
Abstract
In Sub-Saharan Africa, young women 15–24 years of age account for nearly 30% of all new HIV infections, however, biological and epidemiological factors underlying this disproportionate infection rate are unclear. In this study, we assessed biological contributors of SIV/HIV susceptibility in the female genital tract (FGT) using adolescent (n = 9) and adult (n = 10) pigtail macaques (PTMs) with weekly low-dose intravaginal challenges of SIV. Immunological variables were captured in vaginal tissue of PTMs by flow cytometry and cytokine assays. Vaginal biopsies were profiled by proteomic analysis. The vaginal microbiome was assessed by 16S rRNA sequencing. We were powered to detect a 2.2-fold increase in infection rates between age groups, however, we identified no significant differences in susceptibility. This model cannot capture epidemiological factors or may not best represent biological differences of HIV susceptibility. No immune cell subsets measured were significantly different between groups. Inflammatory marker MCP-1 was significantly higher (adj p = .02), and sCD40L trended higher (adj p = .06) in vaginal cytobrushes of adults. Proteomic analysis of vaginal biopsies showed no significant (adj p < .05) protein or pathway differences between groups. Vaginal microbiomes were not significantly different between groups. No differences were observed between age groups in this PTM model, however, these animals may not reflect biological factors contributing to HIV risk such as those found in their human counterparts. This model is therefore not appropriate to explore human adolescent differences in HIV risk. Young women remain a key population at risk for HIV infection, and there is still a need for comprehensive assessment and intervention strategies for epidemic control of this uniquely vulnerable population.
Introduction
According to the World Health Organization, a disquieting estimate of more than 30% of new human immunodeficiency virus (HIV) infections globally are in young adults and adolescents (ages 15–24). 1 In areas such as sub-Saharan Africa, which has 70% of new global HIV infections, young women have incidence rates up to four-times higher than their male peers, 1 and acquire HIV infection at least 5–7 years earlier. 1,2 In high-burden areas, new infections are on the rise with young women (< 20 years) rising from 13% in 2007 to as high as 22% in 2013. 3
The ASPIRE and The Ring trials evaluating the dapivirine vaginal ring for HIV protection failed to demonstrate a benefit in women under 21 (p = .02), but it is unclear whether this is from decreased adherence, biological factors, or both. 4 Although social, high-risk behaviors, access and utilization of health care services, sexual violence, and economic factors may all contribute to increased susceptibility to HIV infection in young women, biological factors could also play a significant role and need to be better understood. In fact, younger individuals experience a higher rate of HIV transmission per coital act, suggesting the importance of biological factors in increased susceptibility of adolescents. 5
The major route for HIV transmission in women during heterosexual intercourse is through the mucosal surface of the female genital tract (FGT). Studies have shown that mucosal inflammation 6 –8 and a decrease in epithelial barrier function 8 –10 are associated with increased sexual transmission and acquisition of HIV, although the mechanisms are not completely understood. Inflammatory factors include increased proinflammatory cytokines, an altered vaginal microbiome, increased number of target cells, their activation status, and phenotype. 11 It is thought that these characteristics are increased in young women, possibly linked to maturation processes in the FGT.
Ovulation is commonly irregular during the first years after menarche, and the transformation zone exhibits a higher degree of cervical ectopy, 12 characterized by a single-layer columnar epithelium, which may enhance vulnerability to STI's. 13 This has been associated with increased levels of proinflammatory cytokines and chemokines in adolescents, including IL-1β, IL-8, and MIP-1α, which have separately been linked to increased HIV acquisition. 14
Vaginal microbiome alterations are also evident in adolescent women. 15,16 One study found evidence that adolescent women (age 16.9 ± 0.2) had lower levels of Lactobacillus jensenii (but not L. crispatus) species in vaginal mucosa in comparison to adult women (30.7 ± 1.1 years of age). 17 However, a statistical difference in ethnicity between these groups was also observed, therefore age may not be the underlying factor contributing to the microbiome differences. In summary, while these observations demonstrate epithelial, inflammatory, and possibly microbial differences in adolescents, they are far from complete, and whether any biological mechanism(s) are responsible for increased risk of HIV in young women has yet to be determined.
The pigtail macaque (PTM, Macaca nemestrina) model is highly relevant to HIV studies as simian immunodeficiency virus (SIV) infection in macaques largely recapitulates HIV infection in humans. 18,19 Extensive longitudinal sampling of the FGT, possible in macaques, is not as easy to perform in human cohorts and can increase infection risk. Experimental lentiviral challenge infections in animal models allow for testing of pharmacokinetics, pharmacodynamics, and ultimately efficacy of protection. In contrast to the closely related model rhesus macaques (RM) with a seasonal breeding cycle, PTM maintain monthly menstrual cycles. 20 PTM are larger in size, therefore sampling of adolescent animals is safer.
The goal of this study was to develop an NHP model for adolescent HIV infection, where mechanisms associated with increased HIV susceptibility and preclinical trials can be tested, and we hypothesized that the PTM model is ideal to compare adolescents and adults during SIV infection.
Methods
Study non-human primate macaques and sampling schedule
The macaques were housed and cared for in Association for the Assessment and Accreditation of Laboratory Animal Care international (AAALACi)-accredited facilities, and all animal procedures were performed according to protocols approved by the Institutional Animal Care and Use Committee (IACUS protocol number 2017-8711-044) of the University of Louisiana at Lafayette.
Nine adolescent (3–4 years) and ten adult (8–11 years) female PTMs (Macaca nemestrina) were sampled for 2 months during Phase I: Baseline Cycle Analysis. EDTA blood, vaginal swabs, vaginal lavages, vaginal cytobrushes, and vaginal biopsy samples were all collected at day 0, 14, 35 and 49 before infection (Supplementary Fig. S1). All animals then underwent Phase II: Low-Dose Challenge, which consisted of a weekly low-dose (LD, 4 × 104 infectious virions) intravaginal challenge of SIVmac239X until plasma viral load resulted in >100 vRNA copies/mL. Once infected, samples were collected for 1 month as part of Phase III: Infection Kinetics. All animals were euthanized 28–29 days postinfection per protocol. No data from Phase III are shown in this article.
Reproductive cycle monitoring was performed by the New Iberia Research Center research staff. A scoring criterion was used to determine changes in color and swelling of PTM sex skin, combined with timing of apparent blood from menstruation, and results were translated into luteal, follicular, or menses (Supplementary Table S1), which is a well-described method of determining menstrual cycle in NHPs. 21,22 We also measured serum progesterone and estradiol levels through mass spectrometry in a subset of the PTM (n = 5 adults, n = 5 adolescents) in this study, but we did not find it helpful in identifying menstrual cycle phases because each animal was so variable (Supplementary Fig. S2).
We used 3–4-year-old postmenarche female PTM to represent 14–16-year-old adolescent humans. This age was chosen given that PTM reach menarche at ∼2–3 years of age, and growth plates close approximately at 6 years of age in macaques (thus 6 years in PTM is approximately equivalent to 18 years of age in humans). 23 We used 8–12-year-old female PTM to represent adult humans, given that maximal bone density occurs at age 9 in macaques (thus representing ∼35 years of age). 24 Further support for this age range in macaques comes from observed differences in immunity demonstrated in RMs that were less than 4 years of age compared with those 8–15 years of age. 25
Flow cytometry staining
Flow cytometry was performed to distinguish innate and adaptive immune cell subsets (Supplementary Fig. S3) as well as distinguishing HIV target cells (Supplementary Fig. S4). Cells from vaginal biopsies were stained with LIVE/DEAD Fixable Aqua Dead Cell Stain (Thermo Fisher). Cells were then stained using surface antigen antibodies, permeabilized using Cytofix/Cytoperm (BD Biosciences, La Jolla, CA), and then stained using intracellular antigen antibodies. Details of antibodies can be found in supplementary methods. Stained samples were fixed in 1% paraformaldehyde and collected on an LSR II (BD Biosciences). Analysis was performed using FlowJo software (version 10.4.2).
Assessment of cytokines and soluble factors
Cytokine and chemokine levels in vaginal cytobrush supernatants were analyzed using the Milliplex MAP (multianalyte profiling) Nonhuman Primate Cytokine Magnetic Bead Panel Premixed 23-Plex Kit (Millipore, Darmstadt, Germany). The levels of the analytes were assessed on a MAGPIX Multiplexing Instrument (Luminex Corp. Austin, TX).
Protein extraction and digestion
Proteins were extracted from frozen vaginal biopsies as previously described. 26,27 Briefly, frozen tissue samples were added to 3 mL of lysis buffer and homogenized, centrifuged, and aliquoted. Final supernatants were pooled, and 700 μL of extracted protein was denatured for 10 min with urea exchange buffer, then passed through a Nanosep 10K cartridge. Cartridges were incubated with 50 mM iodoacetamide, and washed with 50 mM HEPES buffer. Filters were incubated with benzonase solution for 30 min, washed, then incubated overnight with trypsin. Peptides were eluted, dried, and stored at −80°C until ready for mass spectrometry analysis.
Reversed-phase liquid chromatography and mass spectrometry
Samples were cleaned using reversed-phase liquid chromatography as previously described. 27,28 Samples were dried (speed vacuum) and quantified using the LavaPep's Fluorescent Peptide and Protein Quantification Kit (Gel Company) according to the manufacturer's protocol. Equal amounts of peptides for each sample were injected into a nanoflow liquid chromatography system (Easy nLC, Thermo Fisher) connected inline to a Q Exactive Quadrupole Orbitrap mass spectrometer.
MS spectra were processed by Progenesis (Nonlinear Dynamics) and Mascot (Matrix Science) as previously described. 29,30 Technical variability was determined by the addition of a protein mix. Search results were entered into Scaffold (v4.4.1.1; Proteome Software, Portland, OR) to determine protein identifications (80% peptide confidence; 95% protein confidence, with minimum of two unique peptides per protein).
DNA extraction and 16S sequencing
Genomic DNA was extracted from vaginal swab specimens using lysis buffer, chicken egg white lysozyme, SDS, and an RNA/DNA All-prep Kit:DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA). DNA for 16S rRNA sequencing was processed using the Earth Microbiome Project protocols.
Briefly, during library preparation, DNA samples were amplified in triplicate using the FailSafe™ PCR System (Epicentre, WI) and the 515FB-806RB primer pair to generate a 400 bp amplicon from the V4 variable regions of the 16S rRNA gene. The triplicate reactions were pooled, quantified using the Qubit dsDNA High-Sensitivity Assay Kit (Thermo Fisher Scientific, Waltham, MA), and visualized using a LabChip GX (PerkinElmer, MA). Ten nanograms of each library were pooled, cleaned using the MO BIO UltraClean® PCR Clean-Up Kit (MO BIO) and quantified using the KAPA Library Quantification Kit (KAPA Biosystems). Seven pM of the pooled libraries with 30% PhiX phage as a control was sequenced using the 300-cycle Illumina MiSeq Kit, generating 150 bp paired-end reads.
Analysis of 16S rRNA gene sequencing data
Raw 16S rRNA gene sequence data were processed using the QIIME 2 software package through the DADA28 pipeline option, performing quality filtering and identification of exact amplicon sequence variants (ASVs). The SILVA database (release 132) was used for taxonomic assignments. ASVs unassigned at the Kingdom level were filtered out as well as those present in only one sample. OTU ID's were binned to the lowest phylogenetic level identified, except species level which was binned to genus. Any genera identified as having multiple unique OTU ID's had reads from each unique ID summed for final read count within the genera. The total number of reads per genera was summed across all samples.
The most prevalent genera were shown, whereas those making up <1% of reads were binned as “Other”. Trending bacterial changes were determined by Fishers exact t-test, and statistically significant changes to the bacterial composition were determined using multiple comparisons (FDR, Bonferroni).
Statistics
Statistical analyses on immune cells were performed using GraphPad Prism statistical software (version 7.0d; GraphPad Software, San Diego, CA). An average of all four baseline time points was used for each PTM. Statistical significance between adolescent and adult PTMs was calculated using a Mann–Whitney test. p < .05 (trending) and Benjamini–Hochberg adjustment p value p adj <.05 (significant). A Kaplan–Meier plot was used to examine the infection curves of adolescent and adult PTMs. Statistical significance was determined by a Log-rank (Mantel–Cox) test.
Protein abundance values were generated with Progenesis Software. Outliers having a median normalized abundance greater than one standard deviation of all samples in the run were removed before analysis. Proteins with <25% covariance were used for downstream analysis after log2-transformed.
Protein differences were determined using paired t-tests, comparing adult and adolescent protein expressions for all times sampled (Day 0, 14, 35, 49), as well as just follicular phase samples to adjust for menstrual cycle differences in expression. A FDR of q < 0.05 was applied to correct for multiple comparisons using the Benjamini–Hochberg method. Significance values below an unadjusted α = 0.05, but did not pass multiple hypothesis testing, were reported as statistical trends. Unbiased hierarchical clustering with a Euclidean distance metric (RStudio; Version 1.2.5033) was performed, as well as PCA plots (RStudio) and volcano plots (Prism; Version 8.4.3).
Results
Adolescent PTMs are not more susceptible to repeated LD vaginal challenges of SIVmac239
The average lifespan of a female macaque is ∼25 years to a maximum of 40 years. 31 There is evidence that a menopausal-like state occurs in these animals at around 25 years, with the onset of puberty occurring at ∼3.5 years. 31 We examined mucosal characteristics of the FGT in 9 adolescent PTMs (3.9 ± 0.49 years old) and 10 adult PTMs (9.81 ± 0.97 years old). The effects of the menstrual cycle on the FGT was observed with sex skin swelling used to perform a 2-month baseline cycle analysis of SIV susceptibility. We performed an in-depth FGT assessment of immunological parameters in adolescent and adult PTMs pre-SIV infection throughout the menstrual cycle.
To determine if adolescent PTMs were more susceptible to SIV infection, we performed a weekly low-dose intravaginal challenge of SIVmac239X (4 × 104 infectious virions) until infection occurred in all 19 animals. We were powered to detect a 2.2-fold increase in infection rates. Infection status was deemed positive once plasma vRNA hit greater than 100 copies/mL. The Kaplan–Meier plot (Fig. 1) reveals that infection curves between the 9 adolescent PTMs and 10 adult PTMs were not significantly different.

Vaginal susceptibility to repeated low-dose SIVMac239X in adult and adolescent PTMs. Kaplan–Meier plot comparing infection curves of adult and adolescent PTMs in repeated low-dose (LD; 5 × 10E4 copies/mL) vaginal challenges of SIVmac239. Plasma vRNA >100 copies were deemed infected. Infection curves are not significantly different and were determined by a Log-rank (Mantel–Cox) test. PTM, pigtail macaque; SIV, simian immunodeficiency virus.
Differences in innate and adaptive immune cell frequencies in vaginal tissue of adult and adolescent PTMs
Flow cytometry was performed on cells isolated from vaginal biopsies to examine immune cell subsets different between adult and adolescent PTMs. Comparisons that were p < .05 using a univariate/unadjusted analysis (Mann–Whitney U) but did not pass multiple corrections p-value adjustments (p adj. >0.05, Benjamini–Hochberg) were considered trending. No trends were detected in the frequency of CD4+ T cells (Fig. 2A), however lower frequency of CD8+ T cells in the vaginal tissue of adolescent PTMs were trending (p = .0369, adj. p = .10875; Fig. 2B). Neutrophil, monocyte, and B cell frequencies were also compared between age groups. No differences were detected in vaginal neutrophils or monocytes, but adolescent PTMs trended higher HLA-DR+CD20+ B cells in vaginal tissue compared with adults (p = .0435, adj. p = .10875; Fig. 2E).
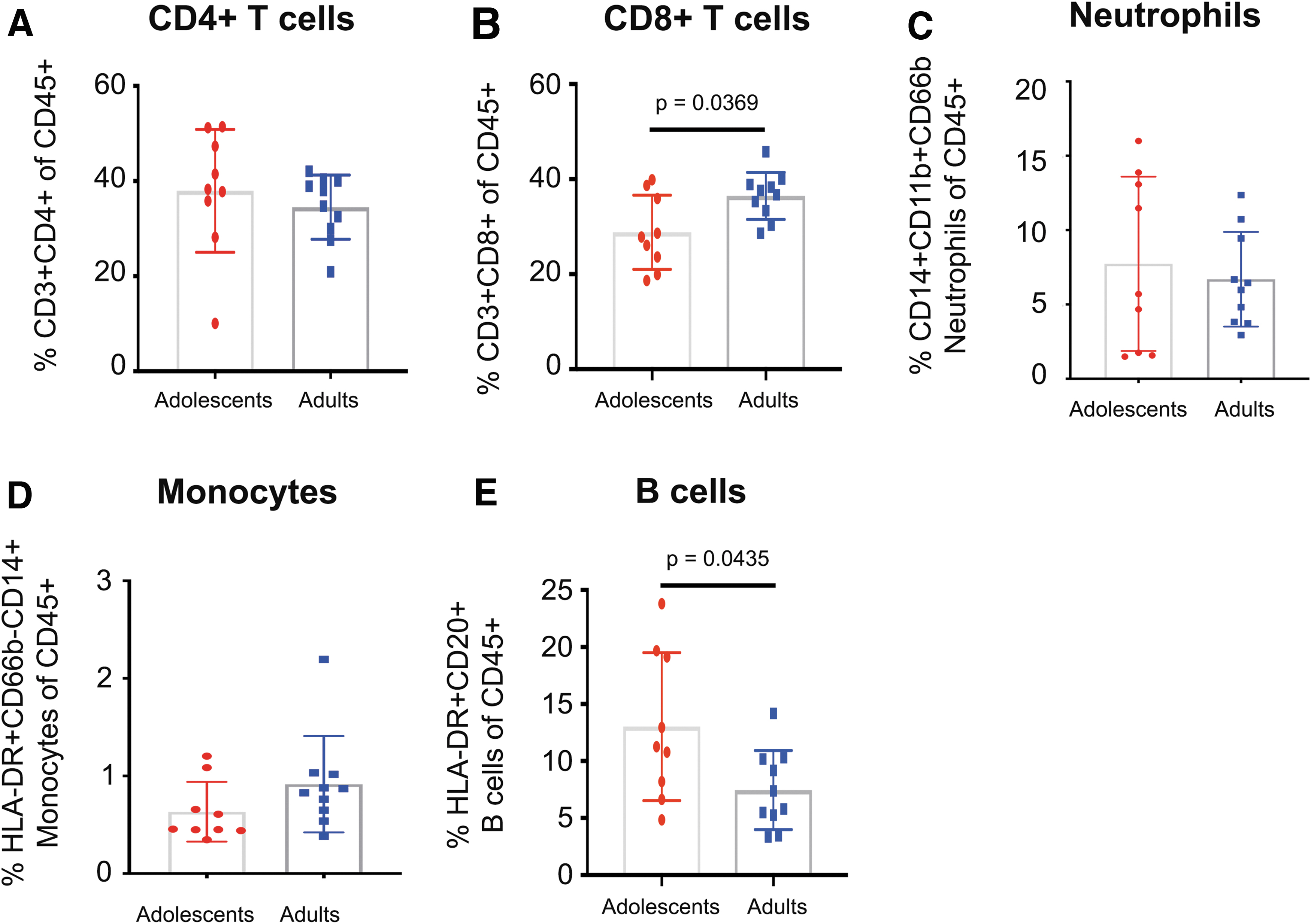
Innate and adaptive immune cell frequencies in vaginal tissue of adult and adolescent PTMs. Flow cytometry was used to determine frequencies of various innate and adaptive immune cells isolated from vaginal biopsies.
We also compared innate and adaptive cell frequencies across the different phases of the menstrual cycle. No significant differences were detected in the frequency of T cells, neutrophils, monocytes, or B cells between menses, follicular, or the luteal phase of the PTMs (Fig. 3). A caveat of this dataset is that we were not able to capture all three menstrual phases for each animal. The luteal phase lacked power with only 5 of the 19 animals having a time point during the luteal phase of their cycle. Much like humans, the luteal phase of the PTMs is short and can vary between and within animals, making it difficult to capture without very frequent sampling. 32

Innate and adaptive immune cell frequencies throughout the menstrual cycle of adult and adolescent PTMs. Flow cytometry was used to determine frequencies of various innate and adaptive immune cells throughout the menstrual cycle.
No difference in SIV target cell frequencies in vaginal tissue of adolescent and adult PTMs
The frequencies of specific CD4+ target cells expressing HIV coreceptors were measured in vaginal biopsies by flow cytometry to characterize the specific lineages that relate to HIV acquisition. TH17-lineage CCR6+ CD4+ T cells have been identified as primary targets of SIV during vaginal transmission. 33 Expression of α4β7 on CD4+ T cells and levels of coreceptor CCR5 and CXCR4 have been shown to directly influence HIV/SIV infection. 34
Within this study, there were no significant differences in any of the previously specified HIV target cells when compared between adolescent and adult PTMs (Fig. 4). Relatively higher frequencies of CXCR4+ CD4+ T cells were detected in vaginal tissue of both adolescent and adults compared with the other target cell subsets, such as CCR6+, CCR5+, and α4β7+ CD4+ T cells. To further investigate the effects of the menstrual cycle on SIV susceptibility, we compared HIV target cell frequencies across each phase of the cycle, but again no significant changes were detected (Fig. 5).

Frequencies of HIV target cells in vaginal tissue of adult and adolescent PTMs. Flow cytometry was used to determine frequencies of HIV target cells isolated from vaginal biopsies.
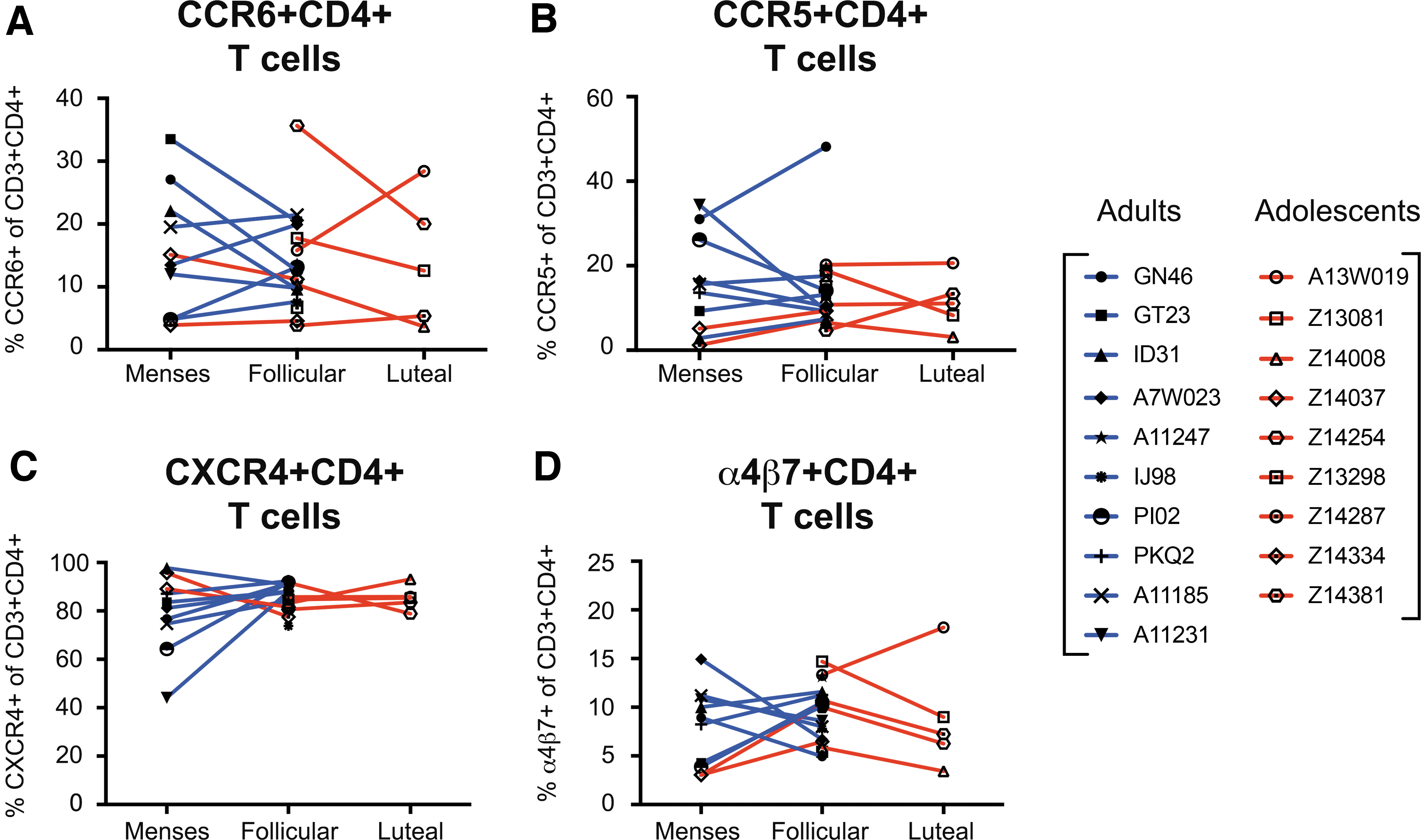
Frequencies of HIV target cells throughout the menstrual cycle of adult and adolescent PTMs. Flow cytometry was used to determine frequencies of HIV target cells throughout the menstrual cycle.
Differences in inflammatory cytokines and chemokines in vaginal fluids of adult and adolescent PTMs
Studies to date have demonstrated that mucosal inflammation is associated with increased sexual transmission and acquisition of HIV, 35 and is a major factor associated with adverse sexual and reproductive outcomes in adolescent females although the mechanisms are not completely understood. 36
Our data revealed that soluble CD40 ligand (sCD40L) was trending higher (adj. p = .606) and monocyte chemoattractant protein-1 (MCP-1) was significantly higher (passing multiple comparisons, adj. p = .02) in adult PTMs (Fig. 6C, D) in vaginal fluids, which have been associated with increased risk of HIV acquisition. 37,38 Other proinflammatory cytokines, such as, IL-8, IL-1Rα, I IFNg, and TNFα that have previously been associated with increased HIV infection 39,40 were measured from vaginal cytobrush supernatant, but no differences were detected between adolescent and adult PTMs (Fig. 6).
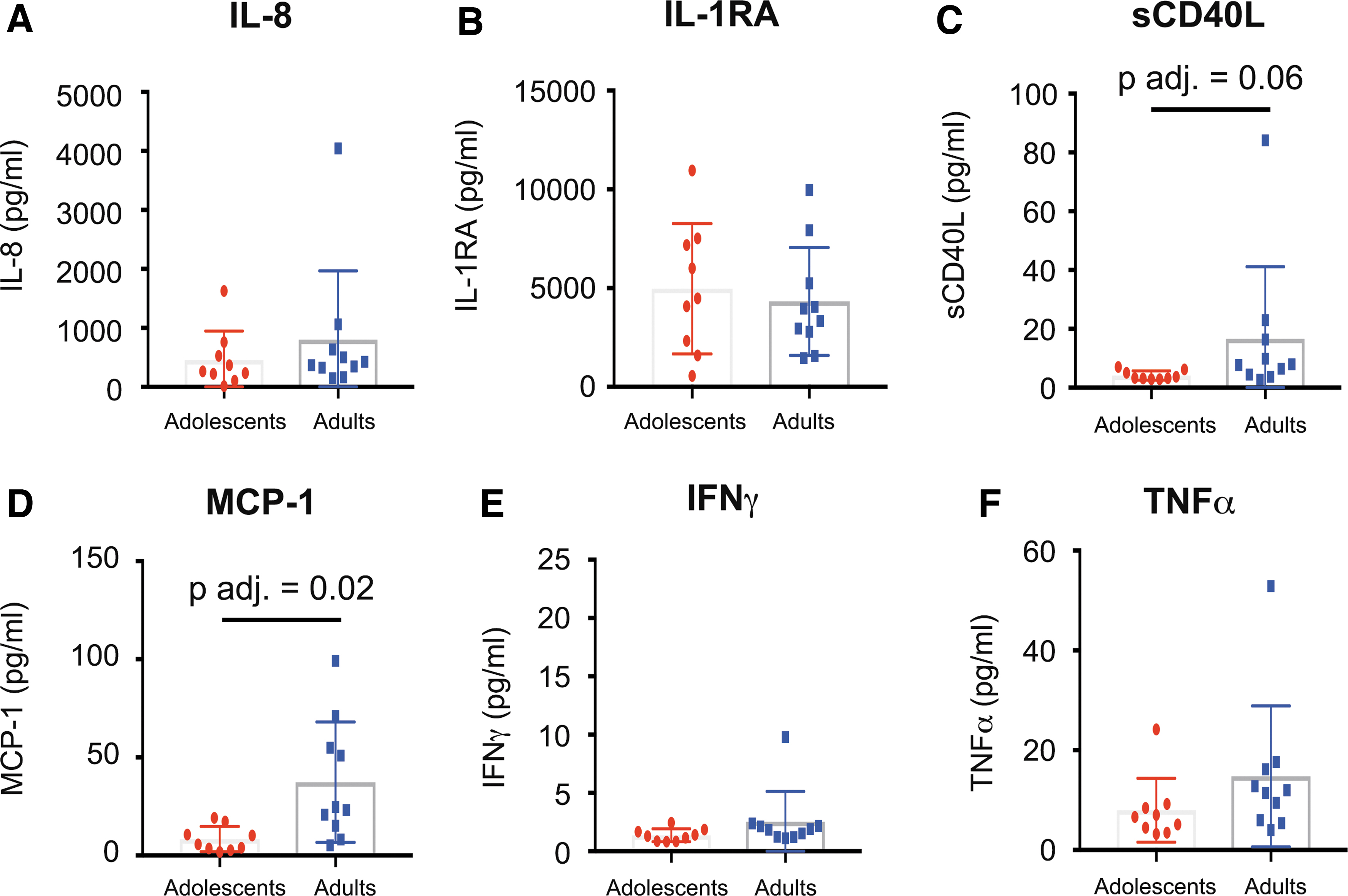
Inflammatory cytokines and chemokines in vaginal fluids of adult and adolescent PTMs. Luminex assay was performed on vaginal cytobrush supernatant to determine concentrations of proinflammatory cytokines in the FGT of adult and adolescent PTMs.
Overall, when examining inflammatory cytokines and chemokines, we saw no significant differences between menstrual phases (Fig. 7). Previous studies have hypothesized that the luteal phase of the menstrual cycle is a more vulnerable time for SIV/HIV acquisition due to increased HIV target cells or increases in proinflammatory cytokines 32,41,42 ; however, in our study, most infections (80% overall, 75% of adults and 87.5% of adolescents) occurred during the follicular phase, and we were underpowered to observe menstrual phase differences as we did not capture any adult luteal phase samples.

Inflammatory cytokines and chemokines in vaginal fluids of adult and adolescent PTMs throughout the menstrual cycle. Luminex assay was performed on vaginal cytobrush supernatant to determine concentrations of proinflammatory cytokines in the FGT of adult and adolescent PTMs throughout the menstrual cycle.
No differences in the vaginal tissue proteome in adult and adolescent PTMs
Previous literature has indicated that increased inflammation as well as a loss in barrier function may be mechanisms for increased risk of HIV. To determine barrier function or inflammatory signaling differences, we performed proteomics on vaginal biopsies taken longitudinally at different times during the menstrual cycle in 5 adult and 5 adolescent PTMs. We were powered to observe a 1.8-fold change difference in protein expression between age groups.
A total of 1687 proteins were identified for all macaques at each time point. After correcting for multiple comparisons, there were no significant differences in protein abundance between age groups from all time points (Fig. 8B). Samples collected only during follicular phase, the phase most sampled, also did not show proteome differences. Euclidean clustering of protein abundance values showed that samples do not cluster by age group (Fig. 8A). Principal component analysis (Fig. 8C) of all samples collected in follicular phase illustrates a nearly complete overlap of vaginal tissue proteomes between adults and adolescents.
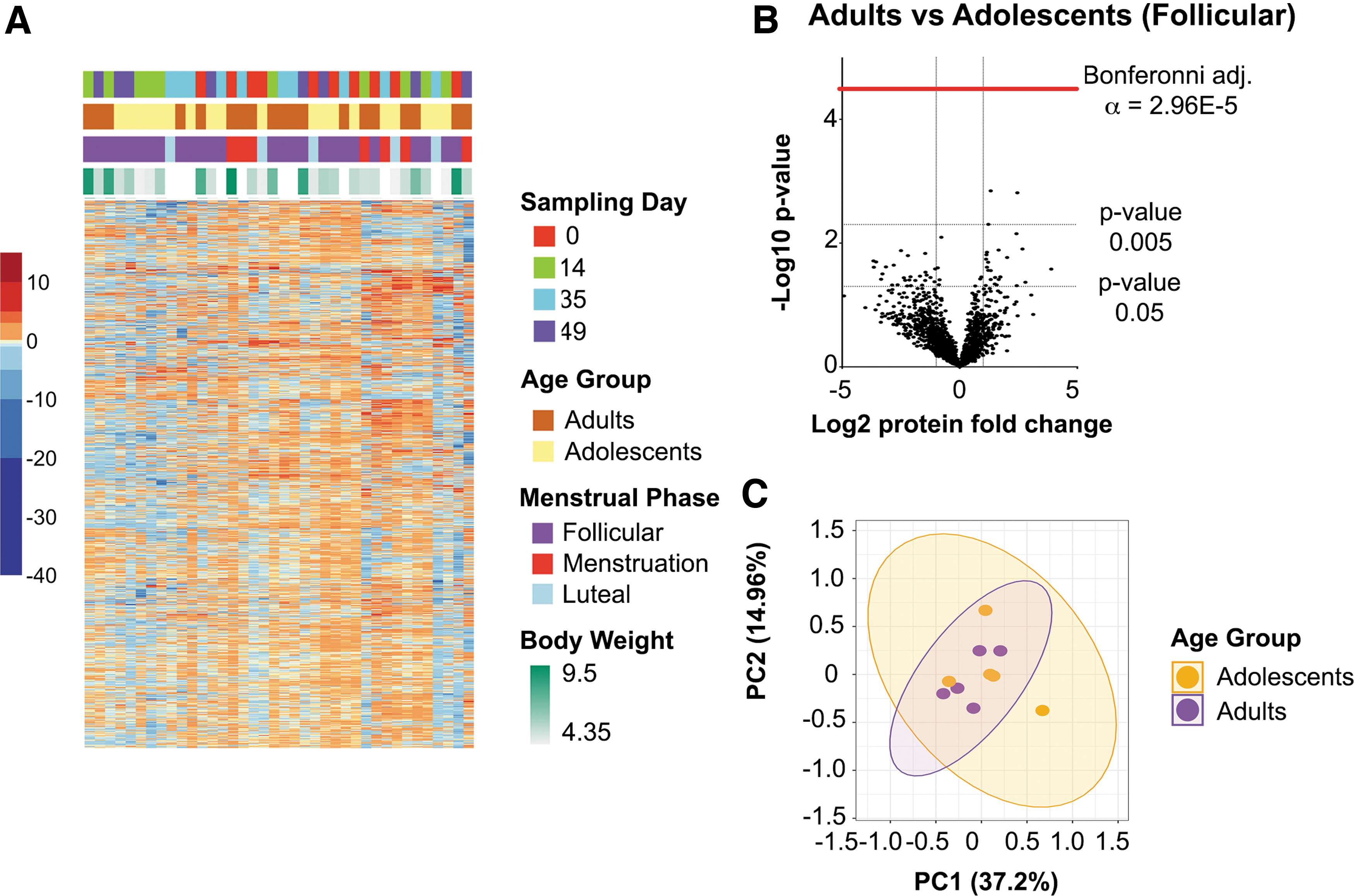
Vaginal tissue proteome differences between adult and adolescent PTMs. Mass spectrometry-based proteomics was used to determine changes in host cellular function of the vaginal barrier.
Alterations in vaginal microbial communities of adult and adolescent PTMs
DNA was extracted from vaginal swabs collected from 9 adolescent and 10 adult PTMs over four time points throughout Phase I. The 16S rRNA sequencing was performed to identify bacterial genera differences between adolescent and adult PTMs. When comparing age groups, only time points collected within the follicular phase (averages for multiple time points) were used, since this was the only overlapping menstrual phase for both groups.
After adjusting for multiple comparisons (Bonferroni) no significant differences were detected between adolescent and adult PTMs, however, some trends in microbiome differences were observed (Fishers exact t test, p < .05). Adolescent PTMs had generally higher relative abundances of Sneathia, Prevotella, and Streptococcus genera, whereas adults had trending higher abundances of Slackia, Fastidiosipila, and Corynebacterium (Fig. 9A). Sneathia and Prevotella species have been identified as bacterial vaginosis (BV)-associated bacteria and have been linked with several adverse health outcomes, including HIV acquisition. 43

Vaginal microbial communities of PTMs. 16S sequencing was performed on vaginal swabs of adolescent and adult PTMs throughout different phases of the menstrual cycle.
Various factors can influence the stability of the vaginal microbiome, including hormonal shifts and the start of menses. 44 To investigate microbial variations that occur over the menstrual cycle, bacterial genera were compared during menses, and follicular and luteal phases (Fig. 9B). Again, no significant differences were detected after adjusting for multiple comparisons (Bonferroni), but trends were identified between different cycle phases (Fishers exact t-tests, p < .05). A higher relative abundance of Atopobium and Actinomyces was observed during the follicular phase in comparison to luteal and menses. In the luteal phase, less Mobiluncus was trending in comparison to the other phases. During menses, more Fusobacterium was trending and less Sneathia bacterium, when compared with the other phases of the menstrual cycle.
Discussion
A validated SIV NHP model of HIV infection in human adolescents is currently unavailable. Preclinical NHP trials do not currently consider the effects of age on infection susceptibility, and ethics approval is more arduous to enroll women under 18 years of age in human trials. The development of an infection model, which can accurately delineate infection susceptibility, correlates of infection, and potential mechanisms underlying altered FGT in adolescents, is needed. In this study, we undertook a highly comprehensive approach to investigate HIV infection risk in adolescent female PTMs and evaluate this animal model.
In our study, we did not detect increased susceptibility to SIV infection in adolescent PTMs compared with adults, or any differences in vaginal CD4+ T cell frequencies between 9 adolescents and 10 adults. We were underpowered to investigate immune cell differences between age groups during menses or the luteal phase, therefore our analysis focused on differences during the follicular phase. Despite this, we were able to identify some trending differences in mucosal immune characteristics that have yet to be described. Slightly lower frequencies of CD8+ T cells and higher frequencies of HLA-DR+CD20+ B cells were observed in adolescents.
A decrease in cytotoxic T cells at the site of viral transmission could play a role in increasing susceptibility to viral pathogens within the FGT. CD8+ T cells are a critical component of the cellular immune response to viral infections and play a key role in eliminating virally infected cells. 45 In addition, HIV can bind to B cells through the binding of complement receptor CD21 (CR2) expressed in most mature B cells, and circulating complement proteins bound to HIV virions. 46 This interaction may facilitate cell-to-cell transmission of HIV. 47 Previous studies have shown that ovarian sex steroid levels have an influence on FGT immunity, specifically the relationship between CD8+ T cells and B cell functionality. 48 Therefore, hormonal-level differences between adolescent and adults may contribute to immune cell differences and needs further investigation.
Th17 lineage CCR6+CD4+ T cells have been identified as primary targets of SIV during vaginal transmission in RMs, 33 and are also important for maintenance of epithelial barriers. 49 Studies have also shown that in humans, dendritic cells (DCs) are the first to take up HIV-1 and that the most robust viral replication occurs within T cells of the genital tract. 50 Through flow cytometry we quantified frequencies of HIV target cells from vaginal biopsies. We detected no difference in CCR6+CD4+ T cell frequencies, HIV coreceptors (CCR5 and CXCR4) and the mucosal homing marker integrin α4β7 between adolescent and adult PTMs.
Inflammation within the FGT can cause damage to the cervix and vaginal epithelium, increasing the risk of acquiring sexually transmitted diseases, including HIV. 51 Indeed, studies have elegantly demonstrated that genital inflammation contributes to HIV susceptibility in women. 39,40,52,53 When examining inflammation within the FGT of PTMs, we discovered that the adults had higher concentrations of MCP-1 with sCD40L. This could be a result of increased exposure to outside pathogens due to their older age. However, the adult baseline time points were only taken during the follicular or menstruation phase, phases associated with increased inflammation, 54 while the adolescent group had only two animals that underwent sampling during menses.
This caveat could be regulated by hormonal contraceptives to synchronize menstrual cycles of the NHP model in future studies. For this study, we decided to not interfere with the menstrual cycle due to lack of information regarding natural differences of inflammation between age groups.
Clinical studies have linked inflammatory cytokines in the female reproductive tract with host proteome changes related to barrier integrity and neutrophil proteases 8 that associate with an influx of HIV-susceptible target cells. We measured protein expression in the vaginal tissues to determine if there were any host cell functional differences that may underlie this increase in HIV susceptibility. As with infection rates, there were no significant differences in protein changes between age groups, although small sample sizes precluded us from having power to detect small differences.
BV is common in women associated with adverse health outcomes such as pelvic inflammatory disease and increased HIV transmission. 15,55 It has been reported that adolescents can present with altered vaginal microbiota, 51 which is a potential mechanism for increased HIV acquisition, and adolescent women are largely Lactobacillus dominant although literature on the subject is limited. 56 Studies comparing adult with adolescent women in the microbiome are also lacking but may suggest a difference in microbiome profiles, 17 although some were not able to determine if this difference was due to altered estrogen levels. 57
From our results, both the adult and adolescent PTMs revealed a vaginal microbiome with high diversity, and although there were no statistically significant differences, the adolescent group did have higher relative abundances of Sneathia and Prevotella genera, which have been associated with inflammation and BV in humans. 43 When comparing all PTMs in this study, trending lower abundance of Mobiluncus was identified during the luteal phase compared with the other two phases, Sneathia trended higher during menses, and numerous bacterial genera trended higher during the follicular phase, including Atopobium.
Neither age group had an abundance of Lactobacillus-dominant vaginal microbiome profiles, which is the optimal profile for human female reproductive health, but this might not be the case in NHPs. 58 This BV-like state in PTM may be due to the natural environment of the macaques to favor non-Lactobacillus-dominant profiles, or due to behaviors such as introducing foreign objects into their vaginal tracts. It should not be assumed that indigenous microorganisms in PTMs function the same way as human microbiota, and this PTM model may not accurately represent the human FGT and microbiome interactions that could lead to increased HIV susceptibility.
Therefore, this model cannot accurately represent differences in the vaginal microbiome in humans, and although we did not observe differences in the microbiome between age groups in PTMs, this does not mean that there are no differences in human adolescent and adult vaginal microbiomes.
Finally, this model may not capture other biological factors that can be observed to play a role in HIV susceptibility in humans, such as an ectropion cervix, which has been observed to increase risk of infection by a two-fold increase for women under 19 years of age in South Africa. 59 In our study, we did not observe any indicators of ectropion cervix in the PTMs, and this may be less likely in macaques.
Conclusion
In conclusion, our attempt at defining a clinically translatable NHP adolescent SIV model did not result in any mechanistic biological insights for increased susceptibility in adolescents. It is possible that differences, if they exist, are subtle, which would require a larger number of animals for sufficient power to detect these features, or that biological differences in human adult and adolescents are not represented in this model. Nevertheless, the present study is, to our knowledge, the first to compare mucosal characteristics of the FGT in adolescent and adult PTMs pre-SIV infection to determine what mechanisms may play a role in SIV susceptibility.
Future HIV prevention research must consider age and hormonal cycling when performing these microbiota-associated human disease studies in females. Sociobehavioral factors should also be accounted for when studying populations of different cultural backgrounds but are not variables in NHP models. Although no differences were observed in this PTM model, young women are a key population at risk for HIV infection as they are twice as likely to acquire HIV as their male peers (UNAIDS 2016 estimates). Therefore, concerted efforts to expand prevention options available to young women in terms of development of novel HIV-specific biomedical, structural, and behavioral interventions are urgently needed for epidemic control of this uniquely vulnerable population.
Footnotes
Acknowledgments
The authors would like to thank Stuart McCorrister and Garrett Westmacott at the National Microbiology Laboratory for mass spectrometry technical support.
Author Contributions
C.M., N.R.K., M.A., F.V., A.D.B., A.R.B., and T.J.H. designed and coordinated the PTM study. C.A.B. performed all DNA extractions from vaginal swabs and rRNA 16S sequencing. C.B.D. performed 16s rRNA analysis and immune marker statistics. M.A. collected all samples and processed vaginal biopsies and stained isolated cells for flow cytometry markers. E.C. assisted C.M. with acquiring flow cytometry samples on LSRII instrument. C.M. performed all Luminex experiments. L.N.-R. performed proteomic QC and statistical analysis. A.R.B. performed proteomic data analysis, and final generation of figures. L.N.-R. and A.R.B. performed statistical analysis of 16S sequencing. J.M. and T.H.-M. provided feedback on data interpretation and article edits. M.A. and F.V. led clinical care of PTMs at New Iberia Research Center. C.M. and N.R.K. led the PTM study and interpreted results. A.R.B., C.M., N.R.K., and A.D.B. contributed to writing the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Funding for these studies was from NIAID R01AI128782 awarded to MPIs N.R.K., A.D.B., and 641 T.J.H.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.