
Abstract
The biology of HIV-1 acquisition through unprotected receptive anal intercourse is understudied. Considering that sex hormones are implicated in intestinal physiology, pathology, and HIV acquisition and pathogenesis, we explored links between sex hormones, ex vivo HIV-1BaL infection of colonic mucosa, and candidate biomarkers of susceptibility to HIV-1 (CD4+ T cell frequencies and immune mediators) in cisgender women and men. No consistent significant associations between sex hormone concentrations and ex vivo tissue infection with HIV-1BaL were detected. In men, serum estradiol (E2) concentrations were positively associated with tissue proinflammatory mediators (IL17A, GM-CSF, IFNγ, TNFα, and MIG/CXCL9) and serum testosterone concentrations were negatively associated with frequencies of activated CD4+ T cells (CD4+CCR5+, CD4+HLA-DR+, and CD4+CD38+HLA-DR+). In women, the only significant interactions were positive associations between progesterone (P4)/E2 ratios and tissue ILRA concentrations and between P4/E2 ratios and frequencies of tissue CD4+α4β7high+ T cells. The study did not reveal relationships between biological sex or phase of the menstrual cycle and ex vivo tissue HIV-1BaL infection and tissue immune mediators. A comparison of CD4+ T cell frequencies between study groups revealed a higher frequency of tissue CD4+α4β7high+ T cells in women versus men. In contrast, higher frequencies of tissue CD4+CD103+ T cells were detected in men versus women in the follicular phase of the menstrual cycle. Overall, the study identified associations between systemic sex hormone concentrations, biological sex, and tissue candidate biomarkers of susceptibility to HIV-1. The significance of these results for tissue susceptibility to HIV-1 and early HIV-1 pathogenesis warrants further investigation.
Introduction
Despite the current evidence estimating a high risk of HIV acquisition during unprotected anal intercourse for the receptive partner, 1 an understanding of biological factors that can potentially modulate this risk is limited. Sex hormones play a complex role in gastrointestinal tract physiology and pathology and therefore it is plausible that fluctuations in sex hormone concentrations may change colonic mucosa susceptibility to HIV-1 infection. To our knowledge, there are no published data exploring relationships between systemic sex hormone concentrations and colonic tissue susceptibility to HIV-1.
Studies have documented involvement of sex hormones in the regulation of intestinal inflammation. For example, systemic concentrations of estradiol (E2) and testosterone are significantly lower in men and women with inflammatory bowel disease than in healthy control subjects. 2 In the colon, estradiol receptor (ER) signaling enhances expression of transmembrane tight junction proteins in noninflammatory conditions 3 and decreases production of proinflammatory cytokines in experimental colitis. 4,5
However, effects of exogenous estrogen seem to have a contrasting effect on colonic mucosa. Hormone replacement therapy (estrogen or estrogen+progestin) in postmenopausal women is associated with an increased risk of ulcerative colitis. 6 The risk appears to increase with increased duration of use and decrease with time since discontinuation. 6
In addition, an association between oral contraceptive pill use in premenopausal women and development of Crohn's disease was reported. 7 Testosterone was reported to be associated with positive outcomes of Crohn's disease. Men who received testosterone therapy demonstrated an improved clinical course of the disease and biochemical parameters, 8 possibly due to anti-inflammatory effects of the therapy. 9
We have previously demonstrated a negative association between serum E2 concentration and cervical tissue infection, following a high HIV-1BaL challenge dose ex vivo, and this was coincident with changes in several immunological parameters, including frequencies of CD4+α4β7+ T cells in the cervical mucosa. 10
Several immunological mechanisms of anti-HIV activity of E2 in different experimental systems have been suggested, including modulation of the β-catenin pathway and induction of IFNα. 11 –13 However, in addition to anti-HIV activity, E2 can also promote infection by stimulating HIV-1 transcription, 14 further emphasizing the need to better understand these complex effects.
This study was designed to investigate potential links between systemic sex hormone concentrations [E2, progesterone (P4), and testosterone], ex vivo colonic tissue infection, and tissue candidate biomarkers of susceptibility to HIV-1 in cisgender women and men. We also analyzed effects of biological sex and the menstrual cycle phase on colonic tissue infection and candidate biomarkers.
Methods
Subjects
The study was approved by the Icahn School of Medicine at Mount Sinai (ISMMS) IRB (study ID#16-00038/GCO#16-0081) and the Population Council IRB. Subjects were enrolled after providing written informed consent.
A total of 33 healthy volunteers were recruited: 22 cisgender women (24–52 years old; average 36.9 ± 8.2 years) not using hormonal contraception and 11 cisgender men (26–69 years old; average 48.1 ± 12.6 years) (Table 1). Subjects were defined as “healthy” based on history, physical examination, and laboratory assessments, including a negative HIV-1 serological test.
Demographics of Subjects Included in the Study and Serum Hormone Concentrations
Hispanic and Non-Hispanic ethnicity.
E2, estradiol; P4, progesterone.
Peripheral blood samples and rectosigmoid biopsies were collected at the time of colonoscopy at ISMMS. Biopsies were collected from all 22 women and from 10 men. There was no evidence of inflammation based on endoscopic or histological examination of biopsies.
The menstrual cycle phase was determined based on the last menstrual period (LMP) in subjects with self-reported regular cycle. The follicular phase group included samples collected between LMP and day 14 of the cycle (n = 6). The luteal phase group included samples collected after day 14 of the cycle (n = 13). Three women had irregular cycles and the phase of the cycle could not be determined.
Rectosigmoid biopsies
Up to 30 biopsies (2 × 2 mm) were collected from each subject using standard cup biopsy forceps in RPMI on wet ice. Twenty-one biopsies were utilized for ex vivo HIV-1BaL challenge. Three biopsies were cultured unchallenged in parallel with challenged tissues. At least four biopsies were used for flow cytometry.
HIV-1BaL
HIV-1BaL was generated as previously described. 15 Briefly, CD8-depleted peripheral blood mononuclear cells were activated for 6 days with 20 U/mL interleukin (IL)-2 (NCI BRB Preclinical Repository, Frederick, MD), 10 nM retinoic acid (RA; Sigma-Aldrich, St. Louis, MO), and 50 ng/mL anti-CD3 mAbs (clone OKT3; e-Bioscience, San Diego, CA) in RPMI 1640 containing 10% fetal bovine serum (FBS; Life Technologies, Grand Island, NY) (cRPMI).
Activated cells were then challenged with a 103 50% tissue culture infectious dose (TCID50) of HIV-1BaL (ABI, Eldersburg, MD) per 106 cells and cultured for 7 days in the presence of RA and IL2. The TCID50 was determined in TZM-bl cells. A single HIV-1BaL stock (2.78 × 105 TCID50/mL) was used in the study.
Ex vivo HIV-1BaL challenge of colonic mucosa
Colonic biopsies were transported to the laboratory within an hour of collection and processed for the ex vivo challenge and culture, as described in the study by Fletcher et al. 16 with modifications. Biopsies were washed in Dulbecco's phosphate-buffered saline (Corning, Corning, NY) and challenged with HIV-1BaL. Individual biopsies were placed in complete Dulbecco's modified Eagle's medium [cDMEM; DMEM containing 10% charcoal/dextran-treated fetal bovine serum (CS-FBS; Atlanta Biologicals, Flowery Branch, GA), 2 mM L-glutamine (Thermo Fisher Gibco, Grand Island, NY), 100 U/mL penicillin and 100 μg/mL streptomycin (Corning), and 80 μg/mL gentamicin (Corning)] in a 96-well plate.
Individual biopsies were then challenged with 1, 5, 25, 125, 625, 3125, and 15625 TCID50/mL HIV-1BaL in 200 μL, three biopsies per challenge dose. Biopsies from two subjects in the beginning of the study were challenged with 1–3125 TCID50 HIV-1BaL. Unchallenged control biopsies and, where feasible, 10 μM of lamivudine (3TC) control were included.
After 2–4 h of challenge at 37°C in 5% CO2, the biopsies were washed, then placed on top of a 12-mm presoaked circular surgifoam raft (Ethicon, Bridgewater, NJ) in cDMEM (three biopsies per raft/well; one well per challenge). Day 0 (d0) sample (postwashout) and d3, 7, 11, and 14 supernatants were collected. Infection was monitored by HIV-1 gag quantitative reverse transcription-polymerase chain reaction (qRT-PCR).
HIV-1 gag qRT-PCR
The infection was monitored in the tissue culture medium samples using HIV-1 gag qRT-PCR [lower limit of quantification (LLOQ): 2,000 copies/mL] using the SYBR FAST One-Step qRT-PCR kit (Kapa Biosystems, Wilmington, MA). 15
The primers used included the forward primer 5′ GGT GCG AGA GCG TCA GTA TTA AG 3′ and reverse primer 5′ AGC TCC CTG CTT GCC CAT A 3′. Five microliters of the tissue culture medium sample was used per reaction and lysed at 95°C for 5 min, followed by 4°C for 5 min, before addition of the SYBR Green reaction mix.
Results were analyzed using the standard curve method, using pNL(AD8) (11346l NIH AIDS Research and Reference Reagent Program, Germantown, MD) and the ViiA7 Real-Time PCR System (Applied Biosystems, Carlsbad, CA). The program for the analysis was as follows: one cycle at 42°C for 5 min; one cycle at 95°C for 5 min; and 40 cycles at 95°C for 3 s and 60°C for 20 s; followed by generation of dissociation curves to verify the absence of nonspecific amplification. Any value below LLOQ was set to
Data from the ViiA7 software were examined using soft (SOFT) and cumulative (CUM) endpoint analyses of the infection levels based on HIV-1 gag copies/mL on days 3–14 of culture. The SOFT endpoint is the virus growth measurement at the last time point where the increase in virus concentration between two consecutive time points is greater than the square root of the sum of sequential changes in virus concentration for the entire assay.
SOFT represents an estimation of the infection level at the start of the stationary phase of virus growth, that is, after exponential growth has been achieved and before the onset of the decline phase. 17 CUM represents the sum of viral growth during culture. 18
Radioimmunoassay
Serum samples were stored at −80°C before radioimmunoassay (RIA). E2 and P4 concentrations were determined using the ImmuChem Double Antibody 125I RIA kits for 17β-estradiol (LLOQ 10 pg/mL) and progesterone (LLOQ 200 pg/mL) (MP Biomedicals, LLC, NY), and testosterone concentrations were determined using the Testosterone Double Antibody Kit (LLOQ 100 pg/mL) (MP Biomedicals).
Any value below LLOQ was set to
Luminex
The Human Cytokine Magnetic 25-Plex Panel Kit (Thermo Fisher Scientific, Waltham, MA) was used to determine the levels of immune mediators in colonic tissue culture media from unchallenged biopsies collected on day 3 of culture. The analytes measured were IL1RA, IL1β, IL2, IL2R, IL4, IL5, IL6, IL7, IL10, IL12, IL13, IL15, IL17A, IFNα, IFNγ, eotaxin, GM-CSF, TNFα, CCL2, CCL3, CCL4, CCL5, CXCL8, CXCL9, and CXCL10.
As per manufacturer's protocol, the assay diluent was mixed 1:1 with cDMEM for tissue culture samples.
Flow cytometry
PBMCs were isolated from heparinized blood using Ficoll-Paque density gradient centrifugation. Colonic biopsies were digested by shaking in Hanks' Balanced Salt Solution (free of calcium and magnesium) (Thermo Fisher) with EDTA (0.5 M, pH 8) (Thermo Fisher) and gentamicin (100 μg/mL; Corning) for 30 min at 37°C, followed by collagenase IV (1 mg/mL; Worthington Biochemical, Lakewood, NJ) and 5% CS-FBS in HBSS for 40 min at 37°C. Cells were passed through a 40-μm strainer.
PBMCs and mucosal cells were stained with the fixable viability stain (FVS 780)-APC-Cy7 (Live/Dead; BD) and anti-CD3-BUV395 (clone UCHT1; BD Biosciences, San Jose, CA), -CD4-BUV496 (SK3; BD), -ICAM-1-BV421 (HA58; BD), -CCR5-AF-700 (HEK/I/85a; BioLegend; San Diego, CA), -CD103-BB515 (Ber-Act8; BD), -HLA-DR-PE-CF594 (G46-6; BD), -α4β7-PE (NHP reagent resource), and -CD38-PE-Cy5 (HIT2; BioLegend).
Samples were acquired on a BD LSR II flow cytometer and analyzed using the FlowJo 8.8.6 software. The gating strategy is shown in Supplementary Figure S1.
Statistics
The Kruskal–Wallis test with Dunn's multiple comparisons (women in follicular and luteal phases of the cycle and men) and Mann–Whitney test (women and men) were used to compare log-transformed cytokine/chemokine (CC/CK) levels and CD4+ T cell frequencies.
Linear regression analysis was used to analyze relationships between hormonal concentrations and log-transformed CC/CK concentrations and CD4+ T cell frequencies. The Grubbs test for outliers was performed. To maintain an overall type 1 error rate of 0.05, Benjamini–Hochberg critical values were used to determine the statistical significance.
Log-normal generalized linear models were used for analysis of tissue infection with HIV-1BaL between the groups (men, women, and women in follicular or luteal phase of the cycle). Analyses of association of infection endpoints (SOFT and CUM) with serum hormone levels (P4, E2, testosterone, and P4/E2 ratio), adjusting for the challenge dose, were done using log-normal generalized linear mixed models with random intercepts for each subject. The analyses were done modeling individual hormone concentration data or modeling all hormone concentration data, adjusting for each hormone.
The analyses were performed using GraphPad Prism 5.0 and SAS 9.4.
Results
Analysis of colonic tissue infection with HIV-1BaL and tissue candidate biomarkers of infection (CC/CK and CD4+ T cell frequencies) relative to serum sex hormone concentrations
To explore links between serum sex hormones and tissue infection, colonic biopsies were challenged with HIV-1BaL (1-15625 TCID50) and hormone concentrations were measured in serum collected at the time of biopsy collection. Paired biopsy infection data and hormonal concentration data were available from all n = 22 women in the study and from n = 10 men. A portion of unchallenged biopsies was used for T cell phenotyping on the day of tissue collection and for analysis of secreted CCs/CKs following a 3-day culture.
The tissue infection level in each subject and hormonal measurements are included in Supplementary Figures S2–S4 and Table 1. We detected high variability in E2 and P4 concentrations within the same phase of the cycle (Table 1 and Supplementary Fig. S4). Mean E2 concentrations in the follicular phase of the cycle were higher than in the luteal phase. Several women in the luteal phase had low concentrations of both E2 and P4. As expected, challenge with increasing TCID50 resulted in an increased number of productively infected biopsies and increased SOFT and CUM values (p < .0001) (Table 2). 3TC inhibited the infection, as evident starting day 7 of culture. The peak of the infection/time point before the stationary phase of viral growth (SOFT endpoint) in most of the subjects was detected at days 7–14, with infection peaking earlier, following higher challenge doses, as expected.
Relationships Between Serum Hormone Concentrations and Colonic Biopsies of HIV-1BaL Infection in Men and Women
Type III test of fixed effects tests the null hypothesis that the group means for CUM or SOFT are equal across virus challenge doses and the null hypotheses that CUM or SOFT levels are not associated with other effects (hormone concentrations), using F-tests.
CUM, cumulative.
In men, a significant positive association between serum E2 concentrations and the CUM HIV-1BaL infection endpoint and a trend of positive association between serum E2 concentrations and the SOFT infection endpoint were detected when all hormone concentration data were modeled in the analysis, adjusting for each virus challenge dose (slope = 0.0498, p = .0314, CUM; slope = 0.04369, p = .0591, SOFT). In models adjusting for just the virus challenge dose and E2, a trend of similar association was observed with E2 (slope = 0.0442, p = .0566, CUM; slope = 0.0392, p = .0832, SOFT). In addition, a significant positive association between P4 concentrations and tissue infection was observed when modeling only P4 (slope = 0.00198, p = .0299, CUM; slope = 0.0019, p = .0291, SOFT); however, the results were not significant when modeling all hormonal concentrations and adjusting for the virus challenge dose (Table 2).
Considering that both approaches to data analyses are relevant and the analysis outcomes are different, the results are interpreted as trends. The age of men did not seem to contribute to the results as no associations between age and E2 or P4 concentration were observed (Supplementary Fig. S5). Serum hormone levels were not associated with tissue infection in women (Table 2).
Of 25 measurements of CC/CK levels, IL2 concentrations were below LLOQ. IL1β and IL15 had single positive values. In men, the analysis revealed positive associations between E2 and concentrations of five proinflammatory/regulatory CCs (IL17A, GM-CSF, IFNγ, and TNFα) and CK MIG/CXCL9 in tissues (Fig. 1A), although only the association between E2 and IFNγ had a p value (.0017) below the Benjamini–Hochberg critical value (.0020). No associations between E2 concentrations and frequencies of tissue T cells were detected (p > .05; not shown). Testosterone was the only hormone in men associated (negatively) with frequencies of tissue CCR5+, HLA-DR+, and HLA-DR+CD38+-expressing CD4+ T cells (Fig. 2A), although only the p value (.0062) for association between testosterone and HLA-DR was below the Benjamini–Hochberg critical value (.0071).
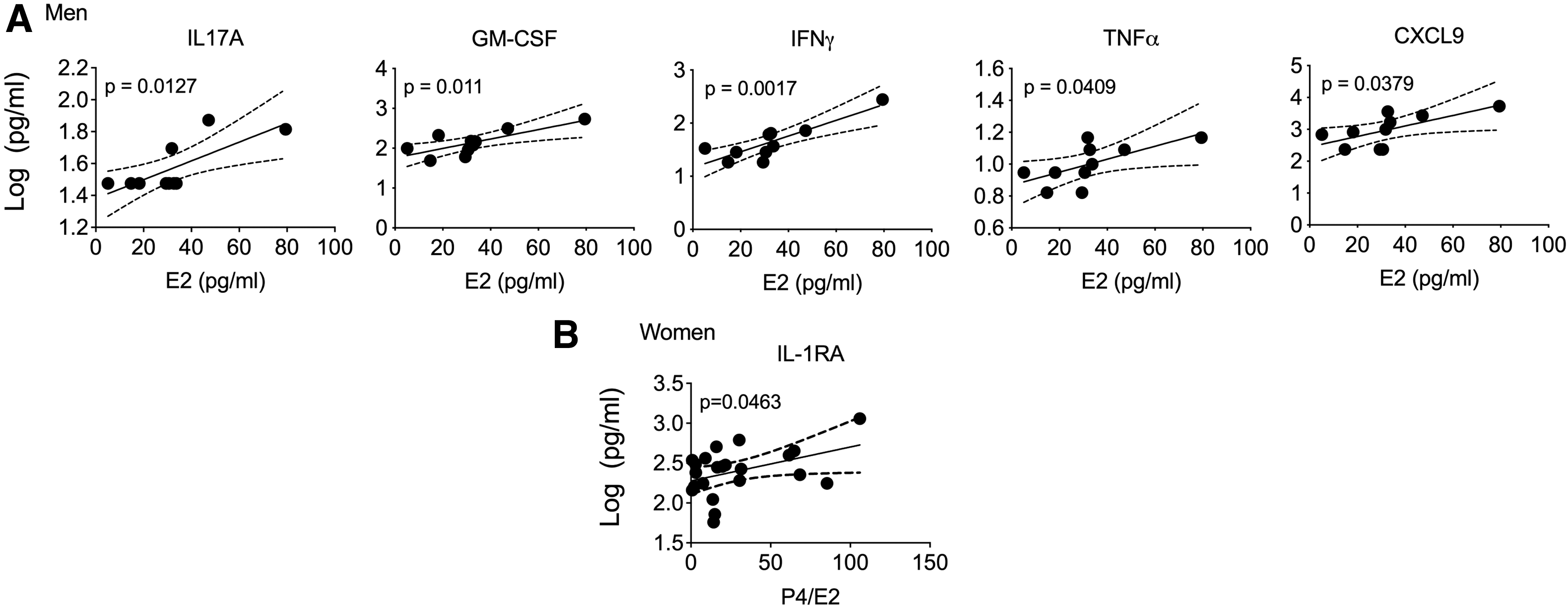
Serum hormones and CCs/CKs secreted by colonic tissue. Tissue culture supernatants from unchallenged colonic biopsies from men

Serum hormones and frequencies of activated colonic tissue CD4+ T cells. Frequencies of CD4+ T cells from men
We cannot exclude the contribution of subjects' age to these results as age negatively associated with testosterone levels (Supplementary Fig. S5).
In women, P4/E2 ratios positively associated with ILRA concentrations in tissues (Fig. 1B) and with frequencies of tissue CD4+α4β7high+ T cells (Fig. 2B), although the p values were not below the Benjamini–Hochberg critical value.
Relationships between biological sex, menstrual cycle phase, ex vivo HIV-1BaL infection of colonic tissue, and candidate biomarkers of infection (CC/CK and CD4+ T cell frequencies)
HIV-1BaL infection levels in colonic biopsies as well as CC/CK concentrations secreted by the tissue did not differ between men and women, irrespective of the phase of the cycle (Supplementary Figs. S6 and S7).
Analysis of the mucosal T cell phenotype revealed higher frequencies of α4β7high+ CD4+ T cells in women than in men. Women in the follicular phase of the cycle had lower frequencies of mucosal and blood CD4+CD103+ T cells than women in the luteal phase. Women in the follicular phase also had lower frequencies of mucosal CD4+CD103+ T cells than men. Frequencies of α4β7+, CCR5+, CD54+, HLA-DR+, CD38+, and HLA-DR+CD38+ CD4+ T cells were similar between the groups (Fig. 3).

Frequencies of colonic and blood T cells. Shown are frequencies of colonic tissue
Discussion
Our study did not reveal consistent significant associations between serum hormone concentrations and the ex vivo colonic tissue HIV-1BaL infection level in cisgender men and cisgender women. The observed trends of positive associations between E2, P4, and ex vivo tissue infection in men need to be confirmed in a larger study. Lack of association between E2 and tissue infection in women contrasts our earlier observation in cervical mucosa. 10 This is possibly due to differences in sex hormone effects at different mucosal sites and/or differences in experimental designs between the studies (analysis of individual ex vivo challenge doses vs. multiple ex vivo challenge doses). In addition, no effect of biological sex or menstrual cycle phase on ex vivo colonic tissue infection was noted.
Similar tissue infectivity data in men and women in our study are consistent with meta-analysis of the literature on HIV-1 infectiousness through unprotected receptive anal intercourse, showing no significant differences in per-act risks for heterosexuals and men who have sex with men (MSM). 1 However, the data are in contrast to a recent substudy of the HPTN 069/ACTG A5305 study by Sekabira et al., 19 which reported a higher infection level in colonic tissue derived from cisgender women (n = 11) than from MSM (n = 54) before and during oral pre-exposure prophylaxis.
Both our study and the study by Sekabira et al. employed the “exposed” edge model, allowing virus access to all surfaces. This model described by Fletcher et al. 16 possibly better reflects in vivo events where preparatory enemas and rectal intercourse are often traumatic to the epithelia, potentially affording direct HIV contact with subepithelial target cells. The discrepant results could partially be due to a smaller number of subjects in our study (n = 22 women and n = 10 men) and differences in subject characteristics.
In contrast to the report by Sekabira et al., women in our study did not use hormonal contraception. The small sample size in our study is partially offset by inclusion of six ex vivo challenge doses for each subject. This approach allowed us to present results across all challenge doses and minimize the effect of a single dose. Our study included older subjects with a broader age range compared with the study by Sekabira et al. and it is likely that older subjects with irregular menstrual cycles were perimenopausal.
We acknowledge variability in detected E2 and P4 levels between women in the same phase of the cycle in our study. As it was not feasible to recruit subjects within defined periods during follicular and luteal phases of the cycle, variability in E2 and P4 levels could be possibly attributed to samples collected during the periovulatory phase and in the beginning or in the end of the luteal phase. We also recognize that the menstrual cycle phase was assigned based on LMP in subjects with self-reported regular cycles. Self-report has limitations.
The tissue infection level in our study could have been impacted by potential differences in biopsy weights and number of HIV target cells, the parameters that are challenging to control. The colon has the largest numbers of immune cells in the body and most of those cells are activated. 20 Importantly, high correlations between weight-adjusted and nonweight-adjusted explant infection values for both cervical and rectal explants were reported. 19,21
Of note, we observed strong correlation between the HIV-1BaL challenge dose and tissue infection level in nonweight-adjusted tissues. Our HIV-1BaL stock was generated in the presence of RA, which could have enhanced infection in the tissues through RA-responsive elements in the LTR region and potentially contributed to equalizing infection levels between the groups. Future studies using transmitter–founder viruses will be important to further explore tissue susceptibility to HIV-1 between cisgender women and men.
Several associations between sex hormone concentrations and tissue candidate biomarkers of susceptibility to HIV-1 were observed in our study. E2 concentrations in men were positively associated with tissue-secreted proinflammatory CCs/CKs, and testosterone concentrations were negatively associated with frequencies of activated tissue CD4+ T cells. In women, the P4/E2 ratio was positively associated with tissue IL1RA concentrations and frequencies of tissue CD4+α4β7high+ T cells.
The results linking E2 and proinflammatory CCs, IL17A and IFNγ, in men in our study are consistent with data showing E2-induced increase in IFNγ secretion through ERα binding to the IFNγ promoter or through upregulation of the Th1 transcription factor, T-bet, 22 –25 as well as data showing that ERα signaling can increase IL17A production by Th17 cells. 26 The observed correlation between E2 and IL17A, GM-CSF, IFNγ, and TNFα points to CC profile characteristics of Th17 cells, which are highly susceptible to infection with HIV-1 and SIV. 27 –30
Importantly, IL17A is known to interact with GM-CSF, IFNγ, and TNFα to exert a proinflammatory effect that may augment tissue susceptibility to HIV infection. CXCL9 was previously shown to be induced during HIV infection of rectosigmoid tissue in vitro. 31 Further implicating the above CCs/CKs in HIV acquisition, stimulated rectal CD4+ T cells from subjects involved in condomless receptive anal intercourse showed high levels of IL17 production and stimulated CD8+ T cells had a high level of IFNγ and coexpression of IFNγ and TNFα. 32
The identified negative associations between the testosterone level in men and frequencies of CD4+ T cells expressing CCR5, HLA-DR, or HLA-DR and CD38 in the mucosa suggest a role for testosterone in controlling T cell activation. The positive association between P4/E2 ratios and frequencies of colonic CD4+α4β7high+ cells in women is consistent with similar association in cervical mucosa. 10
It is yet to be determined whether observed associations are consequential to colonic tissue susceptibility to HIV-1. Only p values for the associations between E2 and IFNγ and between testosterone and CD4+HLA-DR+ cells in men were below the threshold for rejection of the null hypothesis after multiple comparisons correction. Further investigation, in a larger study, would help confirm these and other observed associations.
This study demonstrated similar tissue-secreted CC/CK concentrations in women in follicular and luteal phases of the cycle and in men and revealed higher frequencies of tissue CD4+α4β7high+ cells in women than in men. In the context of similar tissue infectivity between the groups, the finding of higher frequencies of tissue CD4+α4β7high+ cells in women than in men is unexpected and emphasizes that the role of α4β7 in HIV transmission requires further studies, as recently reviewed by Pollock et al. 33
An increase in frequencies of CD103+-expressing blood and tissue CD4+ T cells was observed in subjects during the luteal versus follicular phase of the cycle. In addition, frequencies of tissue CD4+CD103+ T cells were higher in men than in women during the follicular phase. Our data demonstrating similar infection levels between these groups of subjects do not support the role of CD4+CD103+ cells during HIV infection in colonic mucosa. However, a potential proinflammatory role was previously reported for CD4+CD103+ cells in subjects with ulcerative colitis 34 and it may be of importance to further explore this finding in the context of tissue susceptibility to HIV.
Overall, the study identified associations between systemic sex hormone concentrations, biological sex, and tissue candidate biomarkers of susceptibility to HIV-1. The results warrant further investigation in larger cohorts of cisgender men and women and suggest the need to study relationships between hormonal therapy and colonic tissue susceptibility to HIV-1 in transgender subjects.
Footnotes
Acknowledgments
The authors thank Drs. Grace Aldrovandi, Peter Anton, and Julie Elliott for the data discussion.
Authors' Contributions
P.B. was involved in investigation, data curation, visualization, validation, and formal analysis. S.M. and L.K. were involved in investigation and data curation. N.K. was involved in supervision. M.P. was involved in formal analysis and software. S. Mehandru was involved in conceptualization, funding acquisition, resources, project administration, and writing—review and editing. N.T. was involved in conceptualization, funding acquisition, resources, project administration, validation, writing, and writing—review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was funded by grants from the National Institutes of Health, R01AI110370 (N. Teleshova), R01 DK112296 (S. Mehandru), and R01 DK123749 (S. Mehandru).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.