peg-IFN-α2b.
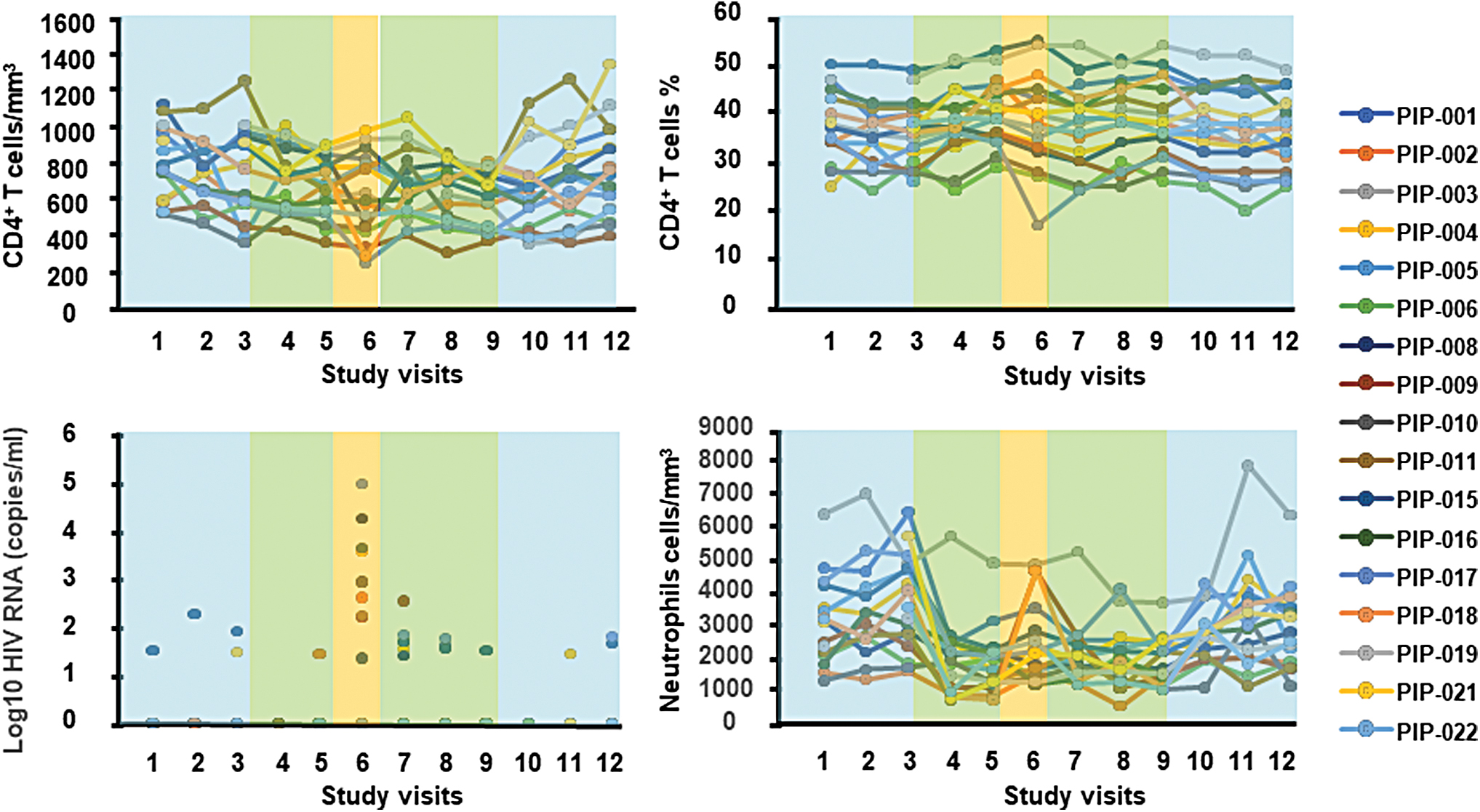
Clinical measurements during follow-up. CD4+ T cells/mm3, CD4+ T cell percentage (%), log10 HIV-1 RNA copies/mL, and neutrophil cells/mm3 levels during follow-up for study participants. Color lines indicate each participant. As described in Fig. 1A, treatment is represented by blue boxes (ART), green boxes (combined administration of ART and Peg-IFN-α2b), or orange boxes (ATI, Peg-IFN-α2b monotherapy). Color images are available online.
Evidence for treatment-induced levels of immune activation and IFN-stimulated molecules in blood and gut-associated lymphoid tissue after 20 weeks of treatment
Changes in the adaptive and innate compartment in blood and gut-associated lymphoid tissue (GALT) were assessed in available paired samples from participants who completed 20 weeks of treatment (Fig. 3). In blood, completion of 20 weeks of treatment resulted in a decrease in peripheral blood CD4+ T cell count [mean (STDEV) for BL and EoT: 717.23 (247.22) and 610 (148.39); p = .0339] (Fig. 3) consistent with expected effects of long-term therapy with Peg-IFN-α2b, suggesting an effect. As summarized in Table 2 and Figure 3, evidence of immunotherapy-associated immune modulation in the blood was given by the increased levels of multiple T and NK cell activation markers detected by 20 weeks of treatment [e.g. CD8+CD38+ % (p < .0001), CD8+PD1+ % (p = .0034), MFI of CD38 on CD56bright (p = .0023), and CD56dimCD16+ (p < .0001)]. Among the soluble markers of immune activation measured in plasma, only sCD163 was detected to increase by 20 weeks of treatment (p = .0174). Consistent with IFN-mediated effects, an increase in the levels of IFN-stimulated molecules was detected in several leukocyte subsets by 20 weeks of treatment [e.g. % of Tetherin+ CD8+ (p = .0017) and CD4+ T cells (p = .0016); MFI of Tetherin (p = .002) and CD169 (p = .0003) on CD14+ cells].
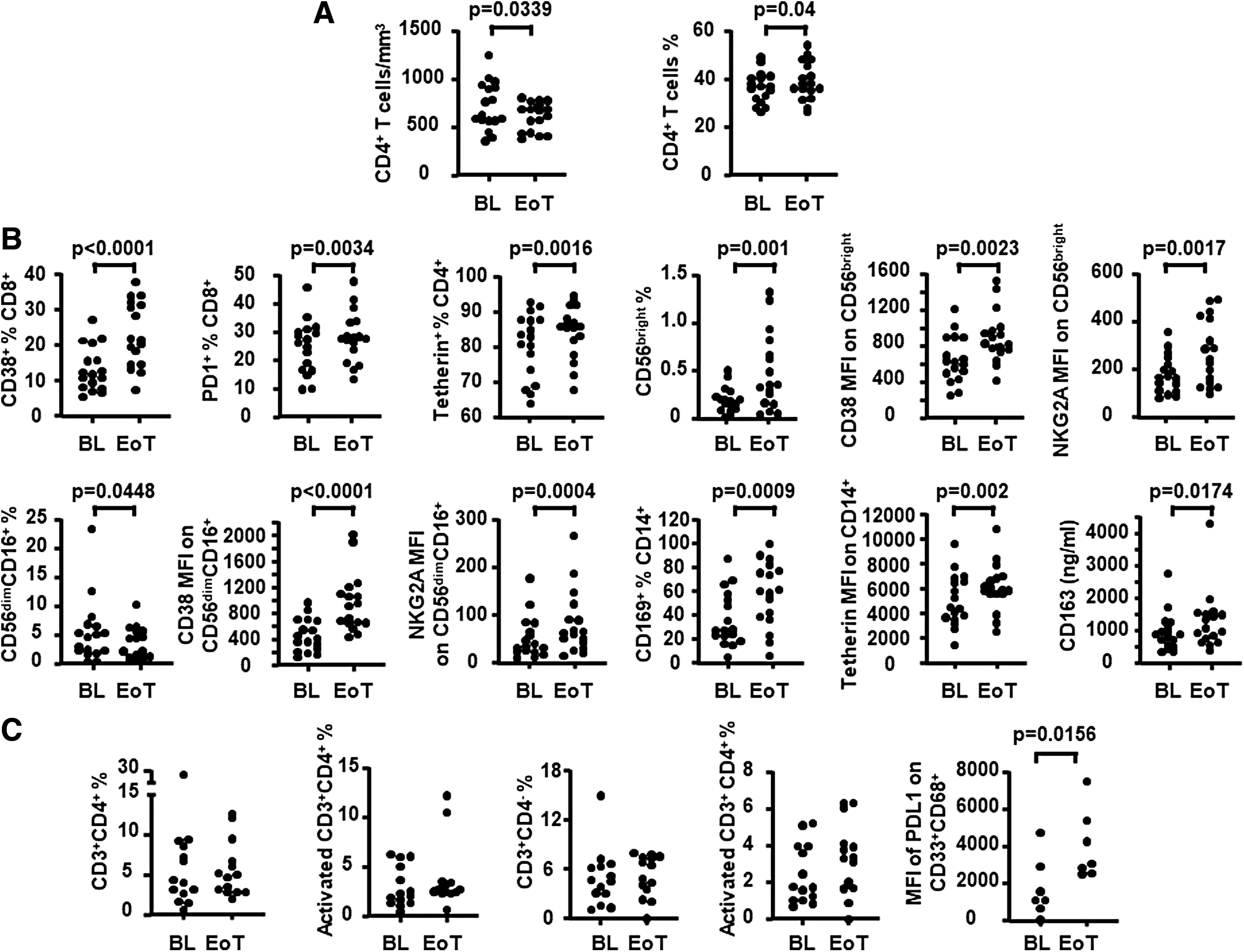
Changes in immune variables in blood and in GALT after 20 weeks of combined administration of ART and Peg-IFN-α2b (inclusive of a 4-week ATI period).
IFN-α-Mediated Changes in PBMC After 20 Weeks of Dual Treatment with Peg-IFN-α2b and ART in the Presence of HIV Reactivation Through ATI
Statistics was performed with JMP Pro11 (SAS Institute, Cary, NC). Briefly, differences between BL and EoT were tested, using Wilcoxon Signed-Rank or paired t-tests depending on data distribution. p Values <.05 were considered statistically significant (unadjusted).
B cells were defined as CD19+BDCA1+.
PDC were defined as CD14−BDCA2+BDCA4+.
MDC were defined as CD19-BDCA1+CD11c+.
Monocytes were defined as CD14+BDCA2−BDCA4−.
ART, antiretroviral therapy; ATI, analytical ART interruption; BL, baseline; EoT, end of treatment; IFN-αR, IFN-α receptor; MFI, mean fluorescence intensity; NK, natural killer; NKG2A, CD94/NK group 2 member A; PBMC, peripheral blood mononuclear cells; PD1, programmed cell death 1; PDC, plasmacytoid dendritic cells; Peg-IFN-α2b, pegylated interferon alpha2b; MDC, Myeloid dendritic cells.
In GALT, no change was found after immunotherapy in the frequency of tissue CD4+ or CD4− T cells, but immune activation changes were detected consistent with blood-associated changes (Table 3 and Fig. 3). Specifically, an increased expression of activation markers was detected in T and NK cell subsets [e.g. MFI of CD69 on CD8+ T cells (p = .0313) and CD56bright NK cells (p = .0391), MFI of perforin on CD8+ T cells (p = .0156) and CD56bright NK cells (p = .0156)]. Detection of an increase in PDL-1 expression on macrophages (e.g., MFI of PDL-1: p = .0156) was also consistent with expected changes as a result of type-I IFN-mediated modulation of myeloid cells. 32,33
IFN-α-Mediated Changes in GALT After 20 Weeks of Dual Treatment with Peg-IFN-α2b and ART in the Presence of HIV Reactivation Through ATI
Statistics was performed with JMP Pro11 (SAS Institute). Briefly, differences between BL and EoT were tested, using Wilcoxon Signed-Rank or paired t-tests depending on data distribution. p Values <.05 were considered statistically significant (unadjusted).
Macrophages were defined as CD3−CD68+CD33+.
GALT, gut-associated lymphoid tissue.
Integrated HIV DNA in circulating CD4+ T cells and GALT changes in HIV DNA and RNA after 20 weeks of treatment
The study end point was the change in the number of copies of integrated HIV DNA/106 CD4+ T cells in periphery after 20 weeks of dual treatment with Peg-IFN-α2b Peg-IFN-α2b+ART, which showed a downward trend (p = .0797, Fig. 4A). The levels of total HIV DNA in mononuclear cell suspensions from GALT biopsies showed a modest significant reduction at EoT compared to BL (p = .05) (Fig. 4B). In contrast to HIV DNA, a significant decrease was detected in the number of HIV-1 RNA+ cells (p = .0151) as directly measured in gut tissue using in situ hybridization (Fig. 5). Specifically, of the 15 participants with available tissue samples at both BL and EoT, 11 showed a clear decline [median (IQR): −4.33 (−12.56, −0.93) HIV RNA+ cells/mm2], 1 showed a 7.5 HIV RNA+ cells/mm2 increase, and 3 showed marginal change at EoT [median (IQR): 0.07 (0.05, 0.1) HIV RNA+ cells/mm2]. These data were further supported by a reduction at EoT of the levels of cell-associated RNA in mononuclear cell suspensions from GALT biopsies (p = .06, data not shown).
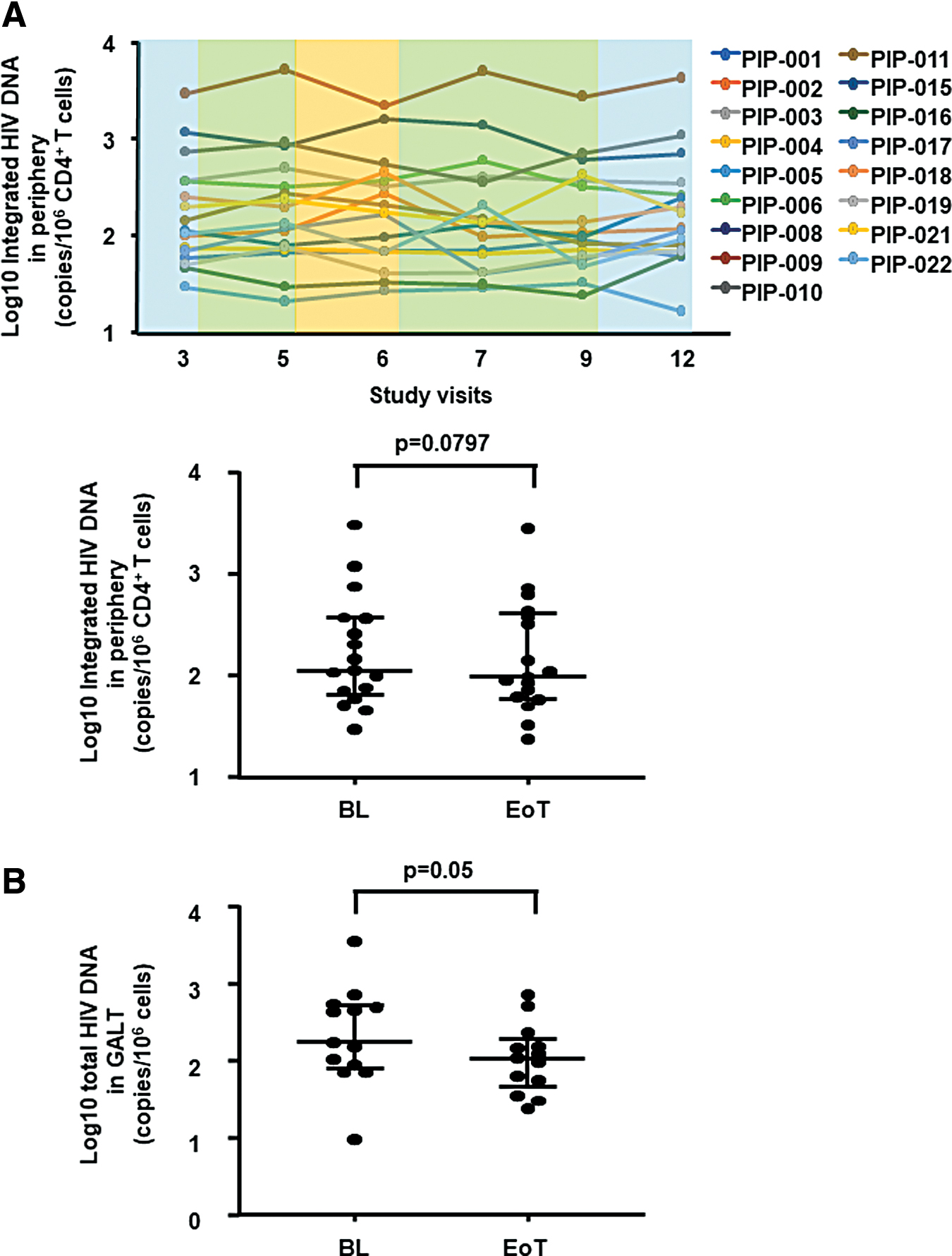
Changes in HIV DNA in blood and in GALT after 20 weeks of combined administration of ART and Peg-IFN-α2b (inclusive of a 4-week ATI period).
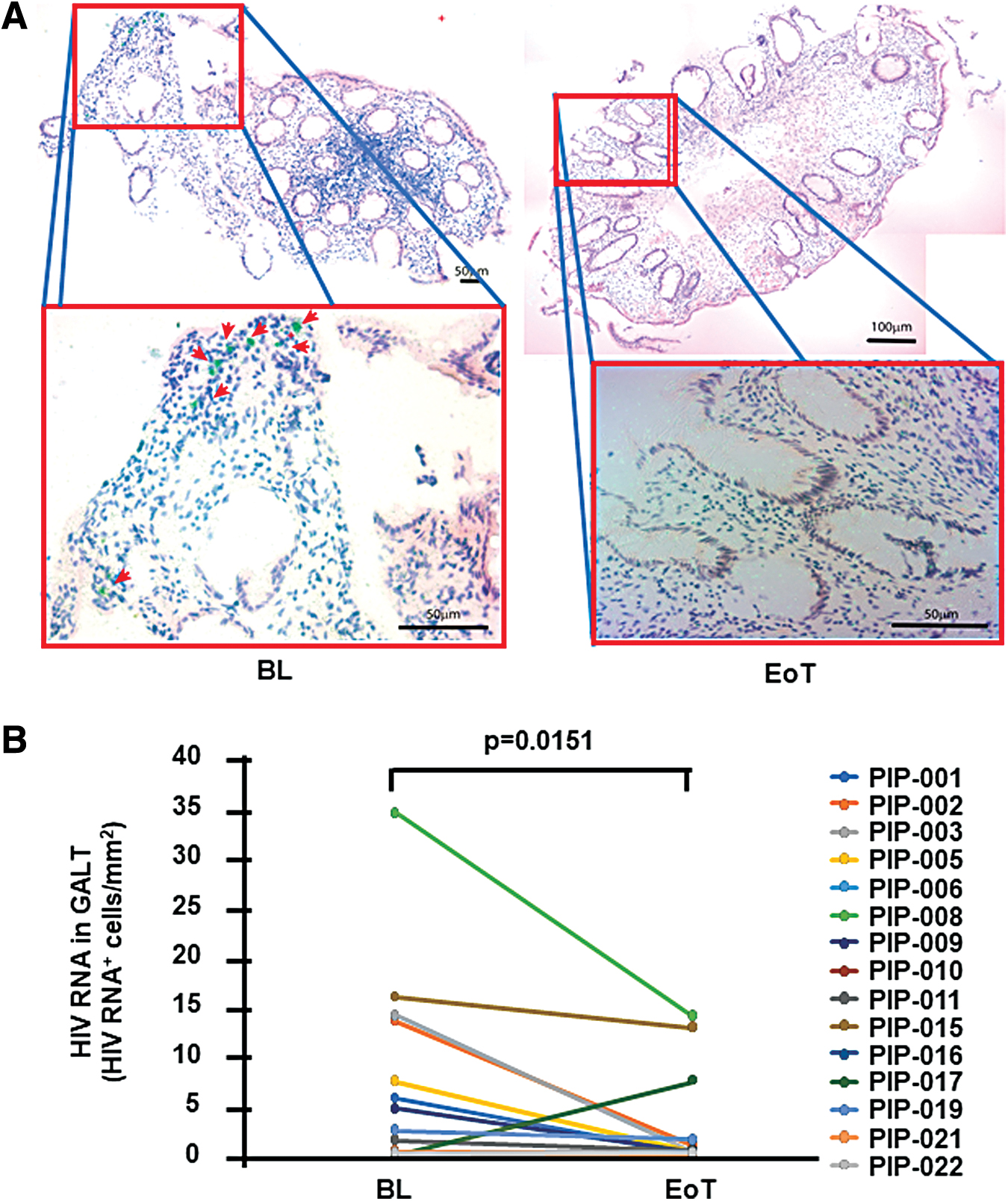
Changes in HIV-1 RNA in GALT after 20 weeks of combined administration of ART and Peg-IFN-α2b (inclusive of a 4-week ATI period).
Immune correlates of the change in HIV-1 RNA+ cells in GALT after 20 weeks of treatment
To assess immune correlates of the change in the number of HIV-1 RNA+ cells in the GALT after 20 weeks of treatment, correlation analysis was performed between viral and host variable Δchanges by EoT. As summarized in Table 4, the Δchange of the number of HIV-1 RNA+ cells was positively associated with the Δchange of the levels of CD4+ T cells (p = .0249, spearman ρ = 0.6162), activated and perforin expressing CD56dimCD16+ NK cells (e.g., CD69+HLA-DR+ % p = .0465, spearman ρ = 0.7143; perforin+ % p = .012, spearman ρ = 0.8333), and activated macrophages (MFI of CD69 p = .0025, spearman ρ = 0.9286).
Correlations of δchange a of HIV RNA in GALT with δchange of Cell Subsets in GALT
δchange was defined as the following difference: end of treatment—BL.
Associations were tested by Spearman correlation test. p Values <.05 were considered significant (unadjusted).
Macrophages were defined as CD3−CD68+CD33+
Discussion
In this pilot clinical trial, we assessed the safety and the effect of 20 weeks of treatment with Peg-IFN-α2b+ART (inclusive of a 4-week ATI) on HIV measures and immune activation in blood and in GALT from chronically HIV+ individuals on long-term suppressive ART. We report for the first time that this treatment can be safe as no SUSAR was observed using the indicated dose of Peg-IFN-α2b, while the only SAE observed was neutropenia which is an expected change of Peg-IFN-α2b immunoregulation. While we only observed a minor change in integrated HIV DNA in peripheral CD4+ T cells, we do provide pilot data to support an increase in markers of activation in blood and in GALT, as well as a reduction in the number of HIV-1 RNA+ cells in GALT in association with changes in innate cell activation. As HIV RNA expressing cells have been described to persist in gut tissues after ART despite long-term undetectable VL in blood, 6 –8 our data are consistent with having achieved a state of greater suppression of HIV expression within tissues as a consequence of Peg-IFN-α2b immunotherapy.
The smaller change in integrated HIV DNA in peripheral CD4+ T cells observed in the current study compared to the change observed with continued Peg-IFN-α2a monotherapy after ATI in our previous clinical trial 18 could be attributed to the continued presence of ART in the current study. It was largely due to an expected greater volume distribution 34 and greater tissue penetration 35,36 of Peg-IFN-α2b compared to Peg-IFN-α2a that we pursued analysis of rectal tissues to test for changes in HIV-RNA in GALT at EoT. The clear reduction in the number of cells that are HIV RNA+ within the total tissue CD4+ T cell population suggests that transcriptionally active cells may be a preferential target for reduction following treatment. This interpretation was not opposed by population based mononuclear cell pellet analysis documenting a reduction in both DNA and RNA despite their lower assay sensitivity to detect a change. Based on the limitations of a one-arm study, future controlled studies will be needed to confirm these observations. The potential to target a reduction of cells expressing viral RNA and antigen would support the benefit of the combined use of Peg-IFN-α and latency reversing agents on ART as currently being tested under the NCT02471430 trial (Peg-IFN-α2a+Panobinostat).
In agreement with previous reports, an increase in the frequency of CD56bright NK cells, as well as an increase in the expression of the inhibitory receptor NKG2A in NK cells 37,38 and in the expression of IFN-responsive molecules (e.g., Tetherin, 39,40 CD169 41 ), was detected in blood following Peg-IFN-α2a immunotherapy evidencing IFN-specific changes after immunotherapy. Consistent with blood changes, increased activation in T and NK cell subsets along with IFN-responsive molecules (i.e., PDL-1) was observed in GALT. These findings are in agreement with reports of modulation by IFN-α resulting in increase in CD56+ NK and CD8+ T cell activation 42,43 and in increase in perforin expression in NK and CD8+ T cells. 44,45
Activation of innate host restriction factors and NK responses have been associated with control of HIV and lysis of autologous HIV-infected CD4+ T cell targets ex vivo 13,46 suggesting that IFN-α immunotherapy may activate similar mechanisms to control HIV infection. Data from our previously reported NCT00594880 clinical trial also showed that following 5 weeks of Peg-IFN-α2a+ART there were increases in innate activation, NK cytotoxicity, and gene expression that were associated with lower plasma VL after ATI and continued Peg-IFN-α2a immunotherapy. 19 We now show that the change in the number of HIV-1 RNA+ cells in GALT following 20 weeks of Peg-IFN-a2b+ART (inclusive of a 4-week ATI period) is positively associated with changes in tissue activation suggesting that the expression of HIV or the immune response against it may induce local activation.
While this pilot study provides safety data for future studies, reported data on anti-HIV effects of immunotherapy have limitations. First, as a pilot study the sample size is limited and lacks a randomized ART-only control group. Second, our study design did not address the effects of immunotherapy on continuous ART as all participants had a 4-week ATI or the effects of Peg-IFN-α2b monotherapy if retained over a longer ATI. Third, no functional anti-HIV responses by NK or T cells were assessed in association with an increase in activation markers. To advance the pilot data presented here and due to the safety of Peg-IFN-α2b when added to ART as demonstrated in this pilot study, a follow-up randomized clinical trial (NCT02227277) in 54 persons is underway to establish the antiviral effects of Peg-IFN-α2b.
Conclusion
Our pilot study supports that 20 week immunotherapy with combined Peg-IFN-α2b+ART (inclusive of a 4-week ATI) is safe, results in increase in blood and GALT immune activation, and can mediate a reduction in the number of cells expressing HIV-1 RNA in GALT in association with changes in innate cell activation.
Footnotes
Acknowledgments
The authors thank the study participants and their providers, as well as Agnieszka Mackiewicz, Jocelin Joseph, Guobin Kang, Kenneth M. Lynn, Angela Kapalko, Linden Lalley-Chareczko, Faten Aberra, Patricia Tsao, and Eline Luning Prak, for study support.
Authors' Contributions
A.P., N.C., Q.L., M.A.-M., B.N.R., and M.F. performed experimental work. K.M., J.R.K., and P.T. selected and recruited patients. E.P. performed the statistical analysis. B.J.H. and D.J.H. provided the study drug (Peg-IFN-α2b). E.P., L.A., N.C., Q.L., M.A.-M., B.J.H., D.J.H., and L.J.M. designed the study and evaluated the results. E.P., L.A., M.A.-M., and L.J.M. wrote the article.
Author Disclosure Statement
The authors declare that they do not have a commercial or other association that might pose a conflict of interest.
Funding Information
This work was supported by the following grants to LJM: U01AI110434, UM1AI126620, Robert I. Jacobs Fund of the Philadelphia Foundation, and the Herbert Kean, MD, Family Professorship. Additional support was provided by the Penn Center for AIDS Research [AI045008] and The Wistar Cancer Center Support Grant [CA10815]. MA-M is supported by the National Institutes of Health (NIH) grants [grant numbers: R01 DK123733, R01 AG062383, R01NS117458, R21 AI143385, R21 AI129636, and R21 NS106970]. NC is supported by the Research Scholar Career Awards of the Fonds de Recherche du Québec (FRQ-S) [grant number 253292].
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.