
Abstract
Human immunodeficiency virus type-1 (HIV-1) antigenic variation poses a great challenge for vaccine immunogen design to elicit broadly neutralizing antibodies (bNAbs). Over the last 10–15 years, great progress has been made to understand the conserved sites of sensitivity on HIV envelope glycoprotein spikes targeted by bNAbs. Plasma neutralization mapping and monoclonal antibody isolation efforts have revealed five major conserved epitope clusters. Most of this work has focused on subtype B and C-infected Caucasian or African donors. It is not clear if the same epitopes and epitope rank order preferences are also true in donors infected with different HIV-1 subtypes and with different racial backgrounds. To investigate this point, in this study we report the first attempt to profile the bNAb specificities of CRF01_AE-infected Malaysian plasmas. We first measured neutralization titers of 21 plasmas against a subtype A, B, and AE pseudovirus panel. This revealed that 14% (3 of 21) plasmas had cross-clade breadth. Focusing on the cross-neutralizing plasma P9, we used AE and JR-FL mutant pseudoviruses, gp120 monomer interference, and native polyacrylamide gel electrophoresis to better understand the neutralization specificity. P9 demonstrates CD4-binding-site specificity with trimer dependence and D368 independence.
Introduction
It is over 30 years since human immunodeficiency virus type 1 (HIV-1) was identified as the causative agent of acquired immunodeficiency syndrome (AIDS) and an effective HIV-1 vaccine is still desperately needed. 1 –3 The adaptive immune response is usually unable to control HIV-1 replication, in part, due to the depletion of CD4 T cells. Neutralizing antibodies (NAbs) target Envelope glycoprotein (Env) spikes on HIV-1 surfaces, thereby preventing infection. Eliciting broadly neutralizing antibodies (bNAbs) became the main focus of vaccine design and many bNAbs have been recovered from chronically infected donors over the last decade. 4 bNAbs generally target five major epitope clusters, namely the V2 apex, V3-glycan, CD4-binding site (CD4bs), interface, and membrane-proximal ectodomain region. These bNAbs provide paradigms for vaccine developers trying to elicit similar bNAbs. 4 –9
HIV-1 subtypes A, B, and C are responsible for most infections worldwide and are generally confined to Africa, North America, and Europe. 10 In Southeast Asia, the circulating recombinant form CRF01_AE (AE) is the major circulating subtype. 11 –17 Efforts of mapping the bNAb specificities of HIV plasmas have largely focused on subtypes B and C. 18 –24 To date, neutralizing serology and specificity of subtype AE-infected Malaysians has not been reported. It is possible that different HIV-1 Env subtypes could induce distinct NAbs related to their inherent genetic differences. Indeed, it was reported that clade AE and B sera form neutralizing serotypes more effectively that neutralize clade-matched viruses. 25 Furthermore, several of the bNAbs recovered over the last decade are unable to effectively neutralize HIV-1 AE strains. 9,26
Given the evidence of enhanced intrasubtype reactivity of AE viruses and their cognate NAbs, we investigated, in this study, bNAb specificities in Malaysian AE infections. The information we recover should help to ensure that vaccine designs induce sufficiently broad NAbs that can crossreact with AE viruses and therefore curtail the epidemic in South East Asia.
Methods
HIV-1 CRF01_AE-infected plasma samples
HIV-1 CRF01_AE-infected plasmas (n = 21) were identified by screening of a pool of donor sera at the National Blood Center of Kuala Lumpur between 2013 and 2014. 11 Plasmas were given a numerical code (e.g., Plasma 5, Plasma 8) and were abbreviated (e.g., P5, P8). Uninfected human plasma was included as a negative control (Sigma-Aldrich). This study was approved by the University Malaya Medical Center (UMMC) Medical Ethics Committee (MEC reference number: 824.2) and Sunway University (SUNREC 2017/031).
HIV-1 Env-bearing plasmids and anti-HIV-1 Env monoclonal antibodies
HIV-1 Env-expressing plasmids include simian immunodeficiency virus SIVmac239; subtype A: BG505; subtype B: WITO, JR-FL, and JR-FL SOS. 27 –29 SOS mutations introduce an intermolecular disulfide bond between gp120 and gp41. AE Env-bearing plasmids C1080.c03, CM246.c1, and 816763.c02 were provided by David Montefiori (Duke University). 30
Anti-HIV-1 Env monoclonal antibodies (mAbs) were obtained from NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH. MAbs include: b12, directed to the CD4bs from Dennis Burton and Carlos Barbas 31 ; VRC01 and VRC03 directed to the CD4bs from John Mascola 32 ; 2G12 directed to N-linked glycans of gp120 from Polymun Scientific 33 ; 17b directed to the CD4-induced (CD4i) discontinuous epitope of gp120 from James E. Robinson 34 ; PG9, PG16, and PGT145, directed to quaternary epitopes of the gp120 V1/V2 loops from the International AIDS Vaccine Initiative (IAVI), 7,8 PGT121 directed to the N332-centered oligomannose patch on the V3 loop, 7 and 10E8, directed to the membrane proximal region (MPER) of the Env, from Mark Connors. 35
AE recombinant 93TH975 and CM235 gp120s were obtained from the NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH; and A244 D11 gp120 was obtained from Barton F. Haynes and Hua-Xin Liao. 36 Subtype B recombinant AD8 gp120 and gp140 were provided by Rob Center (Burnet Institute, Australia) 37 ; Subtype C recombinant 96ZM651 gp120 and CN54 gp120 38 were obtained from NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH. MN gp41 was obtained from ImmunoDX, LLC.
Enzyme-linked immunosorbent assay
HIV-1 gp120 or gp41 (5 μg/mL) were coated on ELISA microplates (Greiner Bio-One, Austria) overnight at 4°C. Following a phosphate-buffered saline Tween 20 (PBST) wash and blocking with 4% skim milk in PBST, graded dilutions of mAbs or plasmas were titrated. Goat anti-human IgG conjugated with alkaline phosphatase (Accurate Chemical) and SigmaFAST p-nitrophenyl phosphate tablets (Sigma-Aldrich) were used to detect binding, both with 1 h incubation each. 3N NaOH was used as the stop solution. Plates were read at optical density 405 nm using a microplate reader (Tecan Infinite 200 PRO, Switzerland).
Site-directed mutagenesis
Mutant HIV-1 Env pseudovirions (PVs) were generated through site-directed mutagenesis using the QuikChange Lightning Site-Directed Mutagenesis Kit (Agilent Technologies). Library Efficiency™ DH5α Competent Cells (Invitrogen) were transformed with mutant plasmids. Point mutations were confirmed by Sanger sequencing.
Neutralization assays
HIV-1 PVs were generated by transiently transfecting HEK293T cells with subgenomic plasmid
Standard neutralization format
Standard neutralization assays were conducted as previously described. 27 HIV-1 PVs were incubated with various plasmas or anti-Env mAbs at graded dilutions for 1 h and spinoculated before adding to the target cell. Luciferase activity in the cell lysate was measured using Luciferase Assay System (Promega) 3 days post-transfection. Luminescence was measured by microplate reader Infinite 200 PRO (Tecan, Switzerland) in relative luminescence units (RLU) that are directly proportional to the number of successful infections by HIV-1 PVs.
Post-CD4/CCR5 neutralization format
In the post-CD4/CCR5 format, JR-FL “SOS” PV was first allowed to attach to the
Gp120 interference
JR-FL gp120 D368R was made in the PPI4 plasmid as described previously. 42 JR-FL gp120 D368R-expressing plasmid was transfected using PEI into HEK293T cells and expressed recombinant proteins in the supernatant were purified through an affinity column containing agarose-bound Galanthus nivalis lectin (Vector). 43 In the gp120 interference assay, graded titrations of plasma or mAbs were incubated with 5 μg/mL JR-FL gp120 D368R for 1 h before incubation with HIV-1 PVs. The remaining steps were similar to the standard neutralization assay.
Native polyacrylamide gel electrophoresis Western blots
HIV-1
Results
Neutralizing activity of HIV-1 CRF01_AE-infected plasmas
Our goal was to probe the NAb specificity of HIV-1 CRF01_AE-infected plasmas from Malaysia. We first profiled plasma NAb ID50s of 21 AE-infected Malay donors against a diverse panel of tier 2 PVs, derived from subtypes CRF01_AE, A, and B. Plasmas neutralized to higher ID50s against clade-matched PVs (Fig. 1A). Thirty-eight percent of plasmas exhibited detectable NAbs against at least one strain (Fig. 1A), 33% of which neutralized at least one of the three AE strains. Plasmas P9 and P133 (mean ID50 109) showed the greatest breadth against four of the seven strains tested. Of the subtype B strains, only P9 neutralized JR-FL (ID50 of 148), only P133 neutralized WITO (ID50 of 128), and only P83 neutralized REJO (ID50 of 38). Only P9 neutralized the subtype A strain BG505, although weakly. P133 and P138 plasmas differentially neutralized subtype AE strains, with potent, moderate, or undetectable ID50s against C1080, CM246, and 816763, respectively. None of the plasmas neutralized 816763 (subtype AE) or SIVmac239.
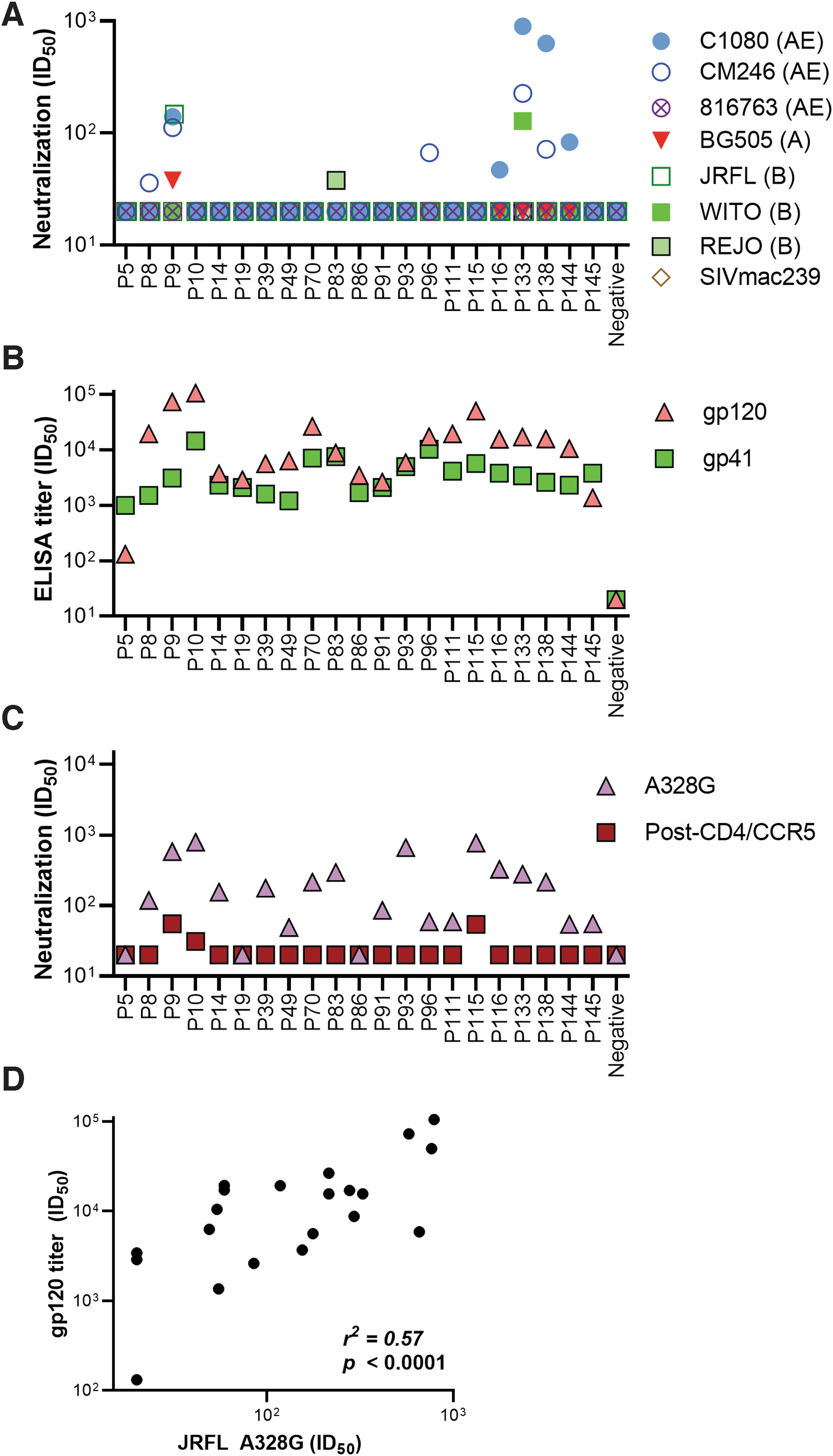
Profile of HIV-1 AE-infected plasmas. Plasma neutralization titer of <20 is plotted at 20. Gp120 ELISA titers were taken as the dilution at which plasma binding exhibited an OD of 0.5 at 405 nm. All assays were performed in duplicate. Data are representative of at least two repeats.
No clinical information on the plasma donors was available. It is therefore possible that some of the 14 non-neutralizing plasmas (Fig. 1A) derive from recently infected donors in whom Ab responses had not yet fully developed. If so, we might expect to find low gp120-binding titers, suggesting ongoing seroconversion. 44 To investigate, we measured plasma ID50s to gp120s from subtype AE, B, and C by ELISA and neutralization of the highly sensitive tier 1 JR-FL A328G mutant reported previously. 40,45
With the exception of P5, low to high anti-gp120 Ab-binding titers were observed for all plasmas (Fig. 1B and Supplementary Fig. S1). Notably, P9 and P10 showed the highest binding titer against 93TH975 (AE) and A244 (AE); and CN54 (C) with moderate activity toward CM235 (AE) and AD8 (B); and weakest against 96ZM651 (C). Aside from P5, P19, and P86, all plasmas also neutralized the JR-FL A328G PV. Indeed, gp120 titers and JR-FL A328G NAb titers correlated (r 2 = 0.57; p < .0001) (Fig. 1D).
Gp41 antibodies were probed by ELISA using recombinant gp41 and by post-CD4/CCR5 neutralization assay to detect NAbs specific toward the MPER of gp41 (Fig. 1B, C). In this format, JR-FL SOS Env binds to CD4 and CCR5 receptors, but is unable to initiate infection by its introduced disulfide bond. 42 This allows anti-gp41 antibodies to target the exposed gp41 epitopes and through the addition of DTT, the disulfide bond is removed to allow PVs to infect the target cells. All plasmas bound well to gp41 in ELISA, P10 and P96 having the highest ID50s. In the post-CD4/CCR5 neutralization assay, P9, P10, and P115 exhibited modest ID50s, but other plasmas, ID50s, were below detection. Notably, while P9 neutralized JR-FL in both the standard and post-CD4CCR5 formats, P10 and P115 weakly neutralized JR-FL only in the post-CD4/CCR5 neutralization format (Fig. 1A, C). P10 and P115 did not detectably neutralize any virus in the standard format. Although MPER NAbs usually show largely equivalent titers in both formats, 41 the discrepancy could be due to slight assay sensitivity differences. Overall, the neutralizing activities of the three best plasmas, P9, P133, and P138 (Fig. 1A), cannot be explained by MPER NAbs (Fig. 1C). This finding is in line with evidence that anti-gp41 MPER NAbs are relatively rare. 18,46,47
Overall, we can conclude that 8 out of 20 chronically infected donors exhibit detectable heterologous neutralization, with generally higher intrasubtype AE neutralization ID50s. Furthermore, this neutralizing activity did not appear to target the MPER. We focused further efforts on plasma P9, because it neutralized four of seven viruses tested, including JR-FL, a strain that has been extensively used in previous mapping studies.
Epitope specificity of plasma P9
Some NAbs target quaternary Env trimer-specific epitopes that may not be present on corresponding gp120 monomers. In contrast, other NAbs recognize these different forms of Env equivalently. To distinguish these two types of NAb, excess recombinant gp120 (with a D368R mutation to eliminate direct gp120-CD4 binding) can be added to plasmas to assay neutralization interference. 18 D368R gp120 was found to interfere with 2G12 neutralization (targets the high mannose patch of gp120 and is not trimer specific), but not b12 neutralization (the b12 epitope is knocked out by the D368R mutation) (Fig. 2A, B). Plasma P9 neutralization of JR-FL, C1080, and CM246 PVs (Fig. 1A) was unaffected by adding excess D368R gp120 monomer (Fig. 2C–E). This suggests either that P9 contains trimer-specific or D368-dependent NAbs that can neutralize these three strains. One caveat is that the interfering mutant gp120 (JR-FL) did not match two of the PV strains (C1080, CM246). However, the fact that the P9 plasma neutralized all three strains supports the idea that if gp120 could interfere with neutralization against one strain, it should likely also interfere with neutralization of other sensitive strains. Such quaternary NAb are common in plasmas from donors infected with other clades. 18,48
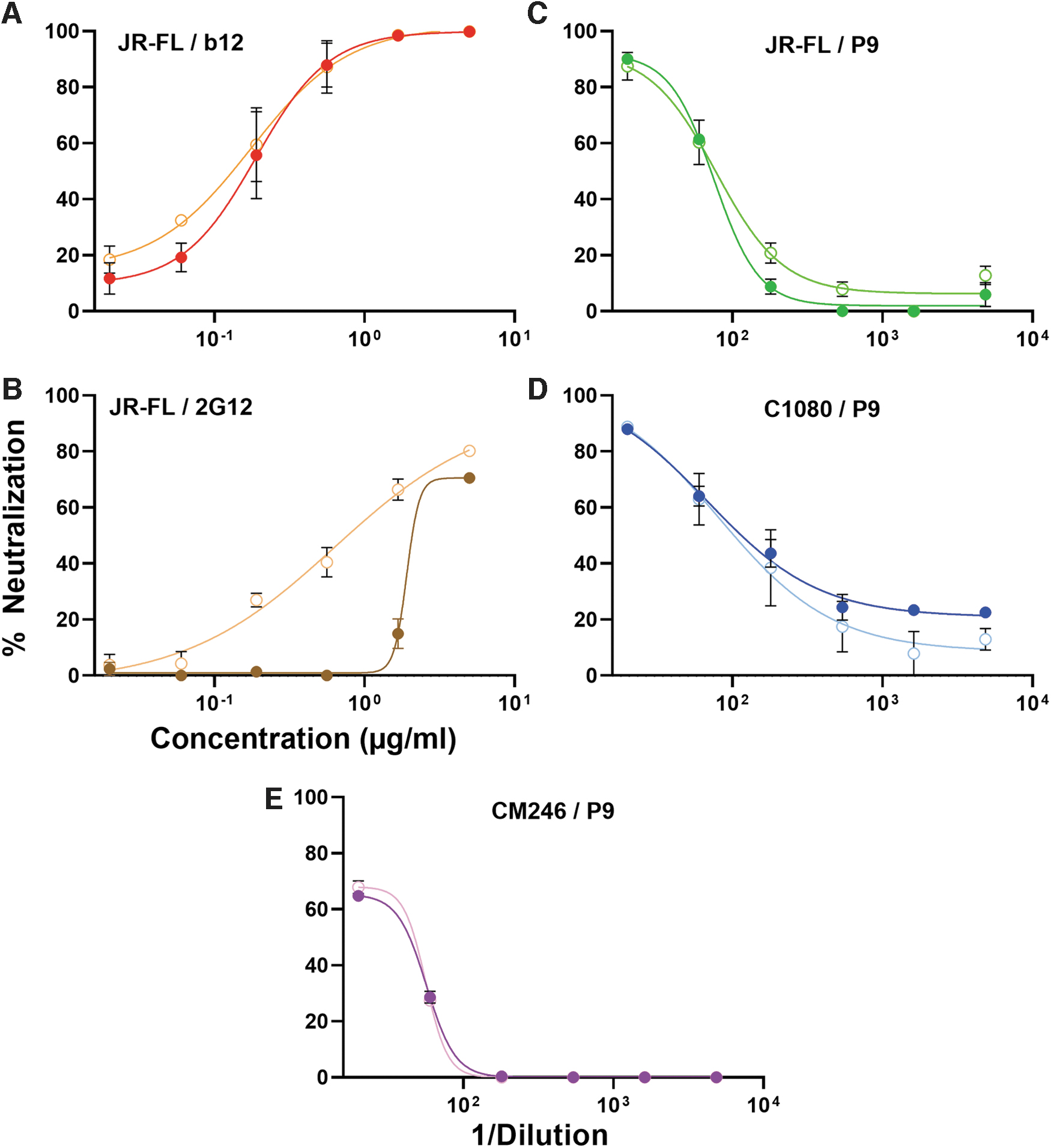
Neutralizing activities of P9 and mAbs with (filled circles) or without (open circles) added JR-FL gp120 D368R.
We next used a panel of mutants of the four sensitive strains to map its specificity (Fig. 3). These mutants were also evaluated with a panel of monoclonal bNAbs to monitor potential structural changes of the mutants. Mutations were introduced to either increase or decrease sensitivity to different specificities that might help reveal the focus of the P9 plasma. For JR-FL, D197N increases CD4bs NAb-resistance; E168K+/-N188A increases sensitivity to V2 bNAbs PG9, PG16, and PGT145; E168K+N362Q simultaneously improves sensitivity toward CD4bs b12, VRC01, and V2 bNAbs (Fig. 3). 8,45,49 The sensitivities of these mutations to 2G12 and PGT121 did not significantly differ from the JR-FL wild-type (WT) parent.
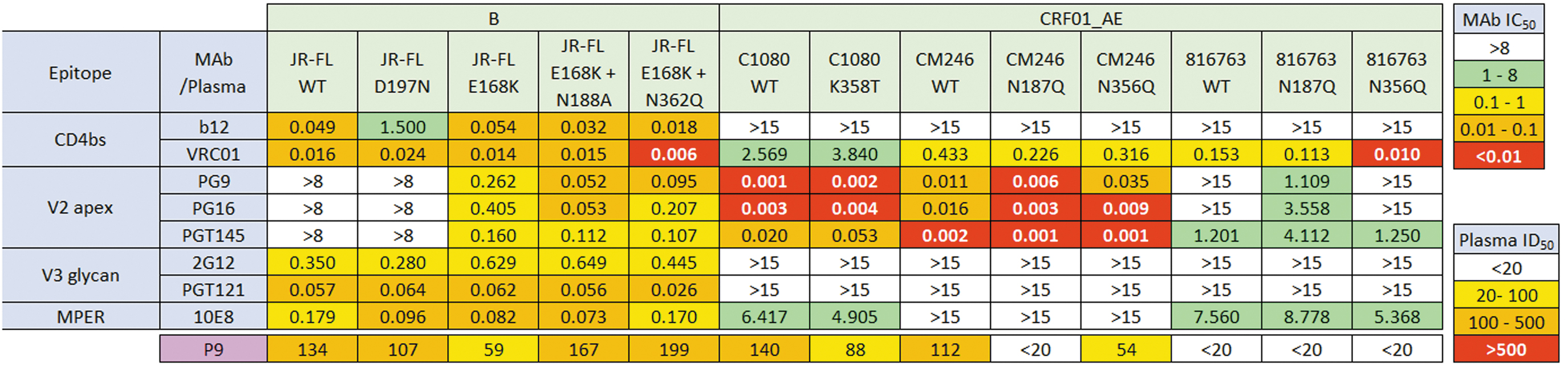
Minimum mean concentration (μg/mL) of mAbs (IC50) and dilution (ID50) of P9 plasma to neutralize WT and mutant PVs. IC50 (μg/mL) or ID50 (1/plasma dilution) are color coded with the lower the concentration or higher the dilution, the warmer the color. Assays were performed in duplicates and repeated at least twice. No 17b neutralizing activity was observed the across the subtypes and mutants (data not shown). IC50, inhibitory concentration at which 50% HIV-1 neutralization is attained; WT, wild type.
The fact that the JR-FL WT parent is insensitive to known V2 NAbs (e.g., PG9, PGT145), but is sensitive to the P9 plasma suggests that the P9 NAbs may not target the V2 apex (at least on the JR-FL). This point is underlined by the observation that E168K mutation reduces P9 sensitivity. However, overlaying a N188A glycan knockout mutation restored P9 sensitivity, perhaps by relieving glycan competition at the adjacent N188 and N189 sequons, with the one at N188 clashing with V2 NAbs and possibly the P9 plasma. In addition to improving V2 bNAb sensitivity, it is possible that N188 glycan knockout impacts sensitivity to other NAb specificities. Indeed, three lines of evidence suggest b12-like specificities in the P9 plasma. First, the N188A mutation slightly increased b12 sensitivity, similar to P9. Second, the D197N mutation modestly reduced both P9 and b12 NAb sensitivities. Third, N362 glycan knockout increased sensitivity to both P9 and b12. Like N197, the N362 glycan is part of a fence of glycans that surrounds the CD4bs and removing these glycans can improve access to CD4bs NAbs. 49 –51 Possible CD4bs-focused NAb activity could also explain why gp120 D368R did not interfere with P9 neutralization (Fig. 2), as the D368R mutation knocks out binding by many CD4bs-directed NAbs as well as CD4 binding. However, this interpretation is complicated by trimer shift assays (Fig. 4). This assay looks at the possible interaction between antibodies and Env expressed on PV. When antibodies are bound to Env, there is an increase in molecular weight of Env/antibody complex. This protein band will resolve slower compared with Env alone (Fig. 4, red asterisks). This is also accompanied by the depletion of native Env trimers (Fig. 4, blue asterisks). Here, plasma P9 bound effectively to JR-FL trimers, regardless of D368R mutation, unlike b12 (Fig. 4). Taken together, it is therefore possible that P9 NAbs targets a CD4bs epitope in a trimer-specific and D368-independent manner.

Blue native PAGE trimer shifts of JR-FL SOS and JR-FL SOS D368R PVs. Respective PVs were incubated alone, or with negative plasma, plasma P9, mAbs b12, or 2G12. Blue asterisks on the left of a band indicate Env depletion and red asterisks on the left of a band indicate protein band shifts. b12 and 2G12 mAbs bind to JR-FL SOS Env and cause a protein band shift (Lane 4 and 5). The D368R mutation abolishes b12 recognition and thus no protein band shift was observed in JR-FL SOS D368R (compare Lane 4 and 9). 2G12 recognizes JR-FL SOS independent of the D368R mutation and thus causes a protein band shift (Lane 5 and 10). The plasma P9 caused depletions in both JR-FL SOS and its D368R mutant, indicating binding toward JR-FL unaffected by the D368R mutation (Lane 3 and 8). No shift was observed in the negative plasma control. PAGE, polyacrylamide gel electrophoresis; Env, envelope glycoprotein.
In this study, 2G12 and PGT121 antibodies were ineffective against the AE strains as they lack the N332 glycan. 5,7 An analysis of 795 HIV-1 sequences across subtypes of the Los Alamos National Laboratory (LANL) HIV sequence database revealed that 95% of AE strains were glycosylated at position 334 and only 2% at position 332. 52 This accounts for the poor 2G12 and the PGT bNAb neutralizing efficacies against AE isolates (Fig. 3). 8,53 –55
Neutralization sensitivity differences between the three AE strains were exploited by toggling key differences in amino acid sequences and glycosylation sites (Fig. 5). We focused more on glycans, given their size and known ability to regulate neutralization sensitivity. 56,57 Indeed, N186 and N187 glycan deletions were reported to sensitize AE Env to b12 neutralization. 55 Alignment of these 3 AE Envs revealed two missing glycosylation sites in C1080 at positions 187 and 356 that are present on both CM246 and 816763 (Fig. 5, purple box). We hypothesized that manipulating these glycan sites may reveal if plasma capable of cross-neutralizing such as P9 are influenced by glycans at these sites. Aligning 4,316 HIV AE sequences from the LANL HIV sequence database revealed that 43% bear the N187 glycan, whereas only 6% bear the N356 glycan. N187Q and N356Q mutants were generated for CM246 and 816763 to test if the removal of these glycans alters P9 neutralization efficacy. Conversely, a K358T glycan was also introduced to C1080 to test if the addition of a glycan at this position (N355), which neighbors N356, increases this strain's neutralization resistance.
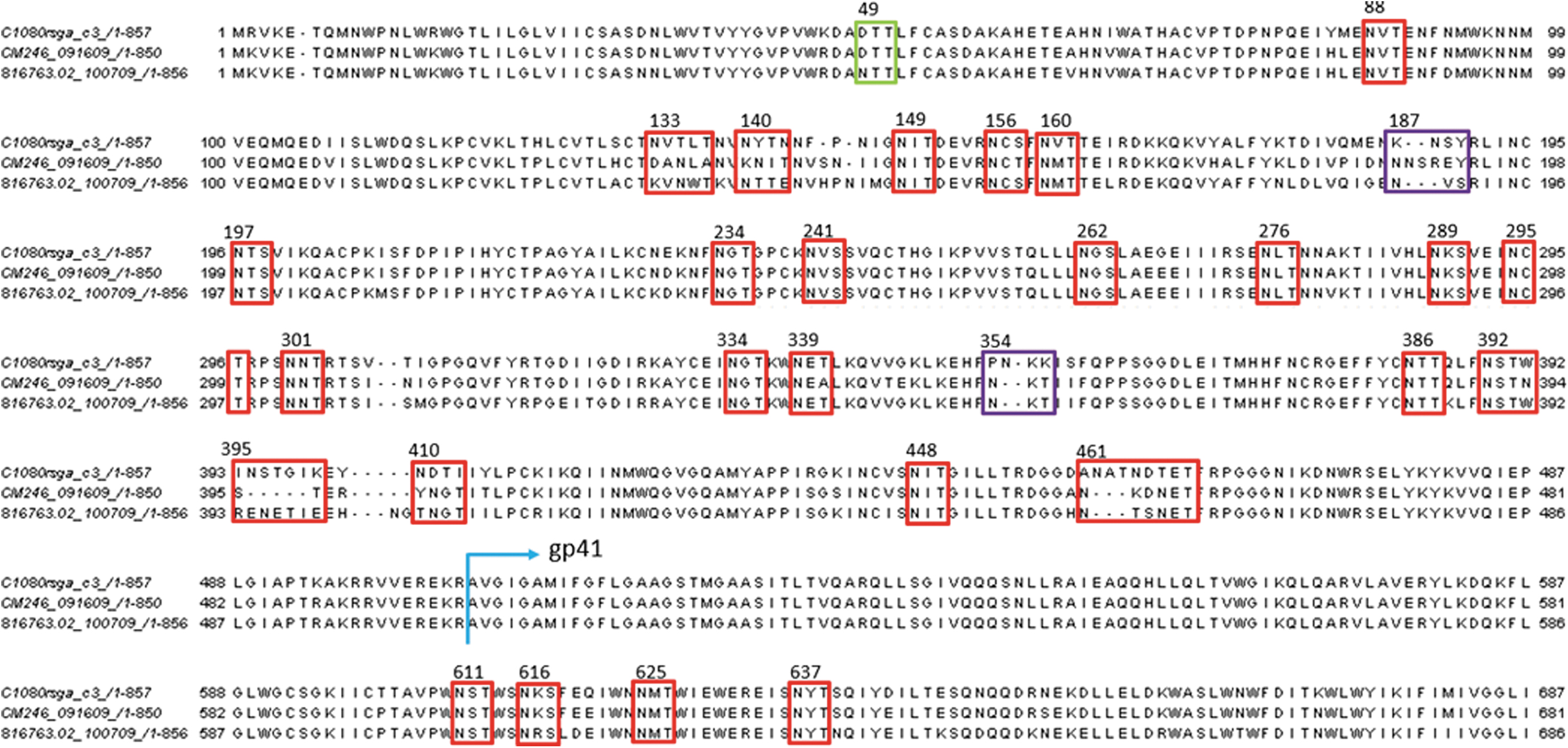
HIV-1 Env amino acid sequence of three AE strains (C1080, CM246, and 816763) are aligned using ClustalO. Amino acid numberings are based on HXB2 (accession no.: K03455). Blue arrow indicates where gp41 begins. PNGs are based on glycan sequon of NXT or NXS. Common and neighboring PNGs among the three strains were highlighted in red boxes. Key PNG differences that may contribute to neutralizing potency differences against the three strains were highlighted in purple boxes. The green box highlights 816763 unique glycan at N49. NXS, N is Asparagine, S is Serine, X is other amino acid except proline; NXT, N is Asparagine, T is Threonine, X is other amino acid except proline; PNG, potential N-glycosylation site.
We first checked the effects of these mutations against the monoclonal bNAb reference panel (Fig. 3). C1080 K358T mutant slightly increased VRC01 neutralization resistance, perhaps due to a role of the N355 glycan in lining the CD4bs glycan fence and its direct proximity to the CD4-binding loop (residues 361–380). 58 CM246 and 816763 WT PVs and their respective N187Q and N356Q mutants did not register significant changes in VRC01 sensitivity, except for 816763 N356Q, which increased its sensitivity to VRC01 by 10-fold. The C1080 and CM246 PVs, but not 816763 PV, were highly V2 Nab sensitive. Interestingly, the 816763 N187Q mutant knocks in PG9 and PG16 sensitivity, but this sensitivity increase was not observed against P9 plasma. This is similar to the effect of N188A in JR-FL that removes a clashing glycan. Conversely, 816763 N187Q was somewhat more PGT145 resistant. 59 In the context of the CM246 strain, N187Q led to improved sensitivities to all 3 V2 NAbs. Weak sensitivities to the MPER bNAb 10E8 were unchanged by these mutations, as expected.
We next tested the sensitivity of these mutants to the P9 plasma. The C1080 K358T mutant modestly reduced P9 sensitivity. This further supports the possibility of CD4bs NAbs in the P9 plasma (Fig. 3). However, the N356Q knockout mutation modestly reduced CM246 sensitivity and failed to knock in 816763 sensitivity. The N187Q mutants of both CM246 and 816763 PVs were P9 resistant.
Taken together, these data further support the possibility of trimer-specific D368-independent CD4bs NAbs in the P9 plasma and reduces the possibility that it targets the V2 apex or PG9-like antibodies. Our other plasmas (Fig. 1A) were also assayed against the AE glycan mutant PVs, but there were no noteworthy effects (Supplementary Fig. S2), suggesting that these glycans did not regulate sensitivity to these plasmas.
Discussion
In this study, we initially screened a modest panel of Malaysian plasmas for cross-neutralization and then tried to understand the specificities underlying the most cross-reactive plasma, P9. We narrowed down P9's most probable epitope target to be trimer specific and appearing to be CD4bs specific and not V2-specific or MPER-specific. Mapping a plasma's Env specificity is not a precise science and we face the same challenges as of similar previous mapping studies where the sera target was not ultimately definitive. Nonetheless, our study provided some useful hints to the epitope target of a cross-neutralizing Malaysian AE plasma and what could be expected in a larger cohort study in Malaysia.
Based on the inability of gp120 to interfere with P9 plasma neutralization, we initially speculated the presence of quaternary V2 bNAbs. 8 This was consistent with the observation that the AE strains, C1080 and CM246, but not 816763, were sensitive to the V2 bNAbs PG9 and PG16 as well as the P9 plasma (Figs. 1A and 3). However, P9 neutralization of JR-FL mutants contradicted this idea (Fig. 3). The E168K mutant knocks in V2 sensitivity that is absent in the parent JR-FL strain. Nevertheless, the JR-FL parent was sensitive to the P9 plasma. Coupled with the decreased sensitivity of the JR-FL E168K mutant, this suggests that P9 does not neutralize through V2 NAbs. Instead, several clues point to CD4bs specificity. First, lack of gp120 D368R interference could be explained by CD4bs NAbs as the D368R mutation knocks out a residue involved in binding by these NAbs. Second, toggling the presence or absence of glycans neighboring the CD4bs (N197 and N362 in JR-FL and N355 in C1080) impacted P9 activity in ways that partition with the general effects observed with these mutants using CD4bs monoclonal bNAbs, where the absence of these clashing glycans generally improved sensitivity and vice versa. Altogether, this provides evidence for CD4bs NAbs in P9 plasma with D368R independence and trimer dependence based on the gp120 interference and band shift assays.
Neutralization against mutant PVs suggested that the N187 and N356 glycans did not equivalently regulate P9 NAb sensitivity (Fig. 3). P9 plasma's reduced neutralization of the two CM246 mutants may be due to conformational effects rather than direct effects. It is possible that the N187 glycan structurally regulates access to the CD4bs, especially for b12-like CD4bs NAbs that target trimers in a tangential manner as compared with VRC01 class NAbs that typically avoid these clashes. 60 Mutating K358T in C1080, which introduces the missing N355 glycan, slightly increased its resistance toward P9, and similarly toward VRC01. The N355 or N356 glycan could contribute to the presentation of the neighboring CD4-binding loop for this observed slight difference in neutralization sensitivity. An N355Q mutant of CRF07_BC led to a slight increase in sensitivity to CD4bs NAbs b12, VRC03, and PG9, but no change for VRC01, and slightly reduced PG16 sensitivity. 61 This implies a role of N355/6 in protecting the CD4bs against some CD4bs NAbs by glycan clashes and may be a peripheral part of the CD4bs glycan fence. On the other hand, an N187Q mutant of an AE Env, 65CC4, was b12 resistant, 62 consistent with the effects of the N187Q mutants in our study (Fig. 3).
Looking at the mAb panel of Figure 3, the P9 plasma neutralized C1080, CM246, but not 816763, similar to the V2-specific bNAbs. It is possible that V2 bNAbs are present in P9 but without the same specificities and potencies as the PG-family bNAbs. From the band shift assay (Fig. 4), P9, was capable of binding to JR-FL SOS D368R to cause a depletion in the band and so potential presence of novel K168-independent V2 NAbs in P9 cannot be completely ruled out. This suggests recognition by P9 antibodies independent of D386R and they may not necessarily be CD4bs exclusively.
Limitations
Caveats of this project were the lack of clinical background information, limited plasma volumes, and lack of follow-up (longitudinal) samples. These plasmas were archived from donors obtained at least 6 years ago, so repeated draws from the same donors were difficult or impossible to obtain. Moreover, such a long gap is likely to result in NAb specificity changes in any surviving cases. More plasma volumes would facilitate comprehensive epitope mapping and allow next-generation neutralization fingerprinting on HIV-infected Malaysian plasmas. 63 –66 We considered that a neutralization assay panel of seven HIV-1 strains and an SIV control was a good trade off to sufficiently identify breadth and a few sensitive strains for mapping and conserving sufficient remaining plasma for further mapping work using a handful of mutants. It would be interesting to be able to study P9 further by comprehensively profiling their plasma NAb targets and discovering unique epitopes on AE Env. For example, double mutants (e.g., C1080 K187N + K358T or 816763 N187Q + N356Q) would have helped refine mapping and provide more robust conclusions.
Efforts to map plasmas are often clouded by the presence of multiple NAbs that may differ in specificity. 18,67 –69 As a result, plasma mapping is rarely definitive, and usually only provides hints. Thus, rescuing monoclonal bNAbs could greatly help to clarify the specificities that contribute to plasma neutralization. With this study, the specificities of AE-infected plasmas from Malaysia are not definitively clear and further investigations are required. Due to Malaysia's limited research infrastructure concerning HIV immunology evident by this study being the first locally, a starting foundation must first be established. The first step would be expanding the sample size to screen a larger panel of AE-infected Malaysian plasmas to provide a better overview on the NAbs elicited by this clade. In the present study, 14% of plasmas demonstrated breadth and potency, consistent with previous larger cohort studies. 70 –73 Moreover, the AE strains used for neutralization here originated in Thailand. There is yet to be a Malaysian AE representative Env for single cycle neutralization studies. Such Env clones would be useful to probe Malaysian AE plasmas against matched regional strains to better understand possible local neutralization serotypes. Autologous env genes isolated directly from neutralizing plasmas could be even more useful to understand bNAb origins and could even provide a basis for vaccine designs.
Conclusion
This study demonstrates that AE Envs can elicit NAbs in HIV-infected Malaysians with considerable breadth and potencies similar to larger cohort studies involving different clades around the world. Plasma P9 showed the greatest neutralization breadth, although its precise target could not be definitively determined. Nonetheless, epitope mapping suggested that P9's breadth and potency may be largely attributed to CD4bs NAbs, although with unusual features of trimer dependence and D368 independence.
Footnotes
Authors' Contributions
K.K.T., J.M.B., and T.T. conceived and designed the experiments; Q.R.N. performed the experiments; Q.R.N., J.M.B., and T.T. analyzed the data; K.K.T. and J.M.B. contributed reagents/materials and advice; Q.R.N., K.K.T., J.M.B., and T.T. wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Sunway University, Malaysia, grant no. INT-FST-DBS-2016-02.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.