
Abstract
The persistence of HIV in the spleen, despite combination antiretroviral therapy, is not well understood. Sustained immune dysregulation and delayed immune recovery, in addition to immune cell exhaustion, may contribute to persistence of infection in the spleen. Eliminating HIV from this secondary lymphoid organ will require a thorough understanding of antiretroviral (ARV) pharmacology in the spleen, which has been minimally investigated. Low ARV exposure within the spleen may hinder the achievement of a functional or sterilizing cure if cells are not protected from HIV infection. In this study, we provide an overview of the anatomy and physiology of the spleen, review the evidence of the spleen as a site for persistence of HIV, discuss the consequences of persistence of HIV in the spleen, address challenges to eradicating HIV in the spleen, and examine opportunities for future curative efforts.
Introduction
Since the dawn of combination antiretroviral therapy (cART) in 1996, the life expectancy for those living with HIV has increased dramatically to approach near that of non-HIV-infected individuals. 1 Following interruption of viral-suppressive therapy, however, rebound viremia occurs after ∼2–3 weeks. 2 –4 It has been estimated that the half-life of HIV from reservoirs is 44 months, indicating that it would take nearly 73 years of continuous suppressive therapy for full eradication from the body. 5,6 The persistence of HIV in the presence of effective cART remains a significant barrier to a cure.
The latent reservoir is established early in infection and comprises memory and follicular helper T cells. 7 While the hypothesis of active replication occurring in lymphoid tissue remains controversial, 8 –10 the persistence of HIV in the lymphoid tissue reservoir has been associated with suboptimal antiretroviral (ARV) concentrations. 11,12 The spleen, a lymphoid organ, is one of the first locations to mount the innate immune response against viral infections, as well as to perform foreign antigen presentation. Recently, the detection of cell-associated viral RNA in spleen tissue may indicate either residual replication or HIV release from stable reservoirs. 13 ARV penetration has been relatively understudied in the spleen.
Novel clinical pharmacology tools may provide valuable insight into the persistence of HIV in this lymphoid reservoir with current cART and future eradication interventions. Our group has reviewed the pharmacological challenges and opportunities related to the persistence of HIV in the gut-associated lymphoid tissues, 14 yet data regarding the persistence of HIV in the spleen have yet to be reviewed. In this study, we review the anatomy and physiology of the spleen, the evidence of HIV persistence and its consequences, challenges to eradicating HIV in the spleen, and the role of the spleen in future HIV cure therapy.
Anatomy and Physiology of the Spleen
There are two main compartments in the spleen: the red pulp and the white pulp. These two functional units have distinct vascular architecture and cellular compositions. While we briefly review the anatomy and physiology of the spleen, more in-depth discussions of splenic structure have been previously published. 15 –18 To assist readers with the following section, Figure 1 illustrates the microcirculation of the spleen.
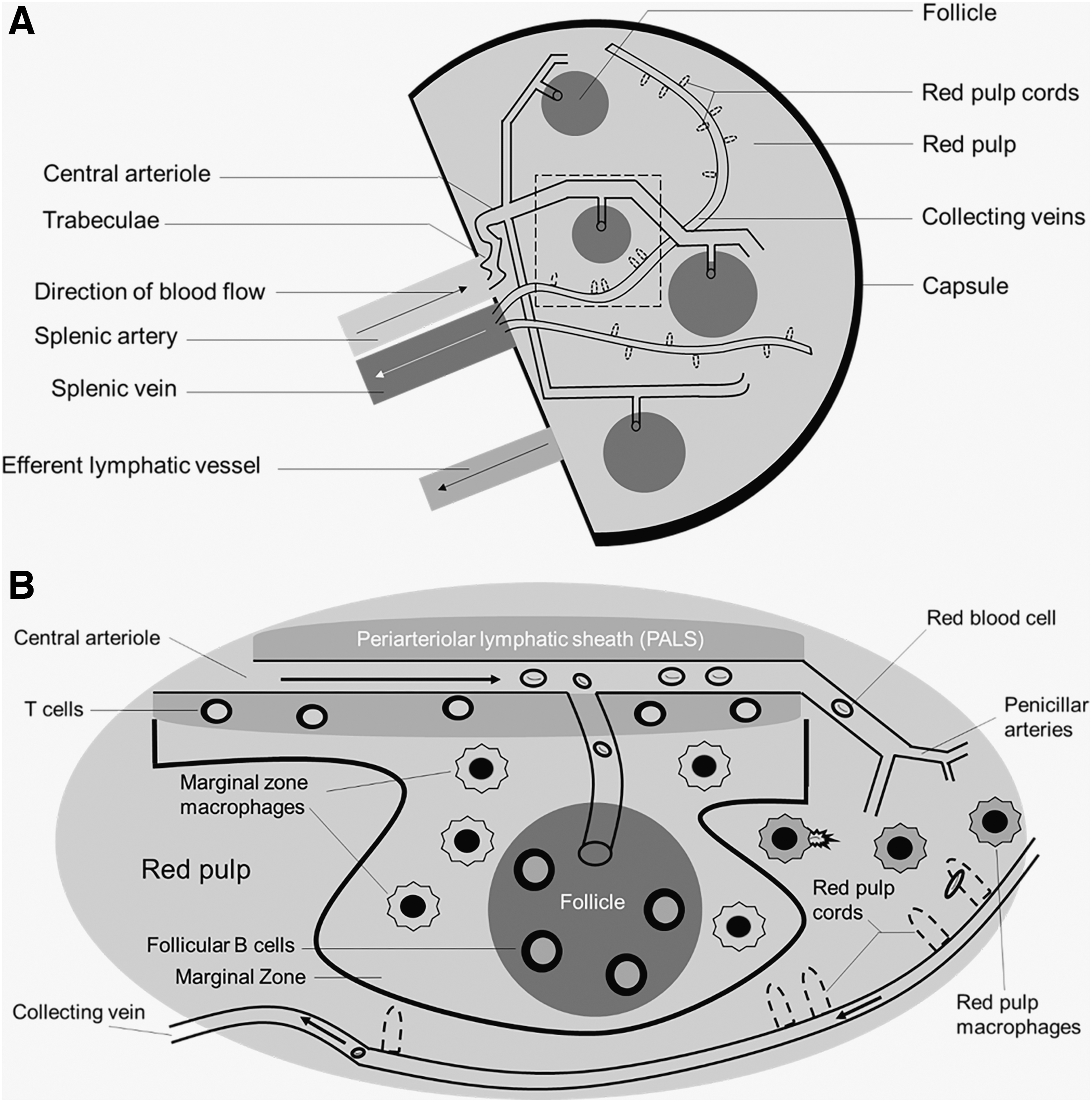
Physiologic overview of spleen anatomy and physiology. The splenic artery comes from the celiac trunk of the descending aorta to provide oxygenated blood to the spleen.
The spleen comprises red and white pulp. The red pulp has three main functions: filtering blood, recycling iron (following erythrophagocytosis), and storage of platelets. 16 Splenic macrophages are responsible for the recycling of iron and compete with bacteria for the utilization of iron for survival. Therefore, the red pulp also serves as a defense against pathogens.
Comprising nearly 25% of the splenic tissue, the white pulp contains numerous immunological cells, including—but not limited to—macrophages, dendritic cells, and plasma B cells. 16 T cells are mainly located in the periarterial lymphatic sheaths (PALS, described below). B cells are primarily located in the follicles and are responsible for the production of antibodies following opsonization of foreign antigens (most notably encapsulated bacteria). The marginal zone of the white pulp contains dendritic cells and macrophages responsible for antigen presentation.
Blood flow to the spleen ranges from 170 to 330 mL/min in humans, which is ∼5% of the total cardiac output. 19 In nonhuman primates (NHPs, a common model for HIV treatment, prevention, and cure), blood flow to the spleen is 21 mL/min, close to 2% of cardiac output, although the spleen weighs significantly less (8 g vs. 180 g in humans). 20
Branching off the abdominal aorta and into the celiac artery, the splenic artery provides oxygenated blood to the spleen. As the splenic artery enters the hilus (fissure) of the spleen, the splenic artery begins to branch off and direct into the fibrous trabeculae. These central arteries are surrounded by lymphatic tissue (PALS) that form the characteristic splenic follicles, comprising the white pulp. 16,17 The central arteries further branch to form penicillar arteries, which enter the red pulp of the spleen. Greater than 90% of the blood flow to the spleen goes to the white pulp, thereby circumventing the red pulp. 21
There are two morphological components of the red pulp that distinguish it from the white pulp: splenic sinuses (sinusoids) and splenic cords (Cords of Billroth). The splenic sinuses are capillaries lined with discontinuous endothelium as well as slits to perform the sifting of erythrocytes in the spleen. These sinuses eventually form veins (venous sinuses). The splenic cords are located within the red pulp and comprise reticular fibers and macrophages. These fibers are composed of collagenous fibers that may be responsible for splenic contraction and are bound by the extracellular matrix and fibroblasts.
From a pharmacologic perspective, the morphology of the splenic sinuses allows the permeation of not only large endogenous molecules, such as albumin, but also large exogenous molecules, such as monoclonal antibodies. For instance, it has been previously demonstrated that rituximab, a monoclonal antibody utilized in the treatment of various cancers and rheumatologically based conditions, distributes into the spleen. 22 –25
Finally, blood circulating into the red pulp collects in the venous sinuses that enter the trabeculae of the spleen, through the trabecular veins, and ultimately into the splenic vein. The splenic vein then drains into the hepatic portal vein and into the liver. Therefore, there is both anatomical and physiological communication between the spleen and the liver. Similar to lymph nodes, the spleen contains an efferent lymph vessel that drains into the lymphatic system; unlike the lymph nodes, the spleen does not contain any afferent lymph vessels. 15,16
HIV Persistence in the Spleen
Although cART can decrease HIV in the plasma to undetectable concentrations, it cannot completely clear HIV infection from the body. Since <2% of total body lymphocytes are in the blood, it is imperative to examine HIV in anatomical sites. 26 The spleen is the largest secondary lymphoid organ and contains approximately one-quarter of the body's lymphocytes, which combines the functionality of the innate and adaptive immune responses. Yet, the spleen is understudied. In 1997, Reinhart et al. noted in eight rhesus macaques infected with simian immunodeficiency virus (SIV) deltaB670 that the spleen contains 2 × 108–10 SIV virions [measured by in situ hybridization (ISH) silver staining] and that the viral RNA was observed in the PALS and cells in the red pulp. 27
Technological advances in quantifying HIV reservoirs have aided in understanding virology and the spatial distribution of virus. These advances include RNAScope and DNAScope, an ISH approach that utilizes “double-Z” target probes to bind in a contiguous manner to complementary RNA or DNA sequences, respectively. 28 In these scenarios, the probability of two probes adjacently nonspecifically binding is low, thereby providing minimal background noise. Deleage et al. observed that the B cell follicles of lymph nodes were anatomical compartments for viral persistence, even after 26 weeks of viral suppressive cART to rhesus macaques. 29 The flexibility of this approach allowed for the same authors to demonstrate persistence of SIV RNA on follicular dendritic cells and within the B cell follicles in the spleens of chronically infected rhesus macaques, similar to what had been seen in the lymph nodes. 28
To further provide evidence of viral persistence in the spleen, Estes et al. examined 27 rhesus macaques with distinct simian AIDS viruses (nine total spleens) using the RNAScope and DNAScope procedures and demonstrated that even in chronically treated macaques, SHIV and SIV viral RNA persist in the spleen as >99% of the SIV RNA+ cells were located in lymphoid tissues. 13 Certainly, the presence of virus in the tissue does not necessarily indicate ongoing viral replication, but the numbers of virions seen in these studies indicate the sources of virus that may reemerge following treatment interruption.
There is evidence from postmortem human spleens that may further indicate persistence of virus. For example, Nolan et al. examined the spleens from five participants on suppressive cART utilizing droplet digital PCR and single-genome sequencing. 30 In this analysis, maximum likelihood phylogenies indicated that both the env and nef viral genes showed viral diversity in the spleen and that because both DNA and RNA sequences were detected in two out of five patients, this suggested active virus replication. 30 However, this is not without controversy.
Bozzi et al. conducted phylogenetic analyses to investigate HIV populations in infected tissues, including the spleen. 31 The authors did not find evidence of ongoing replication or compartmentalization of HIV within the spleen. but did detect clonal expansion of infected cells that predated cART, concluding that the extent of clonal expansion of cells that carry infectious proviruses should be the focus of curative efforts. 31 Therefore, while there may potentially be evidence of actively replicating virus, this is still controversial.
Taken together, there is evidence that suggests HIV persists in the spleen. While much of the conclusions are extrapolated from lymph nodes because of their ability to be extracted for further study, morphological similarities between the two organs might indicate similar processes. Furthermore, splenic data are derived from NHP preclinical models, which can recapitulate the infection process. These data can help inform immune dynamics and viral persistence in the human spleen.
Consequences of Persistence of HIV in the Spleen
Immune dysregulation
Like other pathogenic disease states, the spleen plays an important role in initial antigen presentation and subsequent antibody development against HIV. A key component of the immune cascade is the follicular T helper (Tfh) cell, which is responsible for the development of the germinal center, the site of somatic hypermutation and plasma B cells. There are conflicting reports related to the frequency of Tfh in the blood. Earlier reports indicated that during the acute and chronic phase of HIV infection, frequencies are increased. 32 Conversely, Boswell et al. indicated decreased frequencies of Tfh cells. 33 Within lymphoid tissues, Cubas et al. noted that the triggering of programmed cell death ligand 1 on Tfh cells within the lymph nodes diminishes the B cell responses during HIV infection and potentially lessens control of HIV infection, owing, in part, to the reduction in interleukin (IL)-21 secretion. 34
Rodrigues et al. noted in a rhesus macaque model infected with Leishmania infantum, the abortive differentiation of splenic Tfh cells, relating to the failure of these cells to express PD-1. 35 Ultimately, this led to altered B cell differentiation and reduced production of antigen-specific immunoglobulins. In fact, immunohistochemical (IHC) analysis has indicated the atrophy and disruption of the germinal center during visceral leishmaniasis. In the context of HIV or SIV, Moukambi et al. utilized rhesus macaques of Indian origin infected with SIVmac251 and showed that—in the acute phase of infection—splenic Tfh cells decrease early after infection and remodeling of normal splenic architecture occurs. 32 Accompanying the loss of Tfh cells were the loss of memory B cell subsets and lower titers of immunoglobulins against SIV. 32
Collagen deposition
Mechanisms underlying collagen deposition and subsequent depletion of CD4+ T cells are related to the inflammation and chronic immune activation of HIV infection. Fibrosis in preclinical animal models has indicated the mediator role of transforming growth factor (TGF)-β1 on regulatory T cells in the lymphatic tissues of sooty mangabeys and rhesus macaques. 36 These regulatory T cells expressing TGF-β1 were directly linked with collagen type 1 expression in fibroblasts and may lend credence to the notion that suppressive cART does not completely recover T cells or reverse collagen deposition in the lymphatic tissues. 36 –38
Due to infeasibility of serial sampling, this has not been holistically demonstrated in the spleen, although red pulp enlargement and lymph follicle atrophy were noted in the spleens from 34 splenectomized patients secondary to hepatosplenic schistosomiasis. 39 Given the morphological and architectural similarities between lymph nodes and the spleen, similar processes with HIV would be expected, although this has not been confirmed.
In the context of immune system homeostasis, the secondary lymphoid tissues—specifically the spleen—play a vital role. The architecture of these lymphoid organs is ideal to mount an immune response following the introduction of an antigen. For instance, the T cell zone (TZ) is the primary site of HIV activation as the majority of CD4+ T cells reside here. 40 It has been previously demonstrated that the TZ decreases in size during the progression of HIV infection. 41 Schacker et al. 41 prospectively examined histological sections of inguinal lymph nodes from HIV-positive patients on suppressive cART and noted depletion of the CD4+ T cell population in all stages of HIV infection secondary to damage from collagen. The area of tissue occupied with collagen was significantly negatively correlated (r = −0.55, p = .0008) with tissue-naive CD4+ T cells. 41
This was further supported by a separate, follow-up study by the same group, indicating the deposition of collagen was associated with the depletion and impaired reconstitution of CD4+ T cells. 42 In 2008, Estes et al. expanded these findings to the gut-associated lymphoid tissue (GALT), demonstrating a significant, inverse relationship (r = −0.60; p = .004) between the percent area occupied by collagen in the TZ of the Peyer patches and the CD4+ cell population. 43 Furthermore, treatment in early infection can increase a specific subtype of T cell—CD4+ central memory T cell—in the Peyer patches, even after 6 months of suppressive cART. 43
In the inguinal lymph nodes, Zeng et al. noted that collagen damages the fibroblastic reticular cell (FRC) network, contributing to the loss of naive T cells, mediated by decreased IL-7 production and presentation to the FRC, even following suppressive cART administration. 44 Tien et al. found that plasma fibrinogen and C-reactive protein were independent predictors of 5-year mortality risk in HIV-infected patients. 45 This increased inflammatory state secondary to HIV infection causes damage to the lymphoid tissue. Given these results, adjunct antifibrotic and/or anti-inflammatory therapy have been speculated and trialed with mixed results. 46,47
Inflammatory sequelae
Although cART has allowed for the reconstitution of peripheral CD4+ T cells, HIV-infected adults are at greater risk of non-AIDS-related morbidity, including cardiovascular disease, osteoporosis, and liver disease. 48 One reason for the increased risk of these morbidities is inflammation. A higher frequency of inflammatory monocytes, cytokines, and hypercoagulability biomarkers has been noted, even with consistent, viral suppressive cART. 45,49 –52
Another key sequelae of the chronic inflammatory state is the activation of macrophages and monocytes. Table 1 displays the four main macrophage subsets in the spleen. The activation is accompanied by increased turnover and frequency of circulating monocytes, which may be regulated by the marginal zone macrophages. 53 –56
Macrophage Subsets in the Spleen
F4/80, epidermal growth factor-like module-containing mucin-like hormone receptor-like 1; VCAM-1, vascular cell adhesion molecule 1; HO-1, heme oxygenase 1; SIGN-R1, specific intracellular adhesion molecule-grabbing nonintegrin receptor 1; MARCO, macrophage receptor with collagenous structure; LXRα, liver X receptor-alpha; Tim4, T cell membrane protein 4.
This local, splenic activation may explain the findings seen by IHC. In 72 spleens from postmortem AIDS patients, histological evidence suggested drastic changes ranging from follicular hyperplasia to follicular atrophy, which had been directly attributable to HIV infection rather than concomitant opportunistic infections. 57 Furthermore, the inflammation resulted in splenomegaly (median splenic weight was 280 g in those without secondary pathologic lesions). 57 It had been noted from the spleens of these patients that progressive HIV-impaired activation of macrophages in the spleen secondary to follicular morphological changes. 57,58
This was further explored by Williams et al. using an accelerated SIV-infected NHP model. 59 In this study, uninfected, acutely infected, infected, but asymptomatic, and chronically infected untreated NHP spleens were immunologically stained for CD163+CD68+ macrophages. Two important findings were demonstrated. First, the immunology of marginal zone macrophages changed during the acute and chronic phases of infection. In the acute phase, universal upregulation of CD163 was noted, with accompanying loss of CD68 throughout the organ. In the chronic phase, CD163+CD68+ macrophages entered the germinal center to compensate for the acute phase loss of CD68. 59 Second, the authors noted lymphoid depletion, illustrated by hyposplenism, white pulp depletion, and germinal center burnout related to T cell depletion from infection. 59
Taken together, the pathogenesis of HIV infection directly affects the spleen early in infection and the physical presence of virus in the lymphoid organ causes morphological changes that impair responses to other immunological insults. Furthermore, these morphological changes continue to occur in the chronic phase of infection, although the modulatory effect of cART—if present—has yet to be determined.
Challenges to Eradicating HIV in the Spleen
Clonal expansion
Clonal expansion describes the process of T and B lymphocytes interacting with an antigen or antigen-presenting cell and subsequently generating clones through mitosis against a specific antigen. It has been shown that clonal expansion of HIV-infected cells can persist for over 11 years, even on suppressive cART. 60 Indeed, the identical HIV DNA sequences of infected cells in peripheral blood and tissues lend credence to cellular proliferation being the contributor to the persistence of HIV. 61 –63
Lee et al. 64 performed cross-sectional genetic analyses on RNA sequences from T cell subsets in blood, lymph nodes, and gut tissues from patients on suppressive cART for 3–17.8 years and demonstrated two important findings. First, the number of infected memory T cells decreases with cART initiation. Second, effector memory T cells from the lymph nodes can produce infectious virus through clonal expansion. This maintains HIV persistence and offsets the decay of HIV-infected cells. 64 The effector memory T cells from the lymph node contained provirus, which was genetically identical to those in plasma. 64
Similarly, von Stockenstrom et al. isolated intracellular HIV-1 genomes from CD4+ T cell subsets of peripheral blood, GALT, and lymph node tissues, from patients receiving suppressive cART, separated by 7–9 months. 65 The authors noted that DNA integrant frequencies were stable over time and that clonally expanded populations were found in effector memory T cells isolated from lymph nodes. 65 While these data are derived from lymphoid tissues, data for the spleen remain sparse, and although morphological similarities between lymph nodes and spleen may permit extrapolation, it is important to be able to examine the spleen directly.
Bozzi et al., utilizing phylogenetic analyses of infected postmortem tissues of treated HIV-infected individuals, did not find evidence of ongoing replication in tissues. 31 In the two spleen samples examined (cART duration: 8.0 and 22.7 years, respectively), there were identical HIV sequences determined by average pair wise differences (0.26% and 1.7%, respectively), compared to the variants sequenced from peripheral blood mononuclear cells (PBMCs) after years of cART. 31 Although this is a sample size of 2, these data do point to the spleen potentially containing clones of virus present before starting cART, representing another challenge to fully eradicate HIV.
ARV penetration and modulation
Although the spleen plays a crucial role in immunoregulation, the study of the spleen as a reservoir for HIV is challenging. The spleen is the first place of antigen presentation, but the in-depth study of the organ is limited to postmortem individuals or situations in which the spleen must be removed (e.g., splenectomy secondary to ruptured spleen following motor vehicle accident). However, there have been a few studies examining the distribution of ARVs into the spleen of animal models.
Di Mascio et al. administered 5 mg/kg intravenous bolus doses of a radiolabeled derivative of tenofovir to Sprague-Dawley rats and detailed the biodistribution of the compound into the spleens after 120 min. 66 The authors noted that compared to plasma, splenic concentrations were ∼2-fold lower. 66 Furthermore, positron emission tomography imaging indicated a slower accumulation and clearance of the drug from the spleen compared to plasma. 66 Similarly, Lee et al. described the biodistribution of a one-time administration of radiolabeled tenofovir disoproxil fumarate to two beagle dogs, which were sacrificed at 24 h. 67 The percent injected dose per gram of tissue in the spleen observed by Lee et al. (<0.01%) was similar to Di Mascio et al. (<0.05%). 66,67
Devanathan et al. demonstrated that NHPs achieved higher splenic penetration than humanized mouse models or humans alongside higher molar percentages of the active intracellular metabolites of nucleoside reverse transcriptase inhibitors (NRTIs), indicating a higher rate of active metabolite conversion. 68 However, for the NRTIs and metabolites in humans, inhibitory quotient values did not exceed 1, which may result in lack of virologic control. 68
In a separate analysis, it was noted that between plasma and tissue concentrations, the NRTIs emtricitabine and tenofovir had positive linear relationships and raltegravir had inconsistent relationships in the secondary lymphoid organs, highlighting the heterogeneity of ARV penetration into tissues. 69 Fletcher et al. examined ARV concentrations in PBMCs and mononuclear cells from lymph nodes, ileum, and rectum, and noted that lower drug concentrations correlated with slower HIV decay rate in the tissues. 12
All taken together, this would lead one to believe that it is the lack of ARV tissue penetration that contributes to the persistence of HIV. However, this conclusion remains controversial. 31 Should this be the case, drug resistance would likely occur during the setting of subtherapeutic ARV concentrations, as seen in the plasma of patients with poor drug adherence, but this is not observed. 70 Instead, it has been previously noted that the concentrations in tissue reservoirs may be low enough as to not permit the development of resistance, but allows wild-type virus to continue to replicate, thereby outcompeting resistant variants. 71
Optimizing cART in the spleen is difficult for many reasons. First, as mentioned previously, there are limited data of ARV exposure in the spleen. In addition, drug transporters and metabolizing enzymes may modulate the disposition of ARVs in the spleen. While a comprehensive discussion of drug transporters is beyond the scope of this review, it is well known that ARVs interact with the ATP-binding cassette membrane-associated efflux transporters and solute carrier uptake transporters. 72 The expression of these transporters can be altered endogenously or exogenously, thereby affecting ARV concentrations. In preclinical species, quantifiable Mrp4, Bcrp, and Ent1 concentrations have been demonstrated, although these did not significantly correlate with ARV concentrations . 68
Drug-metabolizing enzymes (DMEs) are highly responsible for the disposition of ARVs. Macrophages derived from bone marrow and blood express cytochrome P450 (CYP) 1A1, 2A6/7, 2D6, 2E1 and 3A4 mRNA, which are responsible for the biotransformation of numerous ARV classes, such as protease inhibitors and non-NRTIs. 73 –75 Furthermore, gene expression of DMEs has been shown in human PBMCs. 76 While the DMEs in the spleen have yet to be fully characterized, it is possible that due to the wealth of lymphocytes in the spleen, DMEs may play a role in the intracellular disposition and metabolism of small molecules, including ARVs.
The role of inflammation on potential modulation of ARV disposition into reservoir tissues has yet to be elucidated. In the context of antimicrobials, pathophysiological changes in critical illness (alterations in protein binding, pH, etc.) dictated by increases in acute phase proteins affect distribution into the active site of infection, resulting in concentrations below the minimum inhibitory concentration. 77 Although it has been previously demonstrated that the protein binding potential was lower than that in plasma, these values did not differ between NHPs infected with recombinant SHIV and uninfected controls. 68 Taken together, there may potentially be a role of inflammation or inflammatory sequelae (e.g., collagen deposition as above) associated with HIV infection that modulates ARV penetration and leads to inadequate penetration in lymphoid tissues, but this needs further study.
Future Approaches to Understanding ARV Tissue Distribution
Mass spectrometric imaging
Evaluations of ARVs in tissues have conventionally utilized liquid chromatography-mass spectrometry (LC-MS) of tissue homogenates, which has been regarded as the gold standard of tissue quantification. 78 However, a limitation to this analysis is that tissue homogenates lose spatial and distributional configurations within the tissue due to total consumption of sample. This would lead to significant overestimation of concentrations at the site of action, and misleading conclusions regarding exposure-response relationships. 79 Our group has demonstrated that distribution of ARVs is not uniform across the tissue reservoirs. 80,81
These data were generated through a novel mass spectrometry imaging method known as infrared matrix-assisted laser desorption electrospray ionization (IR-MALDESI). 82,83 The process and workflow of IR-MALDESI have been described previously. 83 IR-MALDESI combines the technical specifications of traditional LC-MS/MS methods, with heat-map style imaging of variable drug concentrations across a snap-frozen, unaltered tissue section. In addition, IR-MALDESI uses ice as a matrix, which avoids organic matrix peak interference, allows collection of multiple masses within the tissue, reduces voxel-to-voxel variability, and increases sensitivity. 83,84
The heat-map style imaging data generated from MALDESI require tissue slices for analyses. Adjacent spleen tissue slices can be sectioned for IHC and ISH analyses and registered with IR-MALDESI images using numerical programming language (e.g., MATLAB) algorithms. Correlation analyses can be assessed qualitatively by visual comparisons and quantitatively through image analysis of the total percentage of the image occupied by a specific ARV (IR-MALDESI), CD4+ cells (IHC), and vRNA/vDNA using RNAScope/DNAScope (ISH). This combinatorial approach allows for the visualization of ARVs, HIV RNA/DNA, and immunological cells, possibly demonstrating that merely achieving quantifiable splenic ARV concentrations may not be sufficient for effective viral control, as suggested previously. 11,12
Furthermore, the quantitative data abstracted from IR-MALDESI can lend credence to the ARV concentrations needed within the tissue compartment to suppress virus. Thompson et al. assessed the distribution of ARVs in gut tissues across three species and measured the distribution relative to CD3+ T cells and the expression of HIV and SHIV RNA. 81 Across the three species, it was demonstrated that 40%–60% of the CD3+ T cells remained unexposed to ARVs, indicating that ARV exposure in some areas where HIV target cells reside may be limited. 81 These analyses should be similarly performed in the spleen and other tissue reservoirs (lymph nodes, genital tract, etc.) to elucidate the relationship between potential suboptimal ARV exposure and HIV persistence, and represent ongoing work by our group.
Opportunities for HIV Eradication
Small molecules
The concept of “kick and kill” strategies describes latency-reversing agents (LRAs) combined with immunotherapeutic intervention, as described below. 85 Numerous LRA classes have been investigated, but the most widely explored are the histone deacetylase inhibitors (HDACi). Deacetylation of histones leads to compacted chromatin, hindering gene expression. Inhibition allows for the unpacking of the chromatin and subsequent immune recognition of the latent cells. 86 Examples of HDACi are vorinostat, panobinostat, and romidepsin.
Tissue concentrations of HDACi are scarce. Recently, Perrin et al. dosed three rats intravenously with 10 mg/kg panobinostat and concentrations in tissues were quantified, and the mean concentration in the spleen was 2.9 μM. 87 Compared to EC50 values of HIV-1 wild-type and mutant strains determined by Norton et al. (1.6–12.86 nM), 88 the concentrations are over 200-fold higher.
Regarding romidepsin, it was noted that when cutaneous burn-injured and control mice were dosed with 5 mg/kg intraperitoneal injections twice daily, there was an increase in histone acetylation. 89 Within the spleen tissue, there was a 17% increase in histone acetylation (a biomarker of romidepsin's effect). This suggests splenic bioavailability of romidepsin, although drug concentrations are unknown. Since LRAs are used in combination with immunotherapeutic agents, a discussion of these follows.
Monoclonal antibody therapy
Monoclonal antibodies represent a novel approach for the treatment and potential eradication of HIV. Most recently, the sole monoclonal antibody approved in the United States for the treatment of HIV infection is the postattachment inhibitor ibalizumab, which targets the extracellular CD4 domain. 90 –94 Due to their designed specificity for a particular antigen, monoclonal antibodies have been utilized for decades in other immune-related diseases, such as cancer, rheumatoid arthritis, and psoriasis. Monoclonal antibodies greatly differ from conventional small molecules as they are highly polar and have molecular weights of ∼150 kDa. 95
As a result of these different physicochemical properties, distribution of antibodies into tissues occurs primarily through convection. 96 –98 Similar to other proteins, monoclonal antibodies experience a sieving process, which is mathematically represented by reflective coefficients. Most tissues have capillary beds comprising reflective coefficients of 0.95–0.98, indicating virtual impermeability to plasma proteins. In contrast, spleen sinusoidal capillaries contain reflective coefficients of 0.85, lending credence to the notion that plasma proteins circulate into the spleen. 97,99 Indeed, the spleen has been shown to be a preferential site of accumulation of therapeutic antibodies.
FCγ receptors in neutrophils, monocytes, and macrophages are responsible for the internalization and destruction of antibodies. 100 However, binding to FcRn (also known as the Brambell receptor) allows for the recycling of the antibody into the plasma upon acidification. 100 It has been demonstrated that the spleen expresses FcRn receptors. 101 Indeed, in FcRn knockout mice, immunoglobulin accumulation occurred in the liver and spleen, both organs that contain the aforementioned sinusoidal capillary system. 102 A well-documented example is the monoclonal antibody rituximab, which targets splenic antigens (i.e., CD20 antigens on B cell splenocytes), and we direct readers to the numerous references detailing the pathway of rituximab as it relates to the spleen. 22 –25
While therapeutic antibodies were proposed initially as a curative strategy for HIV, suboptimal efficacy in preclinical animal models and early unsuccessful clinical trials in humans has tempered enthusiasm. Currently, broadly neutralizing antibodies may potentially regenerate the interest in large protein therapeutics to treat and prevent HIV infection. While a review of antibody therapeutics is beyond the scope of this article, interested readers are directed to a review by Gruell and Klein. 103
Adoptive T cell therapy
Another novel approach to HIV therapy is adoptive cell therapy, particularly the engineering of T cells with chimeric antigen receptors (CARs) that are directed toward the HIV envelope. 104 Currently, CAR T cell therapy is explored and approved for cancer immunotherapies, including B cell leukemia. In the 1980s, early attempts of CARs utilized soluble CD4 molecules to prevent HIV infection by blocking the interaction between CD4 and the envelope; however, resistance and a short half-life of the soluble CD4 hampered their efficacy. 105,106
Advances have enhanced technical features of T cell therapies in the context of hematological malignancies. One improved design has been to introduce antibodies against viral antigens versus the extracellular regions of CD4 cells, allowing for specific binding to the viral antigen and limited off-target effects, although differences in affinities of the antibody component are being evaluated. 107,108
Despite the advancements in T cell therapies, the pharmacokinetics remain relatively unknown. Biodistribution studies of T cells in mice prepared from splenocytes or tumors concentrated primarily in the spleen and lung after 20–24 h. 109,110 Early development of mathematical models relied on incomplete biodistribution data or a lack of experimental data. 111 Khot et al. utilized chromium-51-labeled T cells from splenocytes that were expanded using anti-CD3, anti-CD28, and IL-2, and administered to a melanoma mouse model. 112 After noting that the T cells are rapidly cleared from the blood following administration, >90% of the T cells distributed in the spleen, liver, lungs, kidneys, bone, and lymph nodes. 112
Other studies utilized chromium-labeled lymphocytes and noted high body surface counts of lymphocytes in the spleen and liver, both of which have an intricate reticuloendothelial system as aforementioned. 113 More sophisticated mathematical modeling approaches will be warranted to characterize the biodistribution of CAR T cell therapies that utilize antibodies in the future.
Conclusion
In summary, the spleen is an understudied organ in the context of HIV, and the gaps in knowledge range from ARV concentration analyses to distributional data examining ARVs in relationship to immunological cells and virus. New pharmacologic methods can help bridge those knowledge gaps. These data will help inform HIV treatment and cure clinical trials that would potentially harness immunologically related therapies with small molecules.
Footnotes
Authors' Contributions
A.S.D. and A.D.M.K. wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors are grateful for the National Institutes of Health Grant R01AI111891 (to A.D.M.K.). A.S.D. is supported by the National Institute of General Medical Sciences of the National Institutes of Health under Award Number T32GM086330.