
Abstract
The persistence of low human immunodeficiency virus type 1 (HIV-1) replication in individuals undergoing antiretroviral therapy (ART) still threatens their health. Previous findings have shown that microRNAs (miRNAs) could interfere with several steps of the viral life cycle. Herein, we set out to investigate the expression of miR-150, miR-223, miR-382, miR-324-5p, miR-33a-5p, miR-34a, and miR-132 in the whole peripheral blood mononuclear cell (PBMC) population from people living with HIV-1 showing different levels of viral suppression. Levels of PBMC-associated miRNAs were analyzed in 30 individuals with undetectable viremia (target not detected) and 30 individuals with detectable low-level viremia (1–200 copies/mL). In addition, 30 samples from treatment-naive (NAIVE) individuals were investigated. Results were compared to a control group of 28 HIV-negative donors. All miRNAs analyzed were strongly downregulated in the NAIVE population, either compared to the treated group or to controls. Stratification of ART-treated donors according to the therapeutic regimen showed the downregulation of miR-33a-5p in subjects treated with non-nucleoside reverse transcriptase inhibitors compared with those treated with protease inhibitors. Collectively, the present study shows that uncontrolled viral replication leads to profound miRNA deregulation while treated individuals, irrespective of the degree of viral suppression, and even the types of antiviral drugs seem to be specifically associated with miRNA expression profiles. These evidences suggest that virological suppression could be favored by miRNA modulation.
Introduction
The control of the human immunodeficiency virus type 1 (HIV-1) infection and improvements in clinical outcomes have been attainable since the advent of antiretroviral therapy (ART) in clinical practice. Nevertheless, most treated individuals do not reach complete viral suppression in peripheral blood. The source of residual viremia is still not well established and most likely results from multiple mechanisms, such as ongoing rounds of viral replication and intermittent reactivation of latently infected cells. 1 The persistence of viral replication and the ensuing consequences, such as the increased risk of viral rebound, 2 –4 and the higher levels of inflammation markers, 5 remain challenging for clinical and basic researchers. During the HIV-1 life cycle, a complex virus-host interplay is established, and host factors are likely to play a critical role in the biological processes underlying HIV-1 replication.
MicroRNAs (miRNAs) are noncoding, single-stranded RNAs involved in post-transcriptional gene regulation by base-pairing with target messenger RNAs, leading to translation repression or transcript cleavage. 6 This mechanism of gene silencing, known as RNA interference (RNAi), 7,8 plays a pivotal role in various cell biological processes. 9
In addition to their role in cellular physiology, miRNAs are also involved in human viral disease; host miRNAs can positively or negatively affect the replication and pathogenesis of a broad range of viruses by either directly targeting pathogen untranslated regions (UTR), 10,11 RNA-based virus coding sequences 12 or by affecting host gene expression.
Likewise, findings on cellular miRNA-mediated regulation of HIV-1 disease progression are accumulating. 13 T cell activation is essential for productive HIV-1 infection, and an altered miRNA profile has been observed according to their differentiation and activation status. 14 For instance, activated CD4+ T cells isolated from HIV-negative donors (HD) showed high miR-132 levels, and its overexpression correlates with enhanced in vitro HIV-1 replication. 15 It has been shown that miR-34a and miR-217 may induce Tat activation by inhibiting the expression of the human sirtuin-1 (SIRT1) gene 16 –18 ; in support of this evidence, increased levels of miR-34a have been found in HIV-infected human T cells. 19
In contrast, RNAi machinery can inhibit HIV-1 replication. Previous studies revealed that miR-33a-5p and miR-29b-3p overexpression significantly decreased p24 levels in supernatants recovered from primary CD4+ T cells infected with HIV-1. 20 Besides, cellular miR-324-5p was predicted to counter the expression of the viral Vif gene, 21 thereby favoring APOBEC3G/F activity on viral cDNA that may lead to the establishment of a replication-defective reservoir.
The 3′ UTR of viral transcripts can be targeted by a cluster of miRNAs, including miR-150, miR-223, and miR-382. Since these miRNAs were found highly expressed in resting CD4+ T cells and their inhibition resulted in augmented viral particle production, their role in maintaining viral latency has been proposed 22 along with other miRNAs. 23 –25 Furthermore, the overexpression of miR-150, miR-223, and miR-382 in freshly isolated monocytes inhibits HIV-1 replication, suggesting their participation in the antiviral immune response. 26
In the last decade, many efforts have been made to characterize miRNA expression in HIV-positive (HIV+) individuals. Previous studies have revealed differential miRNA expression profiles by comparing samples from elite controller and long-term nonprogressor individuals to HIV-negative, untreated and treated HIV+ donors. 20,27,28
The potential use of miRNAs as a marker of disease progression has also been addressed. Munshi et al. 29 reported a positive association between levels of miR-150 and CD4+ cell counts, and the absolute quantities of miR-150 and miR-146b-5p, both in peripheral blood mononuclear cells (PBMCs) and plasma samples, may discriminate symptomatic treatment-naive (NAIVE) individuals from ART-treated and individuals harboring ART-resistant viruses. 29
These findings emphasize the role of host miRNAs during HIV-1 infection and their potential applications in clinical practice. However, their differential expression has never been investigated among people showing different viral loads, including those who achieved complete and sustained virological suppression. For this reason, we sought to investigate the expression of PBMC-associated miRNAs that have been experimentally proven to participate in viral replication and maintenance of viral latency to identify potential molecules responsible for controlling viral replication in treated HIV+ individuals.
Materials and Methods
Study population
Expression levels of miR-150, miR-223, miR-382, miR-33a-5p, miR-324-5p, miR-34a, and miR-132 were evaluated in unsorted PBMC samples collected from 90 people living with HIV. Sixty donors were on ART, and 30 were NAIVE. According to plasma viral load observed over the past 3 years, treated individuals were stratified into two groups: (1) 30 with undetectable viremia (TND, target not detected); (2) 30 with detectable low-level viremia (LLV, defined as 1–200 copies/mL). Twenty-eight volunteer health care workers were enrolled in the HD group.
The local Ethics Committee granted ethical approval for the study (protocol number 6312-0528/2021). Written informed consent for anonymous clinical and biological data was obtained from all individuals.
HIV RNA evaluation
HIV RNA was quantified using the kinetic polymerase chain reaction (PCR) molecular system (kPCR, Versant HIV-1 RNA kPCR 1.0; Siemens Healthcare Diagnostics, Tarrytown, NY). The kPCR assay's linearity range is between 37 and 11 × 106 copies/mL. Qualitative results below the lower limit of quantification were reported indicating the presence or absence of residual viremia.
PBMC separation
Isolation of PBMCs from whole blood was obtained by Ficoll-Hypaque density gradient centrifugation.
RNA extraction
Total RNA was isolated from ∼5 × 106 PBMCs using the mirVanaTM miRNA Isolation Kit (Life Technologies, Carlsbad, CA) according to the manufacturers' recommendation.
Reverse transcription PCR
The cDNA was synthesized from total RNA using a specific stem-loop reverse transcription (RT) primer supplied in the TaqMan® MicroRNA Reverse Transcription Kit (Applied Biosystem, Foster City, CA, USA). For each 15 μL RT reaction, 5 μL of RNA was combined with master mix solution prepared as follows: 1.5 μL of 10 × RT buffer, 1 μL of MultiScribe™ RT enzyme (50 U/μL), 0.19 μL of RNase inhibitor (20 u/μL), 0.15 μL of dNTP mix (100 mM total), 4.16 μL of nuclease-free water, and 3 μL of the specific primer. The primers used were: hsa-miR-33a-5p, 002135; hsa-miR-324-5p, 000539; hsa-miR-34a, 000426; hsa-miR-223, 002295; hsa-miR-132, 000457; hsa-miR-150, 000473; has-miR-328, 000572 (Applied Biosystem). The reverse PCR thermal profile was 16°C for 30 min, 42°C for 30 min, and 85°C for 5 min.
Quantitative PCR
Quantification of cellular miRNA levels was performed with quantitative PCR Brilliant II Probe Master Mix (Agilent Technologies, Santa Clara, CA) using specific primers and probes provided by TaqMan MicroRNA Assay Kit (Applied Biosystem). Briefly, 5 μL of cDNA was added to 12.5 μL of the master mix, 6.2 μL of nuclease-free water, and 1.3 μL of primer and probes. Real-time PCR was carried out using STRATAGENE Mx3000P (Agilent Technologies) thermocycler and analyzed with the included software. The cycling condition was as follows: 95°C for 10 min, 40 cycles of 15 s at 95°C, and 1 min at 60°C.
All data were presented as a relative quantification with efficiency correction based on the relative expression of target miRNAs versus the small-nucleolar RNU6B. The differential miRNA expression was calculated by the 2−ΔCt method and plotted as log 2−ΔCt.
Statistical analysis
Descriptive statistics were reported as count (percentage) for categorical variables and median with respective interquartile range (IQR) for continuous variables. Data were analyzed using Mann–Whitney U test or Kruskal–Wallis with post hoc pairwise Dunn's tests where appropriate. χ 2 test was used to analyze the differences in gender, coinfections, and ART distribution between groups. Two-tailed p values of <.05 were considered statistically significant. Pearson correlation analysis was performed to evaluate the correlation between miRNAs fold change of expression and demographics/clinical characteristics of the study population. Graphs and statistics were computed using GraphPad Prism 6 and STATA (StataCorp LLC, 4905 Lakeway Drive, College Station, 322 TX, USA), version 17.0.
Results
Description of study populations
A total of 90 PBMC samples were collected from individuals with HIV infection. As shown in Table 1, donors were similar in gender distribution and median age matched only between TND and LLV groups [TND, 55 years (45–58) vs. LLV, 57 years (53–60)], while the individuals belonging to the NAIVE group were younger with a median age of 44 (35–53) years. For TND and LLV groups, the median time from HIV-1 diagnosis was 17 (11–24) years, and the median duration of ART received before enrollment was 16 (10–22) years. The rate of HBV/HCV coinfection was similar among the three groups.
Demographics and Clinical Characteristics of Human Immunodeficiency Virus Type 1-Positive Individuals
Results were expressed as median (IQR) or a n (%).
Data were analyzed using the Mann–Whitney U test (+TND vs. LLV; ++TND vs. NAIVE; +++LLV vs. NAIVE) or Kruskal–Wallis where appropriate. χ 2 test was used to analyze the differences in gender, coinfections, and types of antiretroviral regimens between groups. p Value <.05 was considered statistically significant.
ART, antiretroviral therapy; IQR, interquartile range; LLV, low-level viremia; N/A, not available; NAIVE, treatment-naive; NNRTI, non-nucleoside reverse transcriptase inhibitor; NRTI, nucleoside reverse transcriptase inhibitor; PI, protease inhibitor; TND, target not detected.
Regarding ART, unequal distribution was observed among the HIV+ population 13 (47%) TND donors were treated with a combination of nucleoside reverse transcriptase inhibitors (NRTIs) and protease inhibitors (PIs), while 15 (54%) low-level viremia donors were taking NRTI in combination with non-nucleoside reverse transcriptase inhibitor (NNRTI).
The group of HD included 18 males and 10 females, with a median age of 52 years (IQR: 43–56).
HIV infection interferes with cell-associated miRNA expression
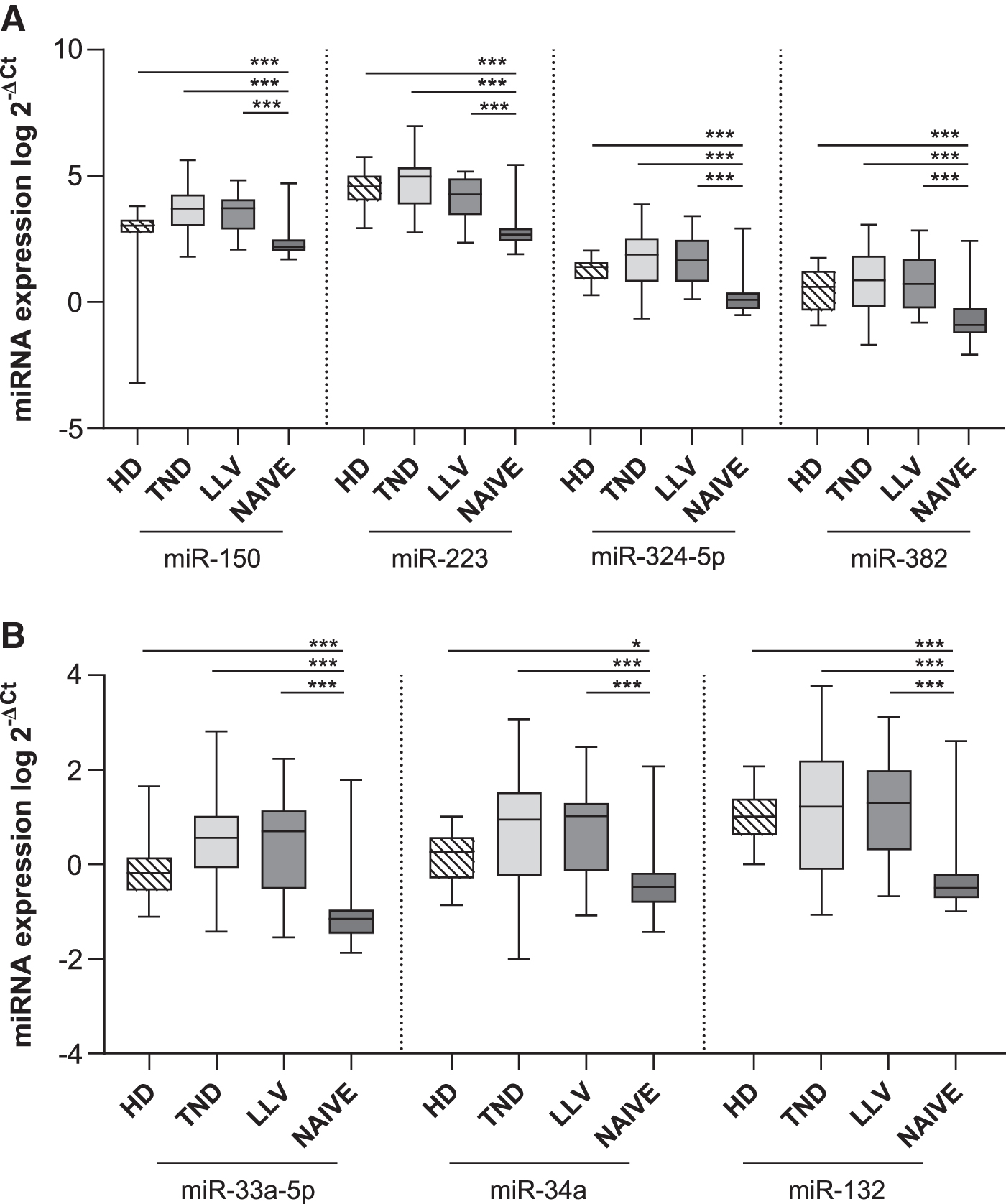
Preliminary analysis was conducted to assess the impact of HIV infection on miRNA expression profiles in PBMCs. Comparing the HIV population with HD group (Fig. 1A, B), only miR-223 expression levels were significantly (p = .005) downregulated in HIV+ population. When dividing HIV+ individuals into NAIVE and treated (TND+LLV) groups, different expression profiles were observed. Specifically, expression levels of the seven miRNAs analyzed were strongly downregulated in NAIVE group compared to TND+LLV and HD (p < .01 or p < .001 as noted in Fig. 1C, D). Besides, miR-150 (p = .025), miR-33a-5p (p = .03), and miR-34a (p = .035) levels resulted in upregulation in TND+LLV than HD (Fig. 1C, D).

To verify whether different viral suppression levels were associated with distinct miRNA profiles, ART-treated donors were stratified in TND and LLV and compared to the NAIVE and HD groups. The results confirmed the previously observed decrease in miRNA expression in NAIVE compared to HD, TND, and LLV (Fig. 2A, B).
NNRTI intake is associated with lower mir-33a-5p expression levels
Stratification of ART-treated donors based on the alternative intake of the main antiretroviral drug classes [NNRTI, PI, and integrase strand transfer inhibitors (INSTI)] revealed a downregulation of miR-33a-5p in people whose ART included NNRTI, reaching statistical significance compared to PI (Fig. 3; p = .032), but not INSTI (p = .249). The other miRNAs' expression levels did not significantly differ among these groups (data not shown).
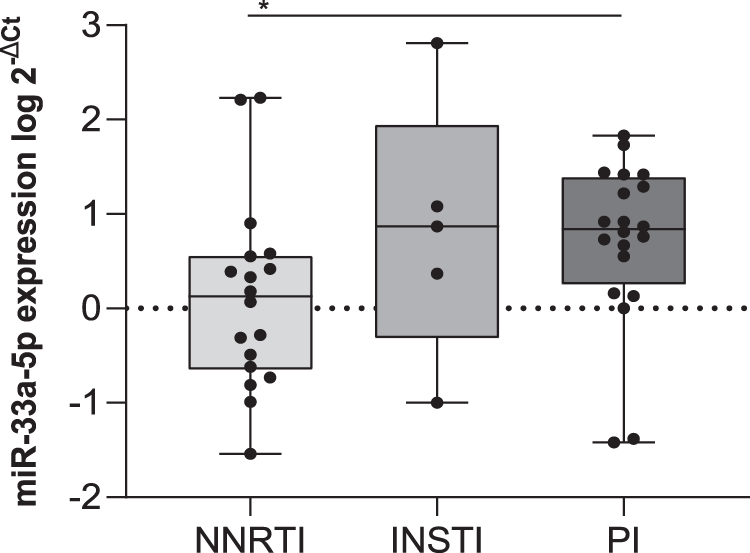
Comparison of miR-33a-5p expression levels among ART-treated donors taking NNRTI, INSTI, and PI. Expression differences between groups were assessed using a Kruskal–Wallis test followed by pairwise Dunn's test with Bonferroni correction for each pair of groups. Significance was deemed to be achieved at the p < .05 level. *p < .05. INSTI, integrase strand transfer inhibitors; NNRTI, non-nucleoside reverse transcriptase inhibitor; PI, protease inhibitor.
Levels of miR-33a and miR-150 correlate with CD4+ T cell counts and time from HIV-1 diagnosis, respectively, in individuals undergoing antiretroviral treatment
Considering each group independently (TND, LLV, and NAIVE), no correlation between miRNA expression levels and age, CD4+ T cell count, CD4+ nadir, years from HIV diagnosis, length of treatment, and viral load was observed (Table 2). However, in the treated population (TND+LLV) the expression fold change of miR-150 slightly correlates with time from HIV diagnosis (r = 0.33, p = .04; Fig. 4A). Conversely, expression levels of miR-33a-5p showed a trend negative correlation with nadir CD4+ count (r = −0.31, p = .05; Fig. 4B).
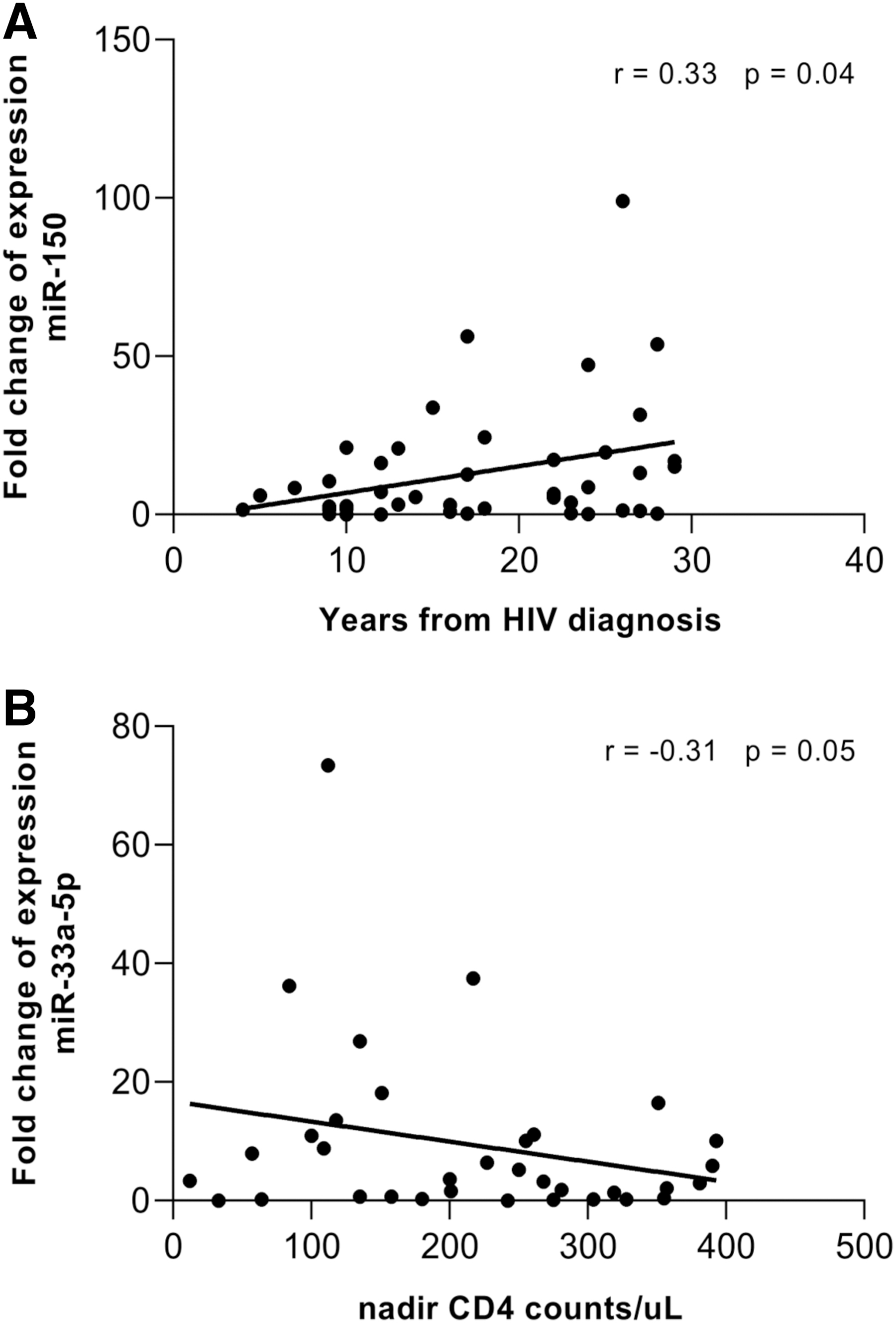
Correlation Between MicroRNA Expression Levels and Clinical Parameters
Correlation analyses were performed using Pearson correlation test, and p value <.05 was considered as significant.
Discussion
Herein, the expression levels of PBMC-associated miRNAs (miR-33a-5p, miR-324-5p, miR-34a, miR-223, miR-132, miR-150, and miR-328), previously reported to contribute to HIV-1 replication and maintenance of viral latency, were analyzed in unsorted PBMC samples from people living with HIV-1 showing different levels of viral suppression. The NAIVE group showed a high downregulation of all miRNAs analyzed compared to HD and ART-treated individuals. Despite the reduced expression of PBMC-associated miR-150 and miR-382 being consistent with literature data, 29 –32 the downregulation of miR-223 and the lack of correlation with viral load observed in our study disagree with Moghoofei et al. 32 findings indicating its overexpression in NAIVE donors and association with HIV RNA amount. In contrast, miR-150, miR-33a-5p, and miR-34a were found upregulated in ART-treated individuals, corroborating the wide dysregulation of miRNAs observed during HIV infection, and suggesting that it might be related to viral replication control and specifically for miR-150 related to the duration of viral infection.
Subsequently, to assess whether differential miRNAs expression was associated with different viral suppression levels, the HIV+ population was stratified into three groups (TND, LLV, and NAIVE) according to the HIV RNA values detected in the previous 3 years. This analysis confirmed the strong miRNA deregulation during uncontrolled viral replication.
As previously reported, miRNAs, including miR-150, miR-223, and miR-382, may contribute to the maintenance of viral latency in resting primary CD4+ T cells isolated from virologically suppressed individuals by targeting 3′ UTR of HIV-1 transcripts 22 ; their translation inhibition activity also seems to influence monocytes and macrophages susceptibility to HIV infection. 14,26 Cellular miRNAs can also affect HIV-1 replication by targeting host molecule expression exploited by the virus for its replication. For instance, miR-33a-5p has been shown to counteract HIV-1 replication by inhibiting the JNK1 expression, 20 a cellular kinase described to phosphorylate the viral integrase enzyme stabilizing its structure. 33
Therefore, the upregulation of mir-33a-5p and miR-150 observed in both TND and LLV than the NAIVE group support their role in dampening viral replication and the potential biomarker use of miR-150 to monitor disease progression and therapy response as previously proposed. 29 However, there are some differences between our findings and previous reports. In 2014, Reynoso et al. 20 reported downregulation of miR-33a-5p plasma levels in chronic progressors, defined by authors as individuals with HIV RNA <50 copies/mL. Herein, cell-associated miR-33a-5p level was evaluated, and its increased expression in ART-treated individuals might be due to a different miRNA distribution between intra- and extracellular compartments. Noteworthy, the lack of differences among TND and LLV groups indicates that the activity of these miRNAs is not sufficient to reach and maintain a complete virological suppression.
There is a growing body of evidence about the mutual interference between miRNAs and immune effector mechanisms, both innate and adaptive immunity, 34 which could influence the course of infectious diseases. Considering treated groups as a whole (TND+LLV), we found a trend toward an inverse correlation between miR-33a-5p expression level and nadir CD4+ T cell count. Hence, despite the pharmacological control of viral replication, miRNA expression could be related to immune system damage during the acute phase of the disease. In addition, T cell proliferation and immune activation status have been related to a differential miRNA expression between virologically suppressed and viremic HIV+ individuals. 31 Following this evidence, the persistent immune dysregulation commonly observed during treated HIV infection, 5,35 and linked to non-AIDS defining events, 36 may be the driving force behind differences in miRNA levels observed in our study.
As mentioned above, miR-223 can target 3′ UTR sequences of HIV RNA in primary resting CD4+ cells and may indirectly regulate cyclin T1 expression, thereby exerting an anti-HIV replication activity. 22,37 The lower expression observed in NAIVE individuals compared to TND and LLV groups indicates a progressive increase of miR-223 levels with the control of the viral replication. These findings support the hypothesis that miR-223 upregulation may result from a physiological response to viral clearance and/or indicate possible antiviral effects of the miRNA in people achieving sustained viral suppression as observed during HCV infection. 38
Finally, it is known that miRNAs contribute to interindividual variability in drug response, both in terms of toxicity and efficacy; thus, these molecules could be an ideal biomarker to monitor treatment efficacy. 39 Given that therapeutic regimens vary significantly in our cohort, HIV+ donors were grouped based on NNRTI, INSTI, and PI intake. Therefore we observed a significant decrease in miR-33a-5p levels in individuals undergoing NNRTI compared to subjects treated with PI, indicating that PI might be favored in hampering viral replication.
The present study has limitations that should be addressed. First, several samples were obtained from HBV/HCV coinfected individuals, and regarding the NAIVE group, data about other coinfections that could directly or indirectly influence the host miRNA profile were not available at the time of the present analysis. In addition, miRNAs expression levels were investigated in the whole PBMC population, preventing us from identifying the trend of their circulating counterparts and the individual cellular subset contribution. Furthermore, analyzing unsorted PBMCs, we could not distinguish the miRNA expression levels among HIV+ and HIV-negative cells.
In conclusion, PBMC-associated miR-33a-5p, miR-324-5p, miR-34a, miR-223, miR-132, miR-150, and miR-328 were found to be dysregulated during HIV infection, showing divergent profiles compared to healthy donors and among treated and NAIVE HIV+ individuals. The lack of correlation between miRNA levels and parameters such as CD4+ T cell count and viral load suggests the involvement of other processes that could be related to the inflammatory response, immune systems activation status, and to indirect effects of viral products that lead to modulation of miRNA expression not confined to a specific cell type. Therefore, further investigations are needed to provide new insight into the host processes required to achieve and maintain complete virological suppression and could enable us to identify new prognostic and treatment-response biomarkers useful to improve the management and the outcome of HIV-positive individuals.
Footnotes
Acknowledgments
The authors thank all the blood donors and health care workers who participated in the research.
Authors' Contributions
F.F., O.T., D.D.C. contributed to the conception and the design of the study. D.D.C., L.M., G.G., A.L., and M.S. contributed to data acquisition. D.D.C., F.F., O.T. performed interpretation of data. G.A., I.M., C.F., G.D.E., and G.I. revised and provided critical comments about the article. G.M. and D.D.C. performed statistical analysis. D.C.D. and T.O. contributed to the article writing and editing. All authors approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by funding from the Sapienza University of Rome (Progetto di Ricerca ATENEO 2019 n. 000307-19).