
Abstract
Human immunodeficiency virus (HIV) infection and the ensuing acquired immunodeficiency syndrome (AIDS) disproportionally affect young women, yet understanding of the factors promoting heterosexual transmission in the female genital tract is limited. Colonization with highly diverse, Lactobacillus-deficient communities (HDCs) increases a woman's risk of acquiring HIV-1 compared with colonization with Lactobacillus-dominated low diversity communities (LDCs). The polymicrobial nature of these communities has made it challenging to elucidate the microbial mechanisms responsible for modulating HIV susceptibility. Here, we analyzed conserved changes in small-molecule metabolites present in the cervicovaginal lavage fluid collected from women colonized with HDCs and LDCs with the goal of identifying possible chemicals influencing HIV infection. As in previous studies, we found that the catabolite of the branched-chain amino acid valine, 2-hydroxyisovalerate (2-HV), was a consistent component of dysbiotic HDC microbiota. Effects of 2-HV on HIV infection were assessed. In experimental infections with HIV, treatment with 2-HV increased infections of resting CD4+ T cells. To understand bacterial production of 2-HV in more detail, we cultured purified HDC and LDC bacteria and used mass spectrometry to identify two HDC bacteria that synthesize high levels of 2-HV. In contrast, protective vaginal Lactobacilli did not produce high levels of 2-HV. A genomic analysis of genes encoding 2-HV synthetic pathways showed a correlation between high-level production of 2-HV and pathways for synthesis of the immediate precursor 2-ketoisovalerate. Thus, 2-HV is a candidate mediator linking vaginal microbiome structure and heterosexual HIV transmission in women.
Introduction
Women and adolescent girls carry a heavy burden of human immunodeficiency virus (HIV) infection in sub-Saharan Africa: 25% percent of all new HIV infections occur in women 15–24 years old, yet they comprise only 10% of the total population. 1 To reduce the infectious burden in this population and others, it is critical to understand the factors influencing heterosexual HIV transmission.
The composition of the vaginal microbiome has been associated with HIV transmission risk. 2 –8 Vaginal microbial community composition can be grouped into five clusters called community state types (CSTs). 9 Four CSTs are dominated by a single species of Lactobacillus: CST I is dominated by L. crispatus, CST II by L. gasseri, CST III by L. iners, and CST V by L. jensenii. 9 CST IV is deficient in Lactobacillus and enriched in diverse anaerobic bacteria.
CST IV can be further subdivided into CST IV-A (modest proportion of Lactobacillus species and low proportions of anaerobic bacteria) and CST IV-B (higher proportions of anaerobic bacteria such as Atopobium, Prevotella, Sneathia, Gardnerella, Mobiluncus, Parvimonas, and others). 10
In a landmark study, Gosmann et al. demonstrated higher rates of HIV acquisition rates in women colonized with high diversity microbial communities (HDCs) that are deficient in Lactobacillus (reminiscent of CST IV-B). 2 Despite large differences in community composition within HDCs, women with HDC communities were associated with HIV acquisition rates four-fold higher than those colonized with low diversity Lactobacillus-dominant communities (LDCs) composed of primarily L. crispatus (CST I) or L. iners (CST III). 2
The HDCs exert numerous immunomodulatory effects on the female reproductive tract that have been proposed to increase HIV transmission risk. 11 –14 Vaginal epithelial barrier integrity, which plays a key role in limiting pathogenic infection, is impaired in experimental models by cervicovaginal fluid from women colonized by HDCs and allows for increased transmigration of cell-associated HIV. 13
In addition, HDCs promote pro-inflammatory cytokine expression and increase the number of infiltrating HIV target cells in the vaginal epithelium, 2 resulting in higher rates of local infection and increased probability of infected T cells entering circulation. 15 These changes have largely been attributed to the loss of protective lactic acid secreted by vaginal Lactobacilli, which suppresses HIV-associated pro-inflammatory cytokine expression in vaginal epithelial cells, 16 increases vaginal epithelial barrier integrity, 17 inactivates viral particles, 18 and kills HDC-associated Gram negative bacteria. 19
Although the loss of lactic acid by itself may contribute to enhanced HIV pathogenesis in HDCs, work from Pyles et al. has demonstrated that HDC colonization of vaginal epithelium co-cultures increases HIV burden relative to both LDC colonization and uncolonized controls, 20 suggesting that both the loss of Lactobacillus and the gain in HDC species may contribute to HIV transmission.
Reducing the HIV acquisition risk in women colonized by HDCs is a high priority. One strategy to reduce acquisition risk would be to transition women from an HDC to an LDC, but it is challenging to achieve long-term recolonization with Lactobacillus. Antibiotic treatments have high initial success rates at suppressing the growth of anaerobic bacteria, but most women suffer a recurrence in less than 3 months. 21,22
Relatively new strategies involve combining antibiotic treatments with experimental vaginal probiotics such as Lactin- V 23 or vaginal microbiome transplants. 24 Another strategy to ameliorate HIV acquisition risk would be to directly block viral replication. Although prophylactic antiretrovirals are available, their efficacies in some women are reduced due to poor adherence 25 –27 and/or HDC-mediated metabolism of the drug. 28,29 Thus, a deeper understanding of the mechanism contributing to enhanced HIV transmission is desirable to design improved interventions.
Hindering our understanding of the detailed mechanisms by which HDCs exert these immunomodulatory changes is the polymicrobial nature of these communities—many bacterial taxa having been associated with increased HIV susceptibility in different studies, complicating analysis. 2,30,31 Due to these difficulties, the mechanisms by which HDCs influence HIV replication and transmission have not been fully clarified.
Mass spectrometry analyses of vaginal fluid from women with dysbiotic vaginal microbiota, referred to as bacterial vaginosis (BV), revealed specific changes in the concentrations of organic acids present 32,33 that correlated with decreased vaginal epithelial barrier integrity, increased growth of pathogenic anaerobes, increased pro-inflammatory cytokine expression, and mucin breakdown. 32,33
Specifically, women colonized by high diversity communities had less lactic acid in vaginal discharge fluid and increased butyric acid, acetic acid, succinic acid, and propionic acid (reviewed in Aldunate et al. 19 ). The HDC-associated increases in these short chain fatty acids are thought to influence HIV pathogenesis by directly enhancing pro-inflammatory cytokine production from vaginal epithelial cells 34 and peripheral blood mononuclear cells, 35 but it has not been fully clarified whether these products can directly affect HIV replication in target cells.
Given the increased permeability of the vaginal epithelial barrier and that several bacterially derived metabolites are potent modulators of HIV target cell physiology, 36,37 it is possible that metabolites secreted by these communities are directly modulating vaginal intraepithelial T cell susceptibility to HIV-1 infection.
Here, we sought to identify molecules stimulating HIV infection produced by HDCs. We conducted a meta-analysis of published metabolomic studies comparing the cervicovaginal metabolome of women colonized by HDCs versus LDCs to reveal a subset of metabolites enriched in HDCs that change significantly and consistently.
We identify one metabolite, 2-hydroxyisovalerate (2-HV), that is a biomarker for HDC colonization; here, we report that 2-HV can stimulate HIV replication in primary resting CD4+ T cells. Using mass spectrometry, we demonstrate that two clinically important HDC members can produce and secrete 2-HV, but LDC-specific Lactobacillus species produce much lower levels.
Results
Metabolites enriched in HDC cervicovaginal lavage fluid
Previous studies of HDC-associated metabolites and HIV infection have focused on the production of short chain fatty acids such as butyric acid, propionic acid, succinic acid, and acetic acid, 34 examining how these may boost HIV replication, 38 –40 modulate immune responses, and influence barrier integrity of cervicovaginal epithelial cells. 17,34
To investigate other possible modulators of transmission, we compared cervicovaginal metabolomic data sets and quantified molecules showing enhanced production in HDCs. We analyzed four publicly available datasets with vaginal bacterial abundance and linked metabolomic data from women diagnosed with or without BV. 41 –44
There are many methods and techniques by which to discriminate HDC and LDC samples, 45 but we favored Nugent scoring because it is reproducible and sensitive. 46 Reported Nugent scores were used to bin vaginal samples into either HDCs (7–10) or LDCs (0–3). Samples with intermediate scores (4–6) were excluded for simplicity. Where possible, Nugent scores were related to other HDC and LDC discriminating characteristics, such as associated CST (Supplementary Fig. S1).
These studies recruited women of different ethnicities and geographic locations as summarized in Supplementary Table S1 and used a variety of different sequencing and mass spectrometry techniques to characterize the cervicovaginal microbiome and metabolome.
Multiple studies report different bacterial taxa that drive HDC and LDC cluster differentiation 9,10,41 –44 ; we investigated linked metabolomic analysis and were able to identify 20 metabolites that changed consistently and significantly in at least three out of the four studies (Fig. 1A). Of the twenty metabolites identified, only two were derived from metabolites other than amino acids: lactic acid and inosine.

Identification of metabolites consistently enriched in HDC colonized women. Data from four published studies were used to search for differences in the metabolome of cervicovaginal fluid from non- pregnant, STI (-) women colonized with HDCs or LDCs.
42
–45
Vaginal community types were divided into either LDC or HDC phenotype based on reported Nugent score (LDC = 7–10, HDC = 1–3).
Globally, multiple amino acids were depleted in HDC lavage fluid whereas metabolites derived from their metabolism were enriched. This could indicate that HDCs have a higher reliance on amino acid catabolism than their LDC counterparts, paralleling functional genomic analyses, suggesting that vaginal Lactobacillus have a relatively low representation of genes for amino acid transport and metabolism. 47
Three metabolites were consistently enriched in HDC lavage fluid from all four datasets: 2-HV, tyramine, and cadaverine. Work from others has shown that cadaverine and tyramine concentrations are both increased in HDCs 48 and in LDCs transitioning to HDCs. 49
2-HV is a catabolite of the branched-chain amino acid pathway produced during valine metabolism. It is typically found in human serum at low concentrations (<10 μM), but it can accumulate up to millimolar concentrations under disease conditions. 50 2-HV is a biomarker for microbiome-independent disorders such as maple syrup urine disease 51,52 and endometriosis, 53 and for microbiome-associated disorders such as neonatal septic shock 54 and insulin resistance. 55
In addition, 2-HV has been reported as an HDC biomarker in some studies of the vaginal microbiome. 41,56 Although 2-HV levels have previously been correlated with certain bacterial species, it remains unclear whether vaginal bacteria are capable of synthesizing and secreting this metabolite. An analysis of the bacteria associated with 2-HV in the metabolomic data showed that HDC strains were commonly positively associated, whereas Lactobacillus was negatively associated (Fig. 1B).
Effects of exogenous 2-HV on HIV replication in primary CD4+ T cells
We tested the effects of 2-HV, tyramine, and cadaverine on the frequency of HIV infection in HeLa-derived TZM-bl indicator cells and found no boosting of HIV infection with tyramine and cadaverine (data not shown). However, initial tests of 2-HV showed robust stimulation, so we investigated the effects of 2-HV further. Following initial surveys, we focused on the effects of 2-HV on HIV replication in primary CD4+ T cells.
CD4+ T cells were pre-treated with varying concentrations of 2-HV for 24–48 h and subsequently spinoculated with HIV BaL, an R5 trophic strain. As a positive control, aliquots of cells were also treated with 1 mM sodium butyrate, which has been reported previously to boost HIV replication. 40 Cells were infected by spinoculation and incubated for 8 days to allow for multiple viral replication cycles in the presence of 2-HV, which was maintained in the culture media.
At 8 days post-infection, the cells were analyzed for cell-free p24 and via flow cytometry to determine viability, intracellular p24 content, and T cell phenotype. Resting and activated T cells were compared. Results over multiple donors are summarized in Supplementary Table S2.
Boosting of cell-associated HIV infection was seen in studies of resting T cells in six out of seven donors (Fig. 2A). The seventh donor showed a similar upward trend in cell-associated p24, but this did not achieve statistical significance. Across all donors studied, Figure 2B shows that boosting in the presence of 16 mM 2-HV reached an average threefold increase in accumulation of cell-associated p24. Figure 2C shows that cell viability remained generally high after 2-HV treatment. For measures of extracellular p24, resting T cells from three donors were tested and two showed an increase in p24 associated with 2-HV treatment (Supplementary Fig. S2A and Supplementary Table S2).
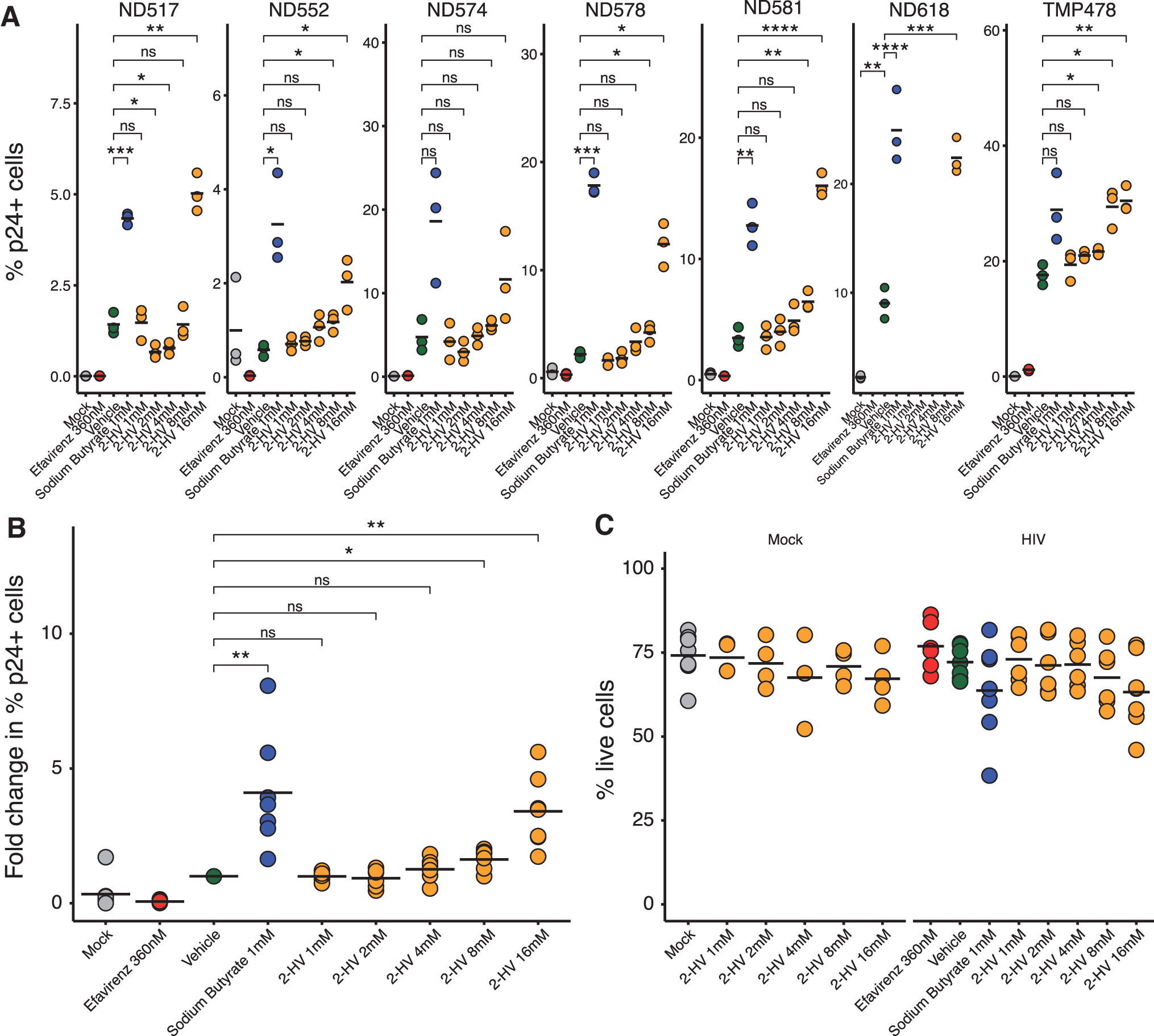
Exogenous 2-HV addition boosts HIV infection in resting primary CD4+ T cells. Effects of exogenous 2-HV addition were tested in resting primary CD4+ T cells infected with 10 ng of BaL p24/1 × 106 cells for effects on HIV infection and cell viability. Cells were pretreated with 2-HV or mock treated for 24–48 h, spinoculated with HIV BaL, and infected for 8 days. Eight days post-infection, cells were fixed and stained for flow cytometry. Efavirenz and sodium butyrate were used for negative and positive controls, respectively.
Increases in the presence of 2-HV were inconsistent for samples from activated T cells. T cells were activated by treatment with antiCD3 antibodies before HIV infection. In the presence of 2-HV, samples from two out of three donors did not show an increase in intracellular p24 (Supplementary Fig. S2B and Supplementary Table S2). Parallel assays of cell-free p24 after infection of activated cells showed a significant increase in only one out of four donors tested (Supplementary Fig. S2C). Thus, boosting of p24 production was inconsistent in T cells activated by exposure to anti-CD3 antibodies.
A possible mechanism for increased p24 production after 2-HV treatment is that 2-HV may be augmenting T cell activation, and prior activation may thus reduce the magnitude of stimulation of 2-HV addition. We investigated the possible effects of 2-HV on T cell activation by measuring the expression of two immune activation markers using flow cytometry: CD38 and HLA-DR. 57,58
We observed increased surface activation markers in cells exposed to 2-HV in two out of four donors tested (Supplementary Fig. S3A). We next investigated the relationship between boosting by 2-HV and the starting level of p24, reasoning that cells already activated may produce high levels of p24 and so be less susceptible to boosting (Supplementary Fig. S3B, C).
In the vehicle control, cells with higher starting levels of extracellular p24 were significantly less likely to show boosting (Supplementary Fig. S3B). For comparisons based on intracellular levels of p24, there was a trend toward greater boosting by 2-HV associated with lower unstimulated p24 values. Thus, these data on activation and 2-HV stimulation are equivocal, but they suggest that T cell activation by 2-HV may be one contributor to increased HIV p24 production.
Production of 2-HV from vaginal bacteria
We next assessed in more detail which vaginal bacterial species can produce 2-HV using metabolomic methods to quantify secretion of 2-HV into culture supernatants for several species (Supplementary Table S3). Culture supernatants were first analyzed using untargeted gas chromatography-time of flight (GC-TOF) mass spectrometry. Many metabolites changed in concentrations relative to the media controls, and some differed between HDC and LDC strains (Fig. 3A and Supplementary Table S4).

Production of 2-HV and other metabolites by HDC bacteria. Metabolomic analysis was conducted on bacterial cell culture supernatant from HDC and LDC community members. Bacteria were grown in liquid culture for 2–3 days in sBHI media treated without Oxyrase (Media 1) and with Oxyrase (Media 2) to enhance growth of strict anaerobes.
Many paralleled the amino acids and metabolites in Figure 1A. Most notably, we found that three species from HDCs produced higher levels of secreted 2-HV: Gardnerella vaginalis, Atopobium parvulum, and Veillonella montpellierensis (Fig. 3A). No LDC strains were similarly high producers.
To confirm the production of 2-HV, we carried out targeted quantification of 2-HV in media from bacterial cell cultures. We found that G. vaginalis and A. parvulum secrete significantly more 2-HV into supernatant than did other strains, paralleling data from the untargeted metabolomic analysis (Fig. 3B), producing between ∼0.1 mM and 0.3 mM of 2-HV (Supplementary Table S5).
V. montpellierensis showed a trend toward higher production but did not reach statistical significance. None of the Lactobacillus strains produced high levels of 2-HV. Thus, several HDC members but not LDC members are robust producers of 2-HV.
Genomic analysis of bacterial enzymes potentially mediating 2-HV production
Our pilot metabolomics experiments suggested that certain vaginal bacteria may be able to produce and secrete 2-HV. However, we were only able to test a subset of bacteria from HDCs. It would be useful to be able to predict potential high-level secretors of 2-HV in heterogeneous HDCs simply by analyzing vaginal bacterial genomes.
Therefore, we carried out a genomic investigation of possible enzymes contributing to high-level synthesis. Previous work has suggested that 2-HV in humans can be synthesized by transforming valine into 2-ketoisovalerate (2-KIV), then further transforming 2-KIV into 2-HV (Fig. 4A, bottom). 59 In another pathway, pyruvate can be transformed via enzymes of the branched-chain amino acid synthetic pathway, leading after several steps to production of 2-KIV, 60 –63 which can then be transformed to 2-HV (Fig. 4A, top). 63
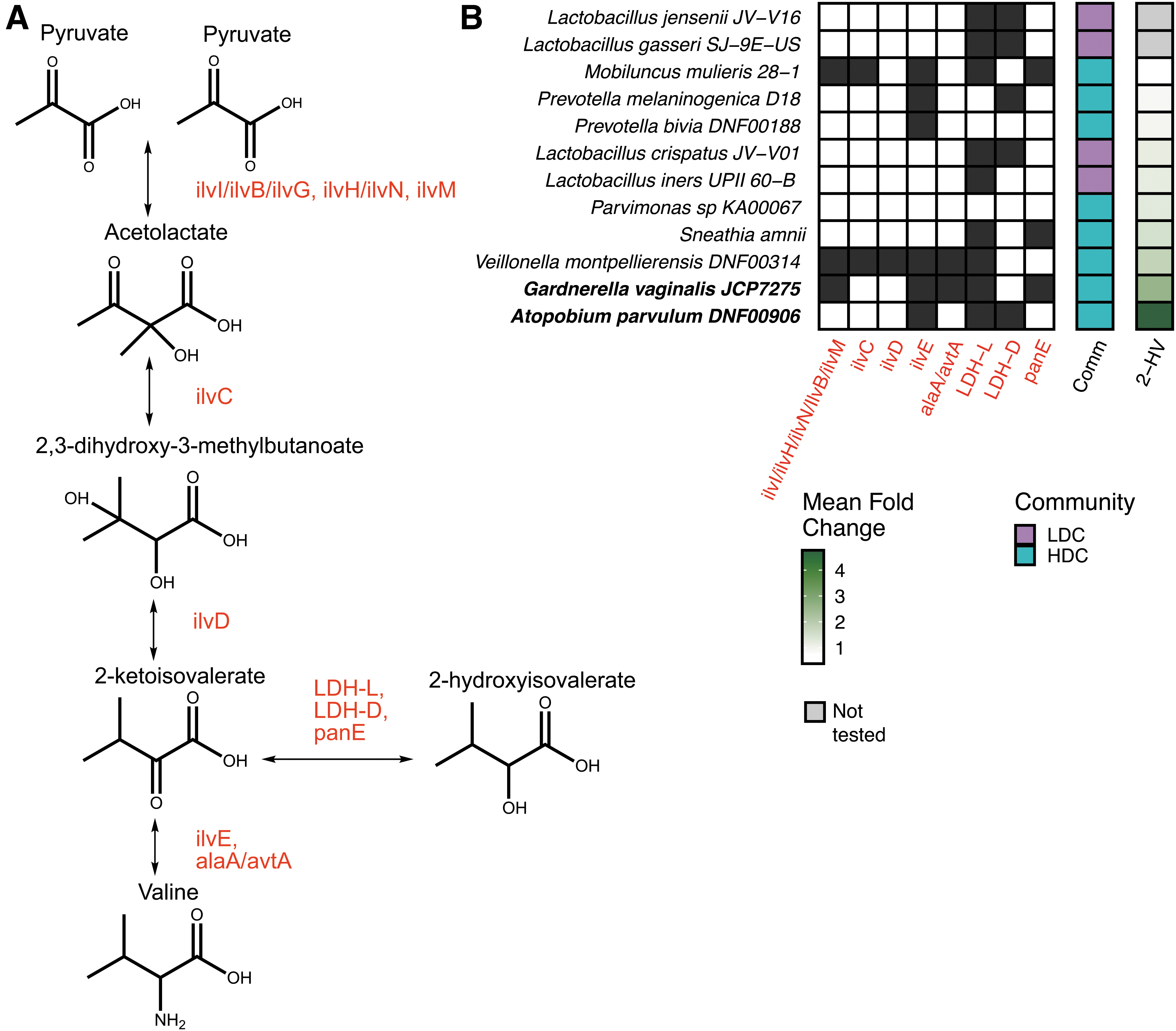
Predicted 2-HV synthesis pathway and genomic analysis of 2-HV synthetic potential.
We thus investigated the representation of enzymes mediating these reactions in the genomes of the HDC and LDC bacteria tested in Figure 3. Of course, the presence or absence of genes for the needed enzymes is only one factor determining whether 2-HV is produced. Efficient production requires presence of the needed precursors for each pathway and sufficient expression of each gene to allow efficient pathway function, both of which were not measured here. In addition, though we used validated conditions for annotating the presence of the encoded enzymes, it is of course possible that enzymes may have diverged sufficiently in sequence in some strains to be misidentified in our analysis.
Nevertheless, some generalizations can be suggested from the data (Fig. 4B). The three HDC strains producing high levels of 2-HV (G. vaginalis, A. parvulum, and V. montpellierensis) had in common genes mediating the production of 2-KIV from valine (ilvE and/or avtA) and production of 2-HV from 2-KIV (LDH-L and/or panE) (Supplementary Table S6).
For this analysis, we accepted lactate dehydrogenase (LDH) as an enzyme capable of transforming 2-KIV into 2-HV because this enzyme in humans has been implicated in carrying out this reaction. 59 In contrast, only one of the seven strains producing low levels of 2-HV, the HDC strain Mobiluncus mulieris, had genes for both of the steps for transforming valine to 2-HV.
One moderately high-level producer of 2-HV, V. montpellierensis, had the full branched-chain amino-acid synthetic pathway detectably intact. Thus, only Veillonella seems likely to contribute to 2-HV production via the branched-chain amino acid synthesis pathway. None of the Lactobacillus strains studied detectably encoded enzymes for pathways leading to the production of 2-KIV.
Thus, we conclude that (1) machinery for high-level production of 2-HV is commonly found in HDC but not LDC strains; (2) high-level production of 2-HV is likely commonly via catabolism of valine; (3) most HDC and LDC strains do not have intact pathways for branched-chain amino acid synthesis and may need to scavenge these amino acids from the environment; and (4) Veillonella may synthesize 2-KIV and valine from pyruvate.
2-HV treatment decreases production of several pro-inflammatory cytokines
Another possible role for 2-HV could be via modulating cytokine production from cells of the vaginal epithelium. We thus queried how 2-HV treatment affects pro-inflammatory cytokine expression in vaginal epithelial VK2 cells using a Luminex assay (Supplementary Fig. S4).
Lactic acid treatment exerts anti-inflammatory effects on vaginal epithelial cells by increasing the production of the anti-inflammatory cytokine IL-1RA and dampening the production of pro-inflammatory cytokines TNF-a, IL-6, IL-8, RANTES, and MIP-3alpha. 34 We thus assessed the effects of 2-HV on these molecules and the interferon-inducible chemokine IP-10. 64
We found that high concentrations of 2-HV decreased the expression of many pro-inflammatory cytokines, including IL-6, IL-8, MIP-3a, RANTES, and IP-10, and increased the expression of the anti-inflammatory IL-1RA cytokine. We did not detect significant TNF-a secretion in any samples. Thus, we conclude that 2-HV does not stimulate pro-inflammatory cytokine expression under these conditions using this model of the vaginal epithelium.
Discussion
Here, we describe metabolites that are consistently enriched in vaginal fluid from HDCs across multiple studies and characterize the effect of one of these, 2-HV, on HIV infection. Previous work has shown that Lactobacillus-deficient, highly diverse vaginal microbiomes are associated with higher rates of HIV transmission, 2 prompting the investigation of possible mechanisms.
We report here that the addition of 2-HV enhanced HIV infection in multicycle infections in resting primary T cells. We also assessed the bacterial production of 2-HV and identified several clinically relevant HDC bacteria that synthesize and secrete 2-HV, likely contributing to the enrichment of 2-HV observed in cervicovaginal fluid in the presence of HDC communities.
In contrast, Lactobacillus species, characteristic of LDCs that are protective against HIV, did not produce abundant 2-HV. Our data suggest that differences in the branched-chain amino acid synthesis pathway underlie the ability of vaginal bacteria to secrete 2-HV. Further work on this biosynthetic pathway is warranted to assess whether modulation could reduce HIV replication.
Tyramine and cadaverine were also found to be enriched in HDCs and recently reported to be potential biomarkers. 48,49 We tested the effects of tyramine and cadaverine alongside 2-HV in preliminary experiments, but we did not see any effects on HIV replication. It would be useful to investigate the possible effects of tyramine and cadaverine over a wider range of cell types, concentrations, and conditions to assess their possible effects on HIV infection more fully.
This work adds to the literature on how metabolites produced by the microbiota can affect HIV replication and transmission. One thoroughly characterized metabolite is lactic acid, a protective metabolite produced from vaginal Lactobacillus species, 17 –19,34,65 that inhibits the growth of pathogenic anaerobes such as G. vaginalis, 66 reduces pro-inflammatory cytokine production, 16,34 inactivates viral particles, 18 and enhances epithelial barrier integrity. 17
Less progress has been made in identifying metabolites from HDCs that may enhance HIV acquisition and replication. Much of the work has been focused on increases in short chain fatty acids such as butyrate, acetate, and succinate, which are observed when lactic acid production is low. 19,32,33 These metabolites can enhance pro-inflammatory cytokine production 34,35 and directly boost HIV infection. 38,40 Butyrate was used a positive control here.
These metabolites have been detected in cervicovaginal fluid, 44 but we have not identified them as being consistently higher in cervicovaginal fluid from HDCs, so they do not seem to be responsible for the enhancement of infection by HDCs. In contrast, 2-HV boosts infection and is made selectively by HDC members.
The vaginal epithelial barrier is known to be compromised in the presence of HDC communities, providing an opportunity for direct contact between 2-HV and T cells and enhancement HIV pathogenesis. 67 –70 In support of this, high concentrations of 2-HV and other catabolites of the branched-chain amino acid pathway are associated with inflammatory disorders. 52 –54,71
Despite this association, we did not obtain clear evidence for enhanced pro-inflammatory cytokine expression by vaginal epithelial cells. However, our study was restricted to studying vaginal VK2 cell monolayers that do not reflect the larger tissue organization of the female reproductive tract, nor did we test this in other cell types that may contribute to pro-inflammatory cytokine production in the vaginal tract.
The pathways yielding 2-HV have only recently been fully clarified. Cheong et al. used synthetic biology to establish a de novo pathway for synthesis of 2-HV in bacterial cells, which showed that it can be made from pyruvate via the branched-chain amino acid synthetic pathway. 63 2-HV can also be produced by the catabolism of valine.
In the last step of both pathways, 2-HV is produced by the reduction of its precursor, 2-KIV. This reaction can be carried out by bacterial panE, and in humans is also known to be carried out by LDH. In humans, LDH was found to promiscuously catalyze this reaction under pathological conditions in the presence of large amounts of the 2-KIV precursor. 59 The LDH enzyme belongs to a large family of 2-hydroxyacid-dehydrogenase enzymes in bacteria.
This enzyme family appears unusual in their substrate promiscuity, with multiple enzymes capable of catalyzing related reactions with different substrates, complicating definitive analysis. 72 We thus included LDH among the enzymes in bacteria capable of catalyzing this reaction. All three of the strains producing high levels of 2-HV encode ilvE and LDH-L and/or panE.
For the two bacteria that produced 2-HV at high levels (G. vaginalis and A. parvulum), the valine catabolism pathway was the only identifiable route to 2-HV. For the third lower-level producer (V. montpellierensis), both the branched-chain amino acid synthetic pathway and the valine catabolism pathway were candidates for the production of 2-HV.
The mechanism by which 2-HV boosts HIV infection is not clarified by our work. Our preliminary data suggest that 2-HV does not affect viral replication solely by augmenting T cell activation, though this may be a contributor. Observing a quantitatively substantial effect of 2-HV addition required multicycle analysis of the viral replication cycle over 8 days, complicating analysis of the step affected in viral replication. Future studies will be needed to clarify the mechanism.
One possible utility of our studies is in the generation of possible small-molecule enhancers of transduction for use with lentiviral vectors. Previously, sodium butyrate has been used to boost transduction. 73 –75 It will be useful to test whether 2-HV is able to boost transduction in cell types relevant in lentiviral vector-based gene therapy. If so, 2-HV may be useful as a simple additive for increasing the efficiency of gene transfer.
This study has several limitations. It is unknown how high the concentrations of 2-HV are that can be reached under physiological conditions in vaginal fluid, so it is not fully clear that tests of HIV infection in our model involve meaningful concentrations. We did not show the effects of 2-HV in an animal model of transmission.
We did not work out the step of the viral replication cycle affected by 2-HV. We did not investigate other compounds produced by HDC communities for their effects on HIV infection or for possible synergies with 2-HV. Our analysis of 2-HV's effect on human cell types was limited to studying pro-inflammatory cytokines in an immortalized vaginal epithelial cell line, which may not adequately model responses in the full tissue.
We also did not assay other factors than may contribute to HIV transmission such as epithelial barrier integrity. It will be useful to investigate these issues in future experiments.
In summary, this study highlights the potential importance of 2-HV in HIV transmission and provides a platform by which to test other HDC metabolites for their effects on HIV replication and transmission. Long term, a deep understanding of the effector mechanisms by which HDCs promote HIV infection may allow modulation of the environment of the female genital tract to interfere with transmission.
Methods
Meta-analysis of published metabolomics data
A literature search was conducted for open access data containing mass spectrometry analysis of cervicovaginal fluid from women colonized by either Lactobacillus dominant or high-diversity communities, yielding four studies with matched metagenomics and metabolomics. 41 –44 Lactobacillus-deficient microbial communities were characterized by reported Nugent score.
Samples with Nugent scores of 7–10 were defined as HDCs, whereas Nugent scores of 1–3 were defined as LDC. Samples with intermediate Nugent scores of 4–6 were excluded from analysis for simplicity (Supplementary Table S1). All subsets analyzed from these cohorts were negative for sexually transmitted infections and were from non-pregnant women.
To facilitate comparisons between different metabolomics studies, we developed a script to search for metabolite synonyms using the HMDB database (
Preparation of 2-HV and control treatments
Two-hydroxyisovalerate (Sigma 219835) and sodium butyrate (Sigma 303410) stocks were prepared fresh before experiments from powder stocks. Efavirenz was stored at −80°C. All metabolites were diluted in water at 10 × final concentration and filtered with 0.25 μm filters, then diluted into cell cultures.
T cell media
Primary T cells were maintained in CTS Optimizer media (ThermoFisher A1048501) supplemented with 10 ng/mL of recombinant IL-2 (Stemcell Technology 78036).
T cell activation
For T cell activation experiments, primary donor CD4+ T cells were obtained from the University of Pennsylvania Human Immunology Core and resuspended at a density of 1 × 106 cells per mL. Cells were activated with 0.5 μg/mL of soluble antiCD3 clone OKT3 monoclonal antibody (ThermoFisher 16-0037-81) for 3 days at 37°C.
Metabolite treatment of primary CD4+ T cells
Primary donor CD4+ T cells were obtained from the University of Pennsylvania Human Immunology Core. For infections, 1 × 106 T cells per replicate were seeded in 96-well V-bottom plates and resuspended in CTS Optimizer media with IL-2 with metabolite or control treatments for 24–48 h in a 37°C incubator.
After pre-treatment, cells were spinoculated by resuspending in media with 10 ng of R5 trophic BaL HIV and centrifuging for 2 h at 1,200 g at 37°C. Following spinoculation, cells were resuspended in CTS Optimizer media with IL-2 with metabolite and control treatments and incubated for a total of 8 days. Media and metabolites were replaced every 3–4 days until staining for flow cytometry.
Flow cytometry staining
Media was aspirated from plates; cells were washed once with PBS, resuspended in 50 μL of 1:60 Aqua LIVE/DEAD stain, and incubated in the dark for 10 min. T cell surface stain was prepared with 0.5 μL of CCR5-APC (Biolegend 359122), 1 μL of CD38-PE (BD 555460), and 2 μL of HLA-DR-BV605 (BD 562845) in 50 μL of PBS. The cocktail was added to cells and incubated in the dark for 20 min at room temperature.
After incubation, cells were washed once with 200 μL of staining buffer by centrifuging at 1,500 rpm for 5 min. Cells were fixed in 200 μL of Cytofix/Cytoperm and incubated for 30 min at room temperature in the dark. Cells were washed twice with 200 μL of Perm/Wash buffer and centrifuged at 1,500 rpm for 5 min each. Intracellular p24 staining was conducted with 0.1 μL of HIV-FITC (Beckman Coulter 6604665) in 50 μL final volume of Perm/Wash buffer. The stain was incubated for 1 h at room temperature in the dark.
After intracellular staining, cells were washed once with Perm/wash buffer at 1,800 rpm for 5 min. Cells were fixed with 250 μL of 1% paraformaldehyde and transferred to FACS tubes before running on a BD® LSR II Flow Cytometer. All samples were analyzed within 4 days of fixing. One batch of patient cells was excluded for failing to show boosting with sodium butyrate.
Extracellular p24 measurement
One hundred fifty microliters of primary T cell supernatants were collected from samples before flow cytometry staining and stored at −80°C until analysis. Cell-free p24 was measured via an enzyme-linked immunosorbent assay kit (R&D Systems HIV-1 Gag p24 DuoSet ELISA kit, catalog number DY7360-05). Assays were conducted according to the manufacturer's instructions.
Anaerobic cultures of vaginal bacteria
Bacteria were streaked from glycerol stocks and grown for 24–48 h on Brucella blood agar plates (Anaerobe Systems) in airtight Snapware containers with anaerobic gas-generating sachets. Five milliliters of starter cultures were prepared by inoculating supplemented BHI media (sBHI; BHI powder with 1% yeast extract, 0.1% glucose, 0.1% sucrose, and 5% human serum) treated with Oxyrase for Broth (Bioworld 30620059) to maintain anerobic conditions.
Starter cultures were grown for 24–48 h before diluting 1:10 in fresh sBHI media treated with Oxyrase. Cultures were incubated anaerobically for 24–72 h. Culture density was obtained by measuring optical density and CFU dilution plating on Brucella blood agar plates or sBHI + agar plates. Samples were centrifuged at 4,000 rpm for 10 min at 4°C. Culture supernatant was extracted and aliquoted in 1 mL volumes for analysis via mass spectrometry. All samples were snap-frozen in liquid nitrogen and stored at −80°C.
Mass spectrometry
Mass spectrometry analysis was performed on culture supernatants by the University of California – Davis's West Coast Metabolomics Center using a combination of both untargeted GC-TOF on a panel of metabolites involved in primary metabolism and targeted absolute quantification of 2-HV. Samples were extracted using 1 mL of 3:3:2 acetonitrile:isopropyl alcohol:water (v/v/v).
Half of the sample was dried to completeness and then derivatized using 10 μL of 40 mg/mL of methoxyamine in pyridine. Samples were shaken at 30°C for 1.5 h. Ninety-one microliters of N-methyl-N-(trimethylsilyl)trifluoroacetamide and fatty acid methyl ester were added to each sample and shaken at 37°C for 30 min to finish derivatization.
Samples were then vialed, capped, and injected into a 7890A GC coupled with a LECO TOF. Data were acquired using the following chromatographic parameters, with more details to be found in Fiehn et al. 76 0.5 μL of derivatized sample was injected using a splitless method onto a RESTEK RTX-5SIL MS column with an Intergra-Guard at 275°C with a helium flow of 1 mL/min.
The GC oven was set to hold at 50°C for 1 min, then ramped to 20°C/min to 330°C, and held for 5 min. The transferline was set to 280°C while the electron ionization source was set to 250°C. Data were collected from 85 m/z to 500 m/z at an acquisition rate of 17 spectra/s.
Mass spectrometry data analysis
The data were preprocessed using ChromaTOF versus 2.32. No smoothing was applied. The peak width was 3 s. The baseline value was subtracted at the noise level threshold. Throughout the chromatogram, a signal-to-noise level of 5:1 was applied along with automatic mass spectral deconvolution. BinBase (available from
The BinBase algorithm was used to report apex masses. Files describing results were exported as .txt files and included absolute spectra intensities. The data were further processed using a filtering algorithm from the BinBase database (rtx5). Within BinBase, we used the following settings for unbiased metabolomics: chromatogram validity (<10 peaks with intensity >107 counts s−1), retention index marker detection (MS similarity >800, validity of intensity range for high m/z marker ions), and a 5th order polynomial regression were used to calculate the retention index.
In the analysis, spectra were truncated to 5% base peak abundance and associated with database entries, starting with the most abundant spectra and progressing to least abundant, according to these matching filters. (1) The retention index window was ±2,000 U (equivalent to about 2 s retention time). (2) The unique ions and apex masses were validated as follows: unique ions need to be included in apex masses and present at >3% of base peak abundance. (3) The mass spectrum must fit criteria based on peak purity, signal-to-noise ratios, and a final isomer filter.
We constructed a new database for failed spectra. These were entered if s/n >25, purity <1.0 and the presence in the study design class was >80%. All quantification was reported as the peak height with the unique ion used as the default; the exception was if a different ion was set in BinView.
For unidentified metabolites, entries were reported that were detected in more than 10% of the samples for each design class from the miniX database. Following the main analysis, a post-processing module was employed to replace any missing values from the .cdf files. These replaced values were labeled as “low confidence.” For high confidence metabolites, the number of detections was recorded along with the ratio of the average height of replaced values to high-confidence peaks. Data were subsequently subjected to manual curation if needed before release.
As controls for the metabolomic analysis, we included Lactobacillus species, and tested them in both Oxyrase negative and positive culture conditions. An important positive control is production of lactic acid from Lactobacillus, and this was seen in samples tested in media lacking Oxyrase. However, Oxyrase is stored in a buffer containing a high concentration of lactic acid, and thus secretion of lactic acid by Lactobacillus species was masked in samples analyzed in Oxyrase media associated with the high starting levels (Supplementary Fig. S5).
Genome analysis
Protein sequences from bacterial genera of interest were downloaded from the KEGG database. 60 –62 Genomes used are in Supplementary Table S3. NCBI protein BLAST was conducted against the reference protein sequences, with positive hits comprising greater than or equal to 80% query coverage and 50% sequence identity. As a positive control, the highly conserved ribosomal subunit protein rsmG was queried in each genome using the same search criteria and found in all genomes tested. Matched enzyme hits are reported in Supplementary Table S6.
Vaginal epithelial cell culture
VK2 cells were cultured in keratinocyte serum-free medium (ThermoFisher; catalog number 17005042) supplemented with 0.1 ng/mL of human recombinant epidermal growth factor, 0.05 mg/mL of bovine pituitary extract, and 44.1 mg/L of calcium chloride (final concentration of 0.4 mM). VK2 cell monolayers were cultured in six-well plates and treated with 2-HV, poly(I:C) (R&D Systems; catalog number 4287), or lipopolysaccharides (Cell Signaling Technology; catalog number 14011) for 24 h. Cell supernatants were collected, clarified by centrifugation, flash frozen in liquid nitrogen, and stored at −80°C until analysis.
Luminex cytokine assays
Luminex assays were carried out by the Immunology Core of the Department of Pathology and Laboratory Medicine, Perelman School of Medicine, University of Pennsylvania, using the Millipore Sigma Kit (catalog number HCYTA-60K-09) according to the manufacturer's instructions. Results were analyzed using Luminex® xPONENT® 4.2; Bio-Plex Manager™ Software 6.1.
Footnotes
Acknowledgments
The authors are grateful to members of the Bushman and Collman laboratories for help and suggestions. They would like to thank the University of Pennsylvania human immunology core for providing CD4 T cells and running all Luminex assays as well as the flow cytometry core for access to equipment and training. They would also like to thank the service core at the University of California Davis West Coast Metabolomics Center, especially Kelly Paglia and the Oliver Fiehn laboratory, for conducting mass spectrometry analysis of their bacterial supernatants and supplying text on methods.
Author Disclosure Statement
The authors have no competing interests as defined by BMC, or other interests that might be perceived to influence the results and/or discussion reported in this paper.
Authors' Contribution
K.M. conceived the study; K.M., C.M., and F.D.B. analyzed the data; and K.M. and F.D.B. wrote the paper. All authors read and approved the final manuscript.
Funding Information
This work was supported in part by NIH grants R01AI129661, T32HL007775, U19AI117950, U19AI149680, UM1AI164570, R01CA241762, The PennCHOP Microbiome Program, The Center for AIDs Research at the University of Pennsylvania (P30-AI045008), and a grant from ViiV Healthcare.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.