
Abstract
Since emerging into the human population in late 2019, the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has reached across the globe to infect >80 million people. The coronavirus disease 2019 (COVID-19) caused by SARS-CoV-2 can range in severity from mild and asymptomatic to severe and fatal. Identifying risk factors for adverse outcomes in COVID-19 is a major challenge. In the context of the existing HIV-1 pandemic, whether COVID-19 disproportionately burdens people living with HIV-1 infection (PLWH) is unclear. The following discussion highlights pressing questions and challenges in the HIV-1 and SARS-CoV-2 syndemic, including (i) age, sex, and race as drivers of COVID-19 severity; (ii) whether chronic inflammation common in PLWH influences immune response; (iii) whether disease severity and trajectory models for COVID-19 ought to be calibrated for PLWH; (iv) vaccine considerations, and finally, (v) long-term health outcomes in PLWH that are further burdened by coinfection with SARS-CoV-2.
Introduction
Unbeknown to anyone at the time, the coronavirus disease 2019 (COVID-19) pandemic began in late December 2019 when a novel coronavirus was isolated in Wuhan, China and sequenced in early January 2020 (isolate Wuhan-Hu-1). 1 Over the next 2 months, highly related coronavirus isolates were identified in the United States (WA1 in Seattle, Washington; UC4 in San Francisco Bay Area, California; NY1-PVC8001 in New York City) and in Europe (BavPat1 in Munich, Germany). 2,3 Within the span of a few months (December 2019 to April 2020), several 1,000 severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) coronavirus isolates were identified, all sharing genetic relatedness to SARS-CoV, first identified in 2011, but distinct enough to form their own evolutionary cluster. Over this first year (December 2019 to December 2020) of the SARS-CoV-2 pandemic, >80 million people worldwide were infected, with global mortality totaling 2 million deaths. 4 By comparison, the HIV-1 pandemic has taken nearly 35 years (1984–2019) to infect an equivalent 80 million people worldwide, has sustained 30 million deaths in total, and 700,000 deaths in this past year alone. 5 Thus, at present, there is a syndemic of 40 million people living with HIV-1 infection (PLWH) and 100 million with SARS-CoV-2 infection worldwide. 4
In contrast with HIV-1 infection, which is nearly always fatal in the absence of potent antiretroviral therapy (ART), outcomes from SARS-CoV-2 infection are more heterogeneous, with ∼10%–20% of infections leading to severe clinical conditions and 1%–10% resulting in death within 2–3 weeks of infection. Although SARS-CoV-2 outcomes remain unpredictable for an individual, emerging research has identified populations at higher risk for severe disease outcomes once infected, including older age (>65 years old), 6,7 male sex, 8,9 and persons with preexisting conditions like cardiovascular disease and obesity. 6,10
Whereas combination antiretroviral therapy (cART) has had a positive impact on PLWH life expectancy, it is not curative and importantly does not restore full immunocompetence. 11,12 The extent to which PLWH experience adverse outcomes upon coinfection with SARS-CoV-2 is an active area of investigation. Initial results based on smaller cohort studies were inconclusive, but more recent and larger cohort studies suggest that COVID-19 mortality is higher in PLWH than in the general population. For example, a meta-analysis of five cohort studies 13 –17 indicated that overall COVID-19-related deaths in PLWH were nearly double that of persons without HIV-1 infection. 18 A study of 24,000 persons in South Africa reported that mortality rates were doubled in PLWH. 14 Similarly, a study of 17 million persons in the United Kingdom, including 27,000 PLWH, also showed a higher mortality rate in PLWH, especially among Black persons. 13 Explanatory factors may include biological and sociological factors such as restricted access to health care resources (needle exchange, counseling, and mental health services) and health delivery infrastructure (transit, continuity of care, adherence to antiretroviral therapy [ARV], and pre-exposure prophylaxis [PrEP] distribution) that resulted from public measures in response to COVID-19. 19,20
What follows is a discussion of challenges in the HIV-1 and SARS-CoV-2 syndemic, including (i) the role of age, sex, and race as drivers or risk factors for COVID-19; (ii) whether chronic inflammation common in PLWH influences response; (iii) whether models for severity and trajectory of COVID-19 may differ in PLWH; (iv) vaccine considerations; and finally, (v) long-term health outcomes in PLWH that are further burdened by coinfection with SARS-CoV-2 (Table 1).
Pressing Questions and Challenges in HIV-1 and SARS-CoV-2 Syndemic
ART, antiretroviral therapy; COVID-19, coronavirus disease 2019; PLWH, people living with HIV-1 infection; SARS-CoV-2, severe acute respiratory syndrome coronavirus-2.
Question 1: Are Biological Aging, Sex, and Race Drivers of COVID-19 Severity?
Role of biological age
Multiple studies indicate that age is a major risk factor for COVID-19 hospitalization and mortality. The Centers for Disease Control (CDC) estimate that the rate ratios of persons 50–85 years of age can be 4- to 8-fold higher for risk of hospitalization and 30- to 220-fold higher for risk of death compared with persons 18–29 years of age; and in persons 85 years or older, a 13-fold higher hospitalization risk and 630-fold higher death rate were observed. 21
Because PLWH are older than the general population, with <50% of PLWH older than the age of 50, compared with 34% of those older than 50 years in the general population, PLWH may be at increased risk for age-related COVID-19 complications. In the absence of SARS-CoV-2 infection, PLWH are already burdened by greater impairments in physical and cognitive function at ages younger than expected, 22 –25 higher levels of chronic immune activation and inflammation, 26 and multimorbidity compared with uninfected persons. 27 –29 The many recent advances in our understanding of basic mechanisms of biological aging 30 coupled with the emerging field of geroscience 31 should provide insights into the observed age-related decline in immune function in the general population 32 and in PLWH. 33,34 These advances should empower investigations into whether age-related complications in PLWH reflect accelerated, accentuated, or asynchronous biological aging and similarly, whether biological aging mechanisms are dysregulated in COVID-19. 35,36
Biomarkers for COVID-19 severity have been recently found to differ in association with age in a Boston cohort. 37 Whether COVID-19 impacts the severity of age-related comorbid conditions in PLWH remains unclear and will require longitudinal follow-up in older PLWH, with and without SARS-CoV-2 coinfection. Whether recovery may be slower in older PLWH or whether persistence of longer term COVID-19-related phenotypes will occur in PLWH will also needs to be determined to optimize medical care and ultimately healthspan. Finally, because age-related attenuation of vaccine response in PLWH has been observed with flu vaccination, 38,39 this may impact efficacy of prophylactic SARS-CoV-2 vaccination in PLWH (although Pfizer and Moderna data show no age-related decrease in vaccine efficacy in their studies in the general population).
Role of gender and sex hormones
Men between 40 and 70 years of age infected with SARS-CoV-2 tend to have more severe disease progression than women of the same age group 9,40 –44 and in some reports nearly twice the risk of death. 8,45 Determining the factors contributing to the higher risk for men has been hampered as data are often presented as aggregate outcomes without direct access to underlying demographic data. 45 Nevertheless, biological sex-specific risk factors likely include differences in immune response to infection, sex hormones as immunomodulatory effectors, sex-specific prevalence of comorbid conditions (e.g., hypertension and cardiovascular disease), and biobehavioral differences (e.g., smoking and drinking). 44 The Iwasaki lab recently identified male-associated increases in cytokine levels (e.g., IL-8 and IL-18) and more robust induction of nonclassical monocytes, whereas in women more robust CD8 T cell increases were observed. 46 Women also appear to display increased neutrophil activity and elevated expression of type 1 interferon-stimulated genes that enhance innate immune Toll-like receptor responses. 47
The primary cellular receptor and coreceptor for SARS-Co-V2 infection are the angiotensin-converting enzyme 2 (ACE2) and the transmembrane protease serine 2 (TMPRSS2), respectively. 48 ACE2 tends to be elevated in men compared with women and is associated with androgen expression levels. 49 Drug screening has also identified androgen signaling as a potential key modulator of ACE2 levels. 50 The coreceptor TMPRSS2 also appears to be androgen responsive. 51 Low testosterone levels in men are associated with more severe SARS-CoV-2 infection, 52,53 and estradiol levels were associated with interferon response. 53 In prior studies, low testosterone levels have been associated with elevated proinflammatory cytokines in men. 54,55 Androgen deficiency and dysregulation of testosterone activity through sex hormone-binding globulin occur frequently in men with HIV-1 infection, 56,57 raising the possibility that androgen dysregulation in the context of HIV-1 infection may impact ACE2/TMPRSS2 pathways. Although provocative, whether low testosterone levels are cause or consequence in COVID-19 severity in men, and whether indirect regulation of inflammatory and interferon response by sex hormones play a role in COVID-19 host response and disease severity remains unclear.
By contrast with COVID-19, among PLWH, women experience greater mortality than men 58 and women living with HIV have higher rates of multimorbidity than male PLWH. 59 In addition, women living with HIV progress to death at the same rate as men despite lower plasma HIV-RNA levels, suggesting different sex-associated risk profiles for PLWH. 60 Of note, despite equivalent ART-induced viral suppression, women living with HIV infection display less reduction in key markers of inflammation and immune activation compared with men. 61,62 Collectively, these observations highlight the need to define risk profiles and disease trajectories for men and women infected with HIV-1 and in those coinfected with SARS-CoV-2 to identify sex-specific and hormone-specific risk profiles for clinical outcomes based on gender and biological sex.
Role of race and ethnicity
Black and Latinx populations have disproportionately higher rates of SARS-CoV-2 infection, hospitalization, and COVID-19-related mortality, based on studies in the United States and the United Kingdom. 63 –65 The CDC estimates that Black and Latinx persons experience approximately twofold higher cases, fourfold higher hospitalizations, and threefold higher deaths, compared with non-Hispanic White persons. 66 In multivariable analysis of a large cohort in Louisiana, black race, increasing age, multimorbidity, public insurance (Medicare or Medicaid), residence in a low-income area, and obesity were factors associated with increased hospitalization. 64 Access to health care and community exposure to infection may also contribute to these disparities. 63,67 Once hospitalized, differences in mortality appear to become less significant, suggesting that structural determinants likely explain the disproportionate infections and deaths from COVID-19, 68 underscoring the need for improved and equitable health care access. 69
Among PLWH, similarly to COVID-19, Black and Latinx individuals have higher rates of HIV-1/AIDS-related morbidity and mortality compared with other racial and ethnic groups. 70 Increased disease burden is not restricted to PLWH, because non-HIV-infected Black and Latinx individuals also have higher rates of age-related functional decline and disability. 71 –73 Collectively, the differential risk for infection, hospitalization, and mortality may be owing to structural inequities, community, and workplace exposure and comorbid burden. Because Black and Latinx populations are more vulnerable than White populations to adverse outcomes from infection with SARS-CoV-2 and HIV-1, this should be a consideration when prioritizing populations for treatment, prophylaxis, and vaccination trials and rollout, as efficacious therapies and vaccines become available.
Question 2: Does Inflammation and Immunocompetence in PLWH Alter the Course of COVID-19?
Acute SARS-CoV-2 infection, when severe, can lead to an acute respiratory distress syndrome, 74 a respiratory condition characterized by rapid and excessive inflammation of the lungs, 75 with an imbalanced increase in inflammatory infiltrates including leukocytes, cytokines, and chemokines detectable in the lungs 76 and in peripheral blood. 77 –84 Levels of inflammation are dynamic over the course of disease, with changes in inflammatory levels related to the stage of disease 85 and viral burden. 86 Inflammatory drivers include infiltrating leukocytes, 87,88 with activation of RNA-sensing interferon and NF-κB-driven pathways, 80 and inflammasome and pyroptosis-mediated cell death and cytokine release. 89
Of note, inflammation is a hallmark driver of biological aging, 30,90 resulting in increased fatigue, declines in walking speed and increased disability, adversely affecting physical function and healthy aging. 90 –96 In the context of HIV-1, PLWH continue to have chronic low-grade inflammation compared with HIV-uninfected persons, despite undetectable viremia. For example, compared with the uninfected, PLWH on effective ART show persistent elevations in IL-6, TNF, sCD14, sCD163, CRP, and MCP-1 levels in peripheral blood. 26,97 –102 This observed HIV-associated inflammation is linked to adverse functional outcomes. 29,103 –105
Many questions remain unanswered. Does chronic inflammation in older PLWH increase susceptibility of PLWH to more severe outcomes with COVID-19? Does chronic inflammation accelerate increases in SARS-CoV-2 viral burden once PLWH are infected? Does chronic inflammation influence disease severity and progression, as well as subsequent time to resolution of SARS-CoV-2 infection? Because biological aging is associated with mechanistic changes in innate and adaptive immune signaling and immune response, 106 and chronic inflammation drives dysregulation between biological and chronological aging, 107 there is a need to better understand whether people aging with HIV-1 infection differ from the general population in their coordination of immune response, leukocyte composition, timing and dynamics of response, and cell signaling upon SARS-CoV-2 infection.
Clinical research efforts to effectively control COVID-19 will need a better understanding of risk factors for infection and, once infected, a more precise alignment between severity of disease and underlying immunophenotypes. Evidence to date indicate that risk factors influencing susceptibility to SARS-CoV-2 infection are multilayered and complex, ranging from cell and tissue expression of the viral target receptor/coreceptor pair (i.e., ACE2/TMPRSS2) to the environment (exposure within communities with higher SARS-CoV-2 prevalence, such as densely crowded spaces with prevalent infections, hospitals, or nursing homes). Clinical variability of COVID-19 represents a major challenge in risk assessment, with disease presentation ranging from asymptomatic infection with viral carriage possibly persisting to critical disease and death. Moderate and more severe COVID-19 can involve multisystem organ complications of the lungs, gut, heart, and brain. Efforts to characterize COVID-19 trajectory into stages begin with acute infection and inflammation, followed by a hyperinflammatory stage (2–5 weeks after symptom onset) distinct from the acute inflammatory phase and associated with coagulopathy and multisystem inflammatory syndromes in adults and children, and a late sequelae stage (4+ weeks postsymptom onset). 108,109 Operational categories for COVID-19 severity include a mild category (i.e., asymptomatic carriage of virus without manifestations of pneumonia on imaging), a moderate category (i.e., fever, respiratory symptoms, and radiologic findings of pneumonia), a severe category (i.e., respiratory distress, hypoxia, or abnormal blood gas), and a critical category (respiratory failure requiring mechanical ventilation and organ failure requiring intensive care). 85 How each of these stages in SARS-CoV-2 disease sequelae may differ in PLWH remains unclear (Fig. 1).
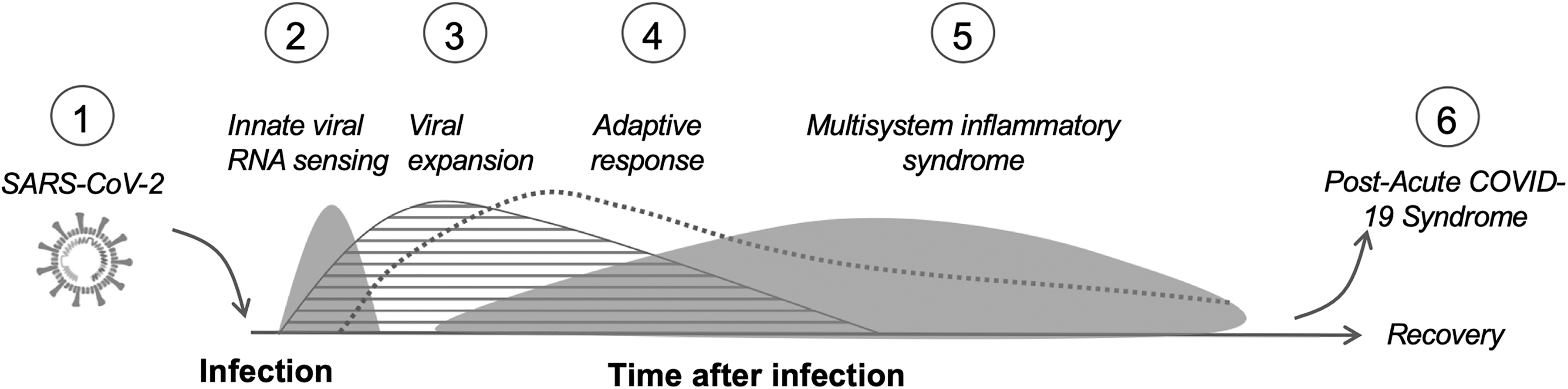
(1) SARS-CoV-2 infection of susceptible cells mediated by ACE2 and TMPRSS2. The infection of susceptible host cells by SARS-CoV-2. Potential dysregulation in sex hormone regulation and levels of inflammatory factors commonly observed in PLWH may impact SARS-CoV-2 receptor/coreceptor levels on susceptible cells, leukocyte trafficking at mucosal surfaces, and viral replication amplitude and duration once cells are infected. (2) Innate viral RNA sensing. Acute and innate viral RNA sensing in key cell subsets (e.g., macrophages) may be attenuated in PLWH, with aggravated activation of inflammatory factors and insufficient interferon type 1 responses, that phenocopy age-related response to other respiratory pathogens (e.g., flu). (3) Viral replication and expansion. The replication of SARS-CoV-2 depends, in part, on access to replication competent cells and host mechanisms limiting viral expansion. Leukocyte trafficking to mucosal sites, immune activation, and clearance of infected cells may be altered or impaired in PLWH. (4) Adaptive response. A coordinated response of innate and adaptive mechanisms necessary to control SARS-CoV-2 infection may be altered or attenuated in PLWH for whom immunocompetence is incomplete, despite effective ART. (5) Multisystem inflammatory syndrome. The chronic dysregulation of inflammatory pathways and multimorbidity that are common in PLWH may influence the composition and magnitude of inflammatory response, as well as time-to-resolution and recovery. (6) Postacute COVID-19 syndrome. Persistence of SARS-CoV-2 in mucosal sites (e.g., gut) may differ in PLWH because dysregulated mucosal immunity in the gut is common. In addition, chronic fatigue and cognitive deficits common in PLWH may be aggravated in postacute COVID-19 syndrome. ACE2, angiotensin-converting enzyme 2; ART, antiretroviral therapy; COVID-19, coronavirus disease 2019; PLWH, people living with HIV-1 infection; SARS-CoV-2, severe acute respiratory syndrome coronavirus-2; TMPRSS2, transmembrane protease serine 2.
The successful control of COVID-19 likely requires immune system coordination of innate responses to infection, 110 as well as adaptive SARS-CoV-2 antigen-specific antibody, CD4+ T cell, and CD8+ T cell immune responses. 84,111 In addition, innate immune response by monocyte subsets (e.g., expansion of intermediate monocytes and nonconventional monocytes) contribute to COVID-19 severity, 83,112 underscoring the need for coordination between innate and adaptive responses. 83,112 Dysregulated immune coordination may attenuate successful viral protection by neutralizing antibodies, considered a primary correlate of protection. 113,114 A wide range of SARS-CoV-2-specific neutralizing antibody titers have been reported after infection, 115 –120 with some reports showing antibody titer levels in asymptomatic persons that are, counterintuitively, lower than titer levels in moderate or severe COVID-19. 121 –124 Of note, the ratio and kinetics of SARS-CoV-2-specific anti-RBD, S1 and N antibody production and duration have been associated with disease severity. 125 A deeper understanding of antibody diversity, magnitude, timing, and duration is needed, 120 possibly through use of high-throughput serological profiling. 126
Age-related loss in coordinated immune response may also contribute to COVID-19 severity. 84 Drawing from earlier work on viral RNA sensing and subsequent signaling pathway analysis in flu infection of monocytes, interferon response, and inflammatory response can become uncoupled with aging, characterized by excessive inflammatory response and attenuated interferon response. 127,128 This uncoupling may also be relevant in SARS-CoV-2 infection. 129 Of note, older PLWH experience an exacerbated attenuation in response to flu vaccination. 38,39 These observations suggest that more insight is needed into the disease trajectory in PLWH compared with uninfected, as well as a deeper understanding of the potential compositional differences (i.e., innate and adaptive immunophenotypes) that underlie a successfully coordinated immune response to SARS-CoV-2 infection.
Question 3: Should We Adapt the COVID-19 Model for Disease Staging and Trajectory for PLWH?
Treatment approaches for COVID-19 are based on a disease model characterized by an early initial stage driven by viral replication and expansion, followed at later stages by coagulopathy and hyperinflammation. 85,108,109,112 Based on this model, antiviral (e.g., remdesivir) and antibody-based (bamlanivimab, casirivimab and imdevimab) treatments are initiated early in disease, 130,131 whereas immunomodulators and anti-inflammatory medications (e.g., dexamethasone, tocilizumab, and baricitinib) and anticoagulants (e.g., heparin) are used at later stages. 85,109
Whether one model of disease progression and trajectory sufficiently accounts for clinical variation associated with age, sex, race, and ethnicity, especially in the context of PLWH remains a major challenge. Transitions in disease from mild to moderate are likely to involve complex immunophenotypes 112 that may differ in timing, composition, and severity in PLWH. Information on disease stage when targeting treatment was highlighted in the RECOVERY trial findings of reduced mortality with dexamethasone treatment in severe or critical COVID-19 but not milder disease. 132 The benefit of selected immunomodulators may depend on better staging. Recent data from the REMAP-CAP trial support an overall benefit of tocilizumab, an IL-6 receptor antagonist, in critically ill COVID-19 patients, 133 although prior studies with tocilizumab showed mixed results, for example, benefit in retrospective analyses, 27 but not in prospective randomized controlled trials (RCTs). 134 Whether a selected immunomodulator is effective may therefore critically depend on the stage of disease, the dosage and timing of the intervention, as a more stratified approach to population selection.
Combinatorial treatment models are emerging that include the use of antiviral agents with immunomodulators. Recent combinatorial trials (e.g., the ACTT-2 trail that used the antiviral remdesivir in combination with baricitinib, an inhibitor of interferon JAK signaling 135 ) suggest that immune modulators and antivirals may be better than the use of an antiviral alone. However, considering that interferon-mediated antiviral response may be critical in the early stages of SARS-CoV-2 infection, and may be attenuated in older persons, and in PLWH, there will need to be clear biomarker profiles that distinguish the different disease stages to inform targeted intervention. For example, the immunosuppressive effects of baricitinib, while appearing useful later in disease, may be detrimental during early stages of infection by delaying viral clearance and increasing vulnerability to secondary opportunistic infections. 136
In the context of chronic inflammation and HIV infection, it remains unclear whether antiviral and/or immunomodulatory therapy should be initiated at a different time, follow a different timeline or dosage and selection of anti-inflammatory/immunomodulators be adapted in PLWH with COVID-19. Multiple circulating biomarkers are in use or proposed in COVID-19 disease risk assessment. 137 –139 Of note, however, many of these biomarkers are chronically dysregulated in PLWH, complicating diagnostics and treatment outcome assessment. Finally, given the polypharmacy in PLWH, drug interactions need to be considered when initiating treatment. Although there are some data on ART regimens and their use in COVID-19 (e.g., lopinavir–ritonavir), more research on short- and long-term outcomes with these and other ART medications with COVID-19-targeted treatments is needed. 140
Question 4: Will Assumptions About Vaccine Safety and Efficacy Hold Up in PLWH?
The development of vaccine candidates for SARS-CoV-2-driven COVID-19 has occurred at a profoundly accelerated pace. 124,141,142 However, from a historical perspective, smallpox is currently the only fully eradicated virus affecting the worldwide human population (e.g., polio is 99% eradicated but persists in some regions). 143 Therefore, eradicating SARS-CoV-2 represents a significant global challenge, especially given that the only known hosts for smallpox and polio are humans, whereas SARS-CoV-2 (and HIV-1) have nonhuman animal reservoirs, permitting zoonotic transmission (i.e., bats/pangolins/minks to humans for SARS-CoV-2, monkeys to humans for HIV-1).
Current vaccine designs include the mRNA genetic vaccines (e.g., Pfizer/BioNTech and Moderna/NIAID), nonreplicating adenoviral vectors (e.g., AstraZeneca/Oxford and J&J/BIDMC), viral proteins with an adjuvant (e.g., Novovax and Sanofi), and inactivated SARS-CoV-2 virus (e.g., Sinovac). 124,141,142 The COVID-19 vaccines based on mRNA technology were the first in the United States to obtain regulatory approval for use in humans, but multiple (over 200) other vaccines are now in trials worldwide. Because the human trials are primarily designed with clinical endpoints, rather than viremia endpoints, an important question is whether vaccines are permissive for asymptomatic carriage, allowing SARS-CoV-2 viremia to persist in the human population, including among vaccinated persons, although at reduced titers. Because persistence of SARS-CoV-2 in the human population is likely, a syndemic presence, along with HIV-1 must be considered. Sustained immunity once vaccinated and effectiveness against viral variants will also be challenges. Vaccine-induced memory, a key factor in protective immunity, will require B cell, CD8 T cell, and CD4 T cell coordinated response. Vaccine-elicited correlates of protection (e.g., neutralizing antibodies), viral clearance (e.g., T cell-mediated responses), and coordination (e.g., T cell-dependent B cell-mediated response) in the context of PLWH, remain to be fully characterized. The time frame for magnitude of response and subsequent decay in circulating B and T memory specific to SARS-CoV-2 appears to be 3–8 months postinfection in unvaccinated persons but is heterogeneous. 144,145 Whether the kinetics of decline and diversity of the memory compartments will differ in PLWH is also unknown but needs to be considered given incomplete immunocompetence in PLWH. 11,12
Although to a lesser extent than the HIV-1 pandemic, viral quasispecies diversity in SARS-CoV-2 represents an emerging challenge in identifying correlates of protection, 146,147 with an expansion of SARS-CoV-2 variants displaying altered phenotypes. 148 –150 Of importance, and in contrast with HIV-1, SARS-CoV-2 has a proof-reading function that limits, but does not eliminate, viral genetic diversity. The SARS-CoV-2 nonstructural protein 14 (nsp14) is evolutionarily conserved across coronaviruses and accompanies the viral replicase during RNA synthesis, excising mis-incorporated ribonucleotides that prevent error propagation. This error-correcting capacity limits diversity. 151 For example, in one study, a SARS-CoV-2 acutely infected patient had an average of one to four intrahost viral genetic variants detected. 152 By contrast, intrahost variants in acute HIV-1 infection can be many fold higher (e.g., 65 variants per person were identified in one recent study 153 ).
Although error-correcting capacity limits SARS-CoV-2 diversity, variants of concern (VOCs) are emerging with the potential for viral escape mutants (e.g., positions E484 and N501) 146,147 and altered phenotypes (e.g., D614G, 148 B.1.1.7, 149 and P.1150). Whether these VOCs and other emerging variants have differing transmission and pathogenic features in the context of prevalent HIV-1 infection is unknown and underscores the need for representation of PLWH in COVID-19 vaccine trials, surveillance studies, and in clinical outcome assessments. Given the lack of full immunocompetence among PLWH, the robustness and complexity of antibody response to vaccination, as well as potential differences in waning immunity and memory function, especially among older PLWH and in response to the evolving genetic landscape of VOCs should be considered.
Question 5: Postacute COVID-19 Syndrome and Long-Term Health Effects in PLWH?
As discussed, the clinical outcomes of acute COVID-19 can be heterogeneous and range from mild or asymptomatic in most cases (∼80%) to moderate and more severe disease in 10%–20% of those infected, with acute infection and clinical presentation occurring generally over the course of ∼3–4 weeks. Although most people survive SARS-CoV-2 infection, symptoms can persist over 60+ days after initial symptom onset, even despite the absence of detectable virus. 154 As of yet, there is no consensus definition of post-COVID-19 syndrome, thereby limiting estimates of prevalence. 155 Recovering persons variably experience symptoms, collectively referred to as “long or long-haul COVID-19” or postacute COVID-19 syndrome. Persisting conditions include substantial fatigue and dyspnea, and increasingly, there are reports of organ dysfunction in the heart, lungs, and brain. These persisting conditions may be owing to direct tissue invasion through ACE2 receptor-mediated infection, persistence of virus for several months in distal sites such as the gut, 156 and/or secondary effects of immunologic response to infection (e.g., leukocyte infiltration, production of autoantibodies, and tissue fibrosis). The duration and long-term health effects of those persons experiencing long-haul COVID-19 remain unclear.
Complications can be multisystemic in post-COVID-19 syndrome. 155 Cardiovascular complications can include myocardial inflammation (i.e., inflammatory infiltrates) and myocarditis (i.e., myocardium inflammation). For example, in one German study of 100 patients, 60% had myocardial inflammation 70+ days postinfection, based on MRI. 157 Whether cardiac cells are directly infected or whether myocardial inflammation is owing to leukocyte infiltration into heart tissue remains under investigation. Pulmonary dysfunction has been observed 3 months after hospital discharge, with evidence of pulmonary fibrosis. 158,159 An Austrian study found that lung damage was visible in >80% of patients 6 weeks after hospital discharge, and >50% at 12 weeks after hospital discharge. 160 Neurological manifestations of postacute COVID-19 syndrome include chemosensory dysfunction (smell/anosmia and taste/ageusia). In addition, “brain fog” reported 2–3 months after illness onset 161 may be related to cytokines and chemokines that cross the blood–brain barrier, affecting neurocognitive function. 162 Neurocognitive disorders might be triggered by direct brain damage from cerebral hypoxia or indirectly from immunologically induced neuroinflammation. 155 Relatedly, cognitive and psychosocial distress and loneliness may occur as a COVID-19-related stigma owing to the consequent need for physical distancing and isolation. 163 Finally, significant and persisting fatigue is often reported in post-COVID-19 syndrome, with >50% of individuals reporting fatigue 4 weeks after initial symptom onset. 154,164 Distinguishing COVID-19-related fatigue from age-related fatigue and from chronic fatigue syndrome/myalgic encephalomyelitis (i.e., CFS) is an active area of investigation.
Many clinics worldwide are now focusing efforts on post-COVID-19 syndrome to better define clinical phenotypes (e.g., clinics have been setup in the United Kingdom, in Boston, United States, and in Modena, Italy). Of note, the Post-Hospitalization COVID-19 Study (PHOSP-COVID) in the United Kingdom will collect data on 10,000 patients for a year, including data on various blood tests scans and biomarkers to identify diagnostic profiles. 165 Integrating PLWH into these clinical studies will be crucial to a better understanding in COVID-19 long-term impact in PLWH. Among aging PLWH, while living near-normal lifespans, multiple health burdens persist including chronic inflammation, multimorbidity (e.g., cardiovascular disease, cognitive impairment, fatigue, and frailty), 166 and polypharmacy. 28 Whether these health burdens in aging PLWH are exacerbated in those persons coinfected with SARS-CoV-2 or lower meaningful clinical thresholds for diagnostic complications associated with long-COVID-19 remains unclear.
Concluding Thoughts
Although great progress has been made in the understanding of SARS-CoV-2 and the disease it causes in humans, in the development of more effective therapies and most importantly in the development of effective vaccines, much work remains in our understanding of the disease in special populations including PLWH but also in ensuring equal access to care and treatment to all.
Internal Review Board Statement
This article is a review of published literature and publicly available data. No data are presented or described that can be linked to personal identity.
Footnotes
Author's Contribution
The author conceptualized and wrote the article in its entirety.
Acknowledgments
The author acknowledges the support of the Boston Claude D. Pepper Older Americans Independence Center and the Harvard University Center for AIDS Research.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The author acknowledges the following funding including P30 AG031679-10 6777 and P30AI060354-17 7331.