
Abstract
Microsporidiosis caused by Enterocytozoon bieneusi is a common opportunistic infection in patients with HIV/AIDS and those on immunosuppressive therapy. A significant loss of mucosal or peripheral CD4+ T cells and subsequent dysfunction of the intestinal immune system may be responsible for the development of chronic microsporidiosis in these patients. We have used the Simian immunodeficiency virus (SIV)-infected macaque model to investigate this relationship. To establish the course of E. bieneusi infection in SIV-infected animals, four SIV-infected animals were experimentally challenged with E. bieneusi when their CD4+ counts dropped to less than 300 cells/μL of blood. Analysis of fecal samples by nested polymerase chain reaction revealed that three out of four E. bieneusi-infected macaques continued to shed spores for 7–24 months after infection, an indication of chronic microsporidiosis. Four other SIV-infected macaques, after having an initial negative phase, spontaneously acquired E. bieneusi infection when their CD4+ counts dropped to less than 600 cells/μL of blood and shed spores for 8–19 months. The shedding of E. bieneusi spores in the feces increased relative to decrease in peripheral blood CD4+ T cell numbers. Gut biopsies were obtained before and after challenge to phenotype the mucosal lymphocyte subsets using flow cytometry. The immunophenotypic analysis showed no restoration of CD4+ T cells after E. bieneusi infection in the intestinal cells. A slight increase in the percentage population of CD4+ T cells in peripheral blood did not have any effect on the control of E. bieneusi infection in the SIV-infected macaques. These preliminary studies demonstrate that SIV-infected macaques develop chronic E. bieneusi infections as their CD4+ counts dropped to below 300 cells/μL of blood.
Introduction
E
The immunological status of the patient may determine whether an infection with microsporidia will be cleared or become persistent, without or with clinical manifestations of various degrees of severity of diarrhea. Studies have shown that persistent microsporidiosis is observed in HIV-infected patients when their CD4+ T lymphocytes dropped below 100 cells/mm3. 14 –16 It is difficult to study E. bieneusi in humans with HIV infection, as it is always confounded by variations in dietary intake and micronutrient supplementation, severity of HIV disease, and the complexities of multiple pathogens and therapies. Moreover, the progress on E. bieneusi studies has been slow, mainly because of the lack of in vitro cell culture and the in vivo animal models for parasite propagation for laboratory investigation are cumbersome. 17
The SIV-infected rhesus macaque model provides a tool to better understand this infection and the factors contributing to disease with the progression of HIV/AIDS. E. bieneusi has been successfully transmitted from AIDS patients to SIV-infected macaques 9,18 and to immunosuppressed gnotobiotic piglets. 19 However, in the pig model, the infection was very mild and the excretion of spores in the feces was sparse and intermittent. 19
SIVs are nonhuman primate lentiviruses that induce an acquired immunodeficiency with remarkable similarities to human HIV/AIDS. 20 The macaque SIV (SIVmac) model has been investigated extensively and has emerged as the premier animal model for AIDS. 21 –24 The clinical disease associated with SIV infection in rhesus macaques is similar in many ways to that observed with HIV in humans, although AIDS in macaques occurs in a considerably shorter time period. 25 As in humans, AIDS in macaques is characterized by progressive loss of CD4+ T lymphocytes and the occurrence of spontaneous opportunistic infection. 26,27
The E. bieneusi obtained from macaques are indistinguishable from E. bieneusi of human origin, morphologically and antigenically. 28 Establishment of persistent E. bieneusi infection in SIV-infected macaques makes it an appropriate model to study the progressive immunological defects and the status at which the infection with microsporidia becomes persistent and possibly symptomatic. In this study, we established and characterized the SIV-infected rhesus macaque as a model to investigate E. bieneusi infection in relation to immunodeficiency due to SIV/AIDS.
Materials and Methods
Animals
Ten SIV-infected rhesus macaques (Macaca mulatta) were used for studying E. bieneusi infection. All macaques were experimentally challenged, intravenously, with the SIV, SIVmac239, using 50 ng of p27 viral-antigen equivalent. Peripheral blood lymphocytes (PBLs) were collected monthly from each animal to monitor their peripheral blood CD4+ counts. Two SIV-infected macaques succumbed to SIV/AIDS-related complications before challenge with E. bieneusi spores. Four of the remaining eight SIV-infected macaques were coinfected with E. bieneusi when their peripheral blood CD4+ counts dropped <300 cells/μL of blood. The E. bieneusi used for these experiments was of human origin orally fed (∼105 spores each) to two SIV-infected rhesus macaques. 18 It was subsequently propagated in rats and orally fed to macaques at ∼105 spores per animal. 17 The genotype was not determined. The remaining four SIV-infected macaques acquired natural E. bieneusi infection. Clinical health of all the animals was recorded daily. Monthly fecal samples were obtained from SIV-infected animals to determine the presence of E. bieneusi infection.
After E. bieneusi infection of SIV-infected animals, fecal samples were obtained on a daily basis for the first 3 weeks and then three times a week from each infected animal. Fecal samples from SIV-naive animals were included as control. The level of E. bieneusi spore shedding was determined by both primary and nested polymerase chain reaction (PCR).
Animals were housed at the New England Regional Primate Research Center and were maintained in a centralized biosecurity level 3 animal-containment facility, in accordance with the Guide for the Care and Use of Laboratory Animals. Clinical procedures, administration of appropriate anesthesia and analgesics were performed under the direction of a veterinarian. If the veterinary staff considered it to be necessary, rhesus macaques were euthanized in accordance with the recommendations of the American Veterinary Medical Association Panel on Euthanasia. All animals received a complete necropsy by a veterinary pathologist after euthanasia, and tissue sections from the liver, spleen, kidneys, stomach, small intestine, cecum, and colon were collected for histopathological examination. All procedures and protocols were approved by the Institutional Animal Care and Use Committee at Tufts University and The Harvard Medical Area Standing Committee on Animals.
DNA extraction and PCR
A modified procedure of DNA extraction from fecal and bile samples was used as described previously. 29,30 Briefly, 200 μL of 0.5 mm glass beads (BioSpec Products, Bartlesville, OK), 400 μL of digestion buffer (100 mM sodium chloride, 25 mM ethylenediaminetetraacetic acid, 10 mM Tris [pH 8.0], 2% sodium dodecyl sulfate), 600 μL of Tris-buffered phenol:chloroform (BioExpress, Kaysville, UT) and 200 mg of feces was added to a 2 mL screw cap tube. The sample was homogenized by the Mini bead-beater (BioSpec Products) at 5,000 rpm for 2 min. After centrifugation at the top speed for 5 min, 400 μL of aqueous phase was adjusted to 0.7 M sodium chloride. The DNA was extracted using the GENECLEAN III Kit (Bio101; Carlsbad, CA) according to the manufacturer's instructions and stored at −20°C.
The first round of PCR (primary PCR) was performed with 1 μL of the DNA preparation described above with primers specific for the E. bieneusi ribosomal internal transcribed spacer (ITS) as described previously. 31 The nested PCR was performed with 1 μL of the product from the primary PCR with primers specific for E. bieneusi ITS-DNA. 31 The size of the product generated with outer primers (primary PCR) was 435 bp and the size of the product generated with nested primers was 390 bp. 30 The PCR products were visualized by the use of ethidium bromide staining after electrophoretic separation in 1.5% agarose gels. Based on PCR analysis, fecal samples were divided into two groups, positive for nested PCR or negative for nested PCR.
Isolation of lymphocytes and immunophenotypic analysis by flow cytometer
Endoscopic biopsies of jejunum and colon samples were obtained before and 2 weeks, 8 weeks, and then monthly after E. bieneusi inoculation of SIV-infected macaques for collection of intestinal lymphocytes. Intestinal lymphocytes (intraepithelial lymphocytes [IEL] and lamina propria lymphocytes [LPL]) were isolated from biopsy samples as described elsewhere.
32
Briefly, tissue samples were rinsed in cold phosphate-buffered saline (PBS) and placed in IEL isolation medium (Hanks balanced salt solution supplemented with 1 mM ethylenediaminetetraacetic acid, 100 U of penicillin per mL, 100 U of streptomycin per mL, and 5% fetal calf serum, pH 7.2) and subjected to rapid shaking at 37°C for 30 min. This procedure was repeated twice. Subsequently, LPL was isolated by incubating tissue samples in LPL isolation medium (RPMI-1640 medium containing 50 U of collagenase (Type II; Sigma), 100 U of penicillin, 100 U of streptomycin per mL, 5 mL
Lymphocytes were found to band at the interface between the 35% and 60% gradients. Isolated lymphocytes were used for staining with mouse anti-human monoclonal antibodies to CD3, CD4, CD8, CD2, CD20, and CD69. Whole blood samples were used for immunophenotyping PBL. Data were collected by using a Vantage flow cytometer and analyzed by CellQuest software (Becton Dickinson).
Immunophenotyping
Cell immunophenotyping was performed to determine peripheral blood and intestinal CD4+ and CD8+ T cell numbers. Cells were stained in the presence of staining media (PBS with 2% mouse serum). After antibody staining, the cells were fixed with fresh 2% paraformaldehyde. Three-color flow cytometry analysis of the cells was performed by use of a FACScan (Becton Dickinson) and CellQuest Software (version 3.2; Becton Dickinson). Appropriate isotype controls were used to establish positive and negative gates. Twenty thousand events were collected from a live gate to exclude cellular debris.
Results
E. bieneusi chronically infects SIV+ macaques
Four macaques were experimentally challenged with E. bieneusi spores when their CD4+ lymphocytes dropped to less than 300 cells/μL of peripheral blood, which varied from 25 to 67 weeks after SIV infection (Table 1). All fecal and bile samples from infected animals were tested for E. bieneusi by both primary and nested PCR, and specificity of the PCR product generated by nested PCRs was confirmed by restricted fragment length polymorphism (RFLP) technique that utilized MspA1I restriction endonuclease and resulted in expected fragmentation of amplified DNA. The result showed that E. bieneusi infection is localized in the feces and bile samples of the infected macaques as detected by primary PCR (Fig. 1A) and RFLP by using MspA1I restriction enzyme digestion (Fig. 1B). An E. bieneusi-specific primary PCR yielded production of 435 bp, and MspA1I restriction enzyme digestion yielded the expected DNA products of 258 and 92 bp (Fig. 1).
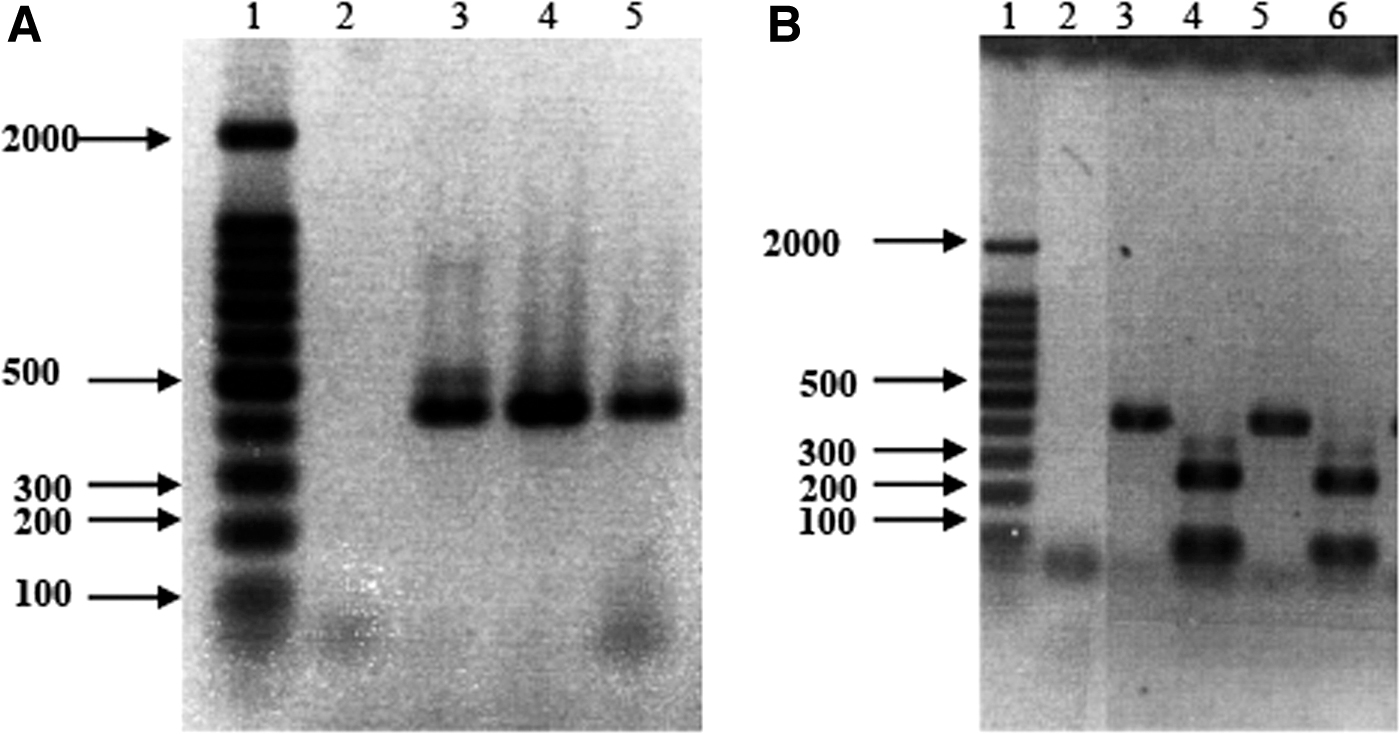
PCR and RFLP analysis of Enterocytozoon bieneusi DNA amplified in macaque fecal and bile samples.
Enterocytozoon bieneusi Infection Outcomes in Experimentally and Naturally Infected Simian Immunodeficiency Virus+ Macaques
Timing of Enterocytozoon bieneusi infection after SIV infection.
Duration of E. bieneusi shedding.
Time of euthanasia after E. bieneusi infection.
Spontaneous, natural infection occurred in four animals once their CD4+ T cells dropped <600 cells/μL of blood.
SIV, simian immunodeficiency virus.
Three of four SIV-infected macaques challenged with E. bieneusi developed chronic microsporidiosis as the infection became persistent and the shedding of E. bieneusi spores continued until euthanasia, which varied from 43 to 82 weeks postinfection (Table 1). One of the four challenged macaques did not develop persistent E. bieneusi infection and succumbed to SIV/AIDS requiring euthanasia at 4.5 months (Table 1). Similarly, four naturally infected SIV macaques, after having an initial negative phase, spontaneously acquired E. bieneusi infections when their CD4+ T cells dropped to less than 600 cells/μL of blood and showed persistent infection throughout the course of the study (Table 1). Three of the four experimentally infected and three of four naturally infected macaques developed clinical signs of anorexia and wasting, but only one of the E. bieneusi-infected animals developed diarrhea during the terminal stage of SIV infection. Thus, the results indicate that chronic symptomatic microsporidiosis can be established in SIV-infected macaques during the chronic stage of SIV infection.
Gross and histological examination of the tissues of all SIV-infected animals was completed after euthanasia (Table 2). In brief, histological examination of the SIV-infected macaques with chronic microsporidiosis revealed that the submucosa and lamina propria of the duodenum, cecum, and colon have multifocal expansion caused by marked infiltrates of lymphocytes, plasma cells, giant cells, and eosinophils. In the lumen, abundant protozoal organisms were also observed, and E. bieneusi was visualized in the lumen of the gastrointestinal tract and in the gallbladder using immunohistochemical staining (Fig. 2).
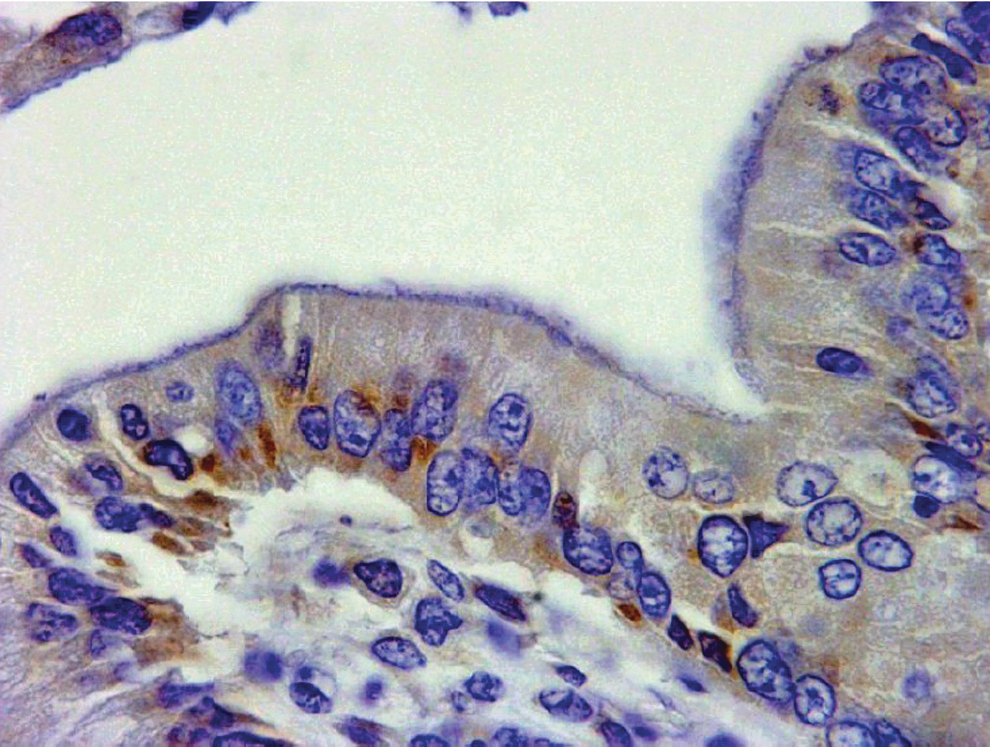
Immunohistochemistry staining for Enterocytozoon bieneusi. Staining of coinfected SIV-macaque, as seen in the mucosa of the gallbladder section of experimentally infected animal no. 3 (Table 1). The section was labeled with specific E. bieneusi monoclonal antibody counterstained with goat anti-mouse-labeled reagent. SIV, simian immunodeficiency virus.
Gross and Histological Observations in Three Macaques Coinfected with Enterocytozoon bieneusi and Simian Immunodeficiency Virus, and of Three Simian Immunodeficiency Virus-Infected Macaques Naturally Exposed with Enterocytozoon bieneusi
Macaques were experimentally infected when their CD4+ T cells dropped <300 cells/μL of blood.
Spontaneous, natural infection occurred in four animals once their CD4+ T cells dropped <600 cells/μL of blood.
SIV+ macaques develop anti-E. bieneusi serum antibodies
The presence of E. bieneusi-specific antibodies was determined in the serum samples taken from SIV+, E. bieneusi-infected macaques, which had chronic microsporidiosis evidenced by consistently high numbers of E. bieneusi spore shedding in the feces, as described above. E. bieneusi-specific antibody was present in the serum of chronically infected macaques, as detected by indirect immunofluorescence assay against purified E. bieneusi spores and compared with the serum from an E. bieneusi-negative animal (Fig. 3). The titer of antibody in the serum was low at 1/10.
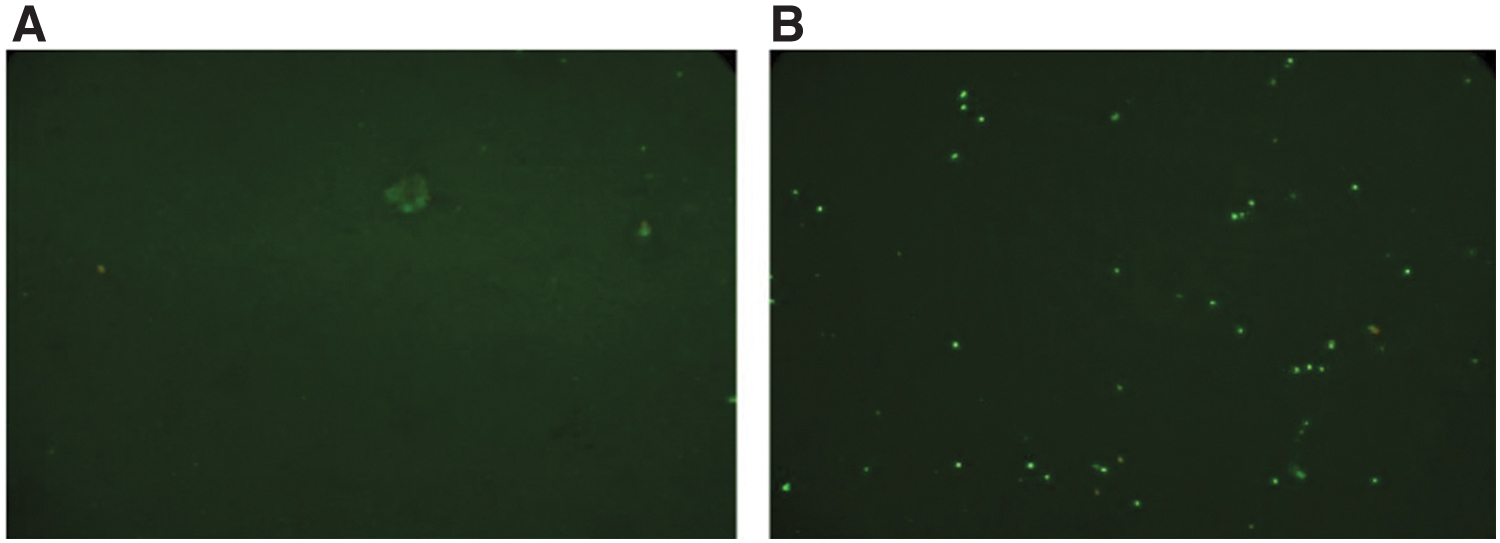
Detection of Enterocytozoon bieneusi-specific antibody in the serum of chronically infected, SIV+ macaque (no. 3; Table 1) by indirect immunofluorescence assay.
Immunophenotypic analysis of T cells in peripheral blood and intestinal tissues
Characterization of normal macaque peripheral blood and intestinal lymphocytes
The prevalence of different lymphocyte subsets was determined in the cell population isolated from peripheral blood, jejunum, and colon tissues by using three-color flow cytometric analysis (Table 3). In uninfected control animals (SIV naive), peripheral blood mononuclear cells (PBMCs) have more CD4+CD8− single-positive T cells than CD8+CD4− T cells (Table 3). Lymphocytes isolated from jejunum and colon biopsies of SIV-naive macaques are predominantly CD8+ T cells: 58% and 28%, respectively. Moreover, there was a predominance of activated CD4+ and CD8+ T cells in the intestine as compared with PBMC of normal macaques (data not shown).
The Mean (as Reflected in Table 1) of T Cell Subsets Obtained from Peripheral Blood, Jejunum, and Colon of Four Naive, Three Simian Immunodeficiency Virus-Infected, and Three Simian Immunodeficiency Virus+ Enterocytozoon bieneusi-Infected Macaques
SIV+Eb macaques were sampled at a time point indicated after Enterocytozoon bieneusi infection.
No restoration of CD4+ and CD8+ T cells in the intestinal mucosa and peripheral blood was detectable after E. bieneusi infection of SIV-infected macaques
After depletion of CD4+ T cells in the intestinal mucosa (23%–1.7%) and peripheral blood (30%–7.7%) after chronic SIV infection, no restoration of CD4+ T cells was observed after E. bieneusi infection of SIV-infected macaques at any stage of infection (Table 3). Although slight increases in the percentage populations of CD4+ and CD8+ T cells were observed at 3 and 4 months after E. bieneusi infection in the peripheral blood and intestinal tissues, this increase was not enough to resolve E. bieneusi infection in SIV-infected macaques. Similarly, there was an increase in the percentage positive population of CD8+ T cells in the intestinal tissues at 2 weeks and then at 4–6 months after E. bieneusi infection (Table 3), but again this increase had no effect on the recovery from infection. What constitutes protective immunity to E. bieneusi infection has not been elucidated.
Discussion
E. bieneusi is an important opportunistic infection causing chronic diarrhea, intestinal malabsorption, and wasting in the HIV-infected human population. Although the organism has been known since 198533 to cause chronic infection in this population, it has been poorly researched due to technical and logistical difficulties, including the lack of in vitro cultivation and poor and cumbersome propagation in animal models. 17 In addition, the length and variability of the incubation period, and the need for a severe immunosuppression either chemically or with dual infection, such as SIV for macaques, makes such studies very challenging. While the significance of microsporidiosis subsided considerably in the HIV/AIDS population in the developed world shortly after the successful introduction of antiretroviral therapy (ART), the disease continues to impact untreated AIDS patients, with the greatest impact in Sub-Saharan Africa, where it is commonly known as “Slim Disease.” The outcome of ART, however, has sadly had a dramatic effect on funding, which brought further investigations in the United States and elsewhere on this serious protozoal infection to a standstill.
E. bieneusi infection has a worldwide distribution and has been reported in almost all mammalian species examined. The infection in animals, as in healthy humans, is largely asymptomatic and tends to be persistent lasting for months and is localized mostly in the upper small intestine and in the hepatobiliary tract. The taxonomy and phylogeny are extremely complex, with many subtypes, and currently more than 90 E. bieneusi genotypes have been identified with considerable genetic diversity, entirely based on the ITS nucleotide sequence on spores recovered from the feces of humans and animals. Host-adapted E. bieneusi genotypes with narrow host ranges have been identified, as well as potentially zoonotic genotypes with a wide host specificity. 34 This study revealed that an immunodeficient primate can become chronically infected with E. bieneusi.
In this study, we show that SIV-infected macaques are a useful animal model to investigate some aspects of the relationship between the two infections, reflecting observations seen in human HIV infection. SIV-infected macaques developed chronic E. bieneusi infections with wasting when challenged with E. bieneusi spores when their blood CD4+ counts dropped to below 300 cells/μL, or after acquiring natural infection when their CD4+ lymphocyte counts dropped below 600 cells/μL of the blood.
We should add that while the work presented in this study is far from perfect due to costs, time involved and logistics, and that the field has moved away from the need of such investigations, it is unlikely that this or similar work will ever be repeated in the future.
Conclusions
In our experiments, seven of eight E. bieneusi-infected macaques developed chronic infection, as indicated by positive PCR of fecal and bile samples. All seven chronically infected animals developed anorexia and wasting, as is common in human HIV patients with chronic E. bieneusi infection. While only one of seven developed the characteristic diarrhea, all of the infected macaques had mucosal abnormalities of the intestinal tract. We performed immunophenotyping to characterize T cell subsets in both normal and SIV-infected macaques, and also characterized changes in T cells over time in chronically infected SIV and E. bieneusi-infected macaques. These analyses demonstrated no recovery of CD4+ T cells over time in the SIV+Eb-infected animals.
Footnotes
Acknowledgments
The authors would like to thank the animal attendants who provided care for these animals for prolonged periods of time, and to the pathology team who performed the necropsies and provided the histological and microbiological results.
Authors' Contributions
S.T.: Conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, resources, supervision, validation, writing—original draft preparation, and writing—review and editing. I.S.: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, validation, writing—original draft preparation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by NIH award—PO1DK55510 and RO1RR13537.