
Abstract
Human immunodeficiency virus (HIV) infection is still an important public health problem, which justifies the research of new therapies to combat it. Recent studies show that Extracellular Traps (ETs) are cellular mechanisms useful in the capture and destruction of some viruses, such as the HIV. Here, we show that neutrophils from peripheral blood, genital tissues, and placenta are activated when exposed to human immunodeficiency virus type 1 (HIV-1) and release Neutrophil Extracellular Traps (NETs). The NETs can capture, neutralize, and inactivate the virus and, also, protect other target cells from HIV infection, as long as the DNA and other constituents of the NETs remain intact. Further, the review indicates that the immunoprotective role of NETs in the context of HIV-1 infection is a promising finding for the development of new antiviral therapies. It is necessary, however, the development of studies that evaluate the tissue injury that NETs can cause and the biological relationships with other cells to improve them as therapeutic targets.
Introduction
In the world, about 38.4 million people are living with the Human Immunodeficiency Virus (PLHIV), a pandemic marked by an annual incidence of 1.87 million cases. 1 In Brazil, since 2019, 966,058 new cases were reported, 2 a scenario that can be explained by weaknesses in the process of screening, diagnosis, and/or treatment of the infection, 3 favoring the continuity of the HIV transmission chain. 4
One of the strategies to combat HIV infection is the development of new immunotherapies. However, there are still gaps in knowledge about the immune response to HIV. Although most studies have been developed in the context of the adaptive immune response, evidence shows that the innate immune system provides important defense mechanisms to the host, including the detection of pathogen-associated molecular patterns (PAMPs). 4 Delays in effective immunotherapies development against HIV are a major public health problem.
Polymorphonuclear cells are innate immune cells specialized in developing strategies to combat various infectious agents. 5 –7 One of these combat mechanisms is the release of Extracellular Traps (ETs), by neutrophils, 8 eosinophils, 9 basophils, 10 and macrophages. 11 ETs are released by neutrophils when they recognize HIV. 12 These cells have an arsenal of effector functions that can destroy HIV, constituting themselves as soldiers on the front line in acute infection. 13 –15 However, it is known that HIV virions use several immune escape mechanisms. 12
We reviewed the role of ETs in the immunopathogenesis of HIV infection as a well-orchestrated innate immune response based on (1) analysis of ET-producing cellular profiles; (2) acting on different body sites; (3) effector response in the acute and chronic phases of infection; (4) regulatory mechanisms of the cellular response; and (5) the role of extracellular DNA networks in the immunoprotection of target cells susceptible to infection.
However, it is worth emphasizing that the findings on the role of immune cells in extracellular DNA network release are recent, and there is still a lack of consensus in the empirical literature on ETs and HIV.
In accordance with our guiding question “Do Extracellular Traps have immunoprotective properties against HIV infection in humans?,” we aimed to gather the most recent evidence on the possible immunoprotective property of ETs in HIV infection in humans. Therefore, here we reviewed recent scientific articles focusing on different characteristics shown in Table 1, and they are presented in a detailed discussion next.
Characterization of the Studies: Objectives, Study Design, Number of Participants, and Body Site Analyzed
The experiments were performed with a cell pool of healthy individuals (number not informed).
HIV-1, human immunodeficiency virus type 1; NA, not applicable; NET, neutrophil extracellular trap.
Release of Neutrophil Extracellular Traps in Chronic HIV Transmission and Infection
The entry points for HIV in the human body are diverse, with genital tissues as the main route of infection. 16 Upon entering this medium, the virus comes into contact with resident leukocytes, including genital neutrophils, which are phenotypically different from the neutrophils found in peripheral blood. 17 In this context, neutrophils in the female genital tract respond to human immunodeficiency virus type 1 (HIV-1) Virus-like Particles (VLPs) by stimuli from Toll-Like Receptors 7 and 8 (TLR7/8) and consequently trigger a cascade of intracellular processes that culminate in the formation and release of neutrophil extracellular traps (NETs) 13 (Fig. 1).
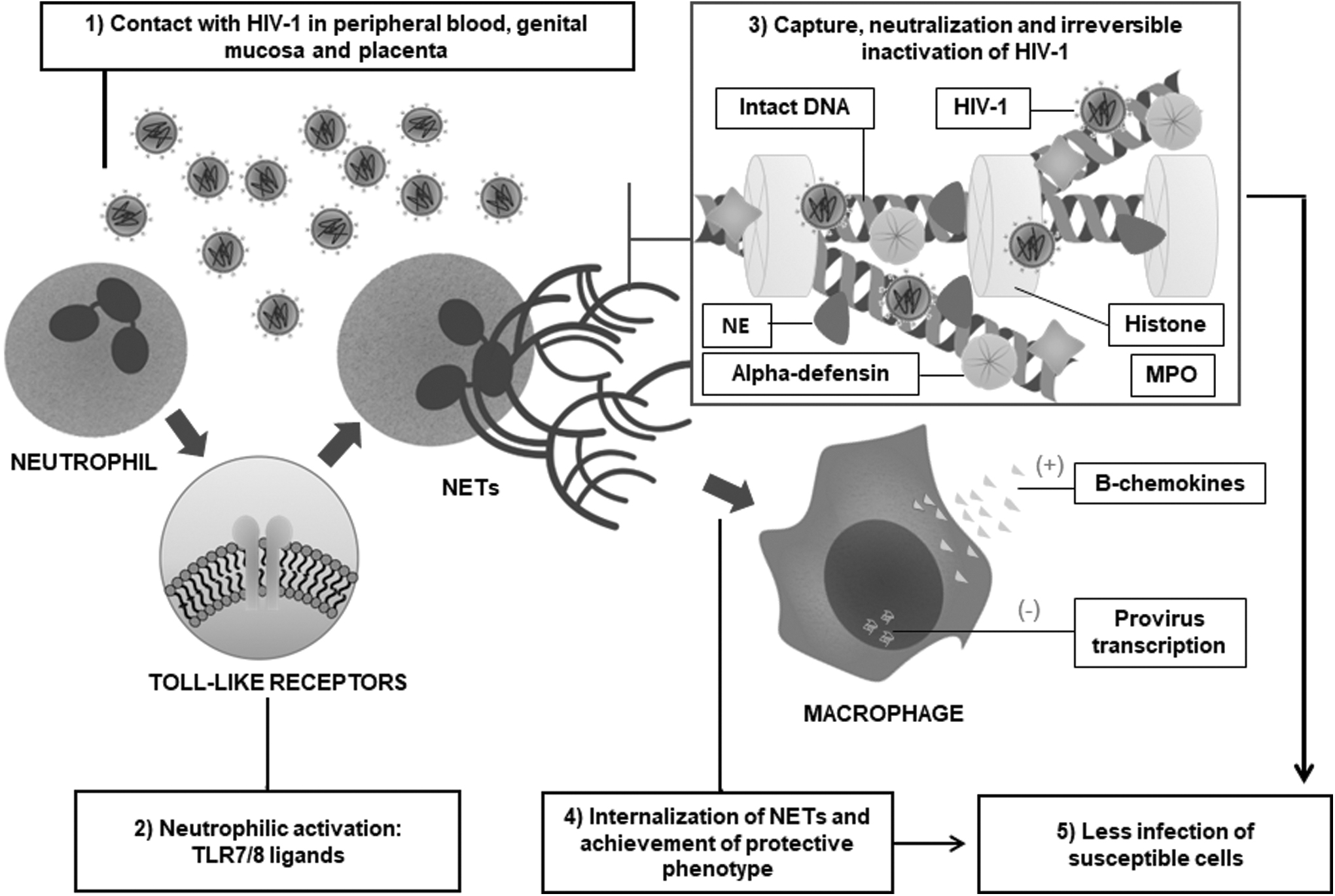
Induction, composition, and effects of NETs against HIV infection. HIV-1, human immunodeficiency virus type 1; NE, neutrophil elastase; NET, neutrophil extracellular trap; TLR7/8, Toll-like receptor 7/8.
Similarly, Saitoh et al. 12 described this process of recognition via TLR 7/8 receptors when performing in vitro assays with peripheral blood neutrophils from healthy individuals exposed to HIV-1 virions. These findings support the results found in experiments with human placentas. According to Moodley et al., 15 PLHIV placentas had a significantly higher number of NETs than the other groups. These studies reveal evidence that neutrophils can recognize HIV and activate intracellular mechanisms that converge to release NETs to combat viral spread. Corroborating this idea, a study showed neutropenia as a risk factor for acquiring HIV. 17
As demonstrated by Giraldo et al., 18 peripheral blood neutrophils also interact with HIV and TLR ligands, modulating the expression of these receptors in cells, regulating the production of Reactive Oxygen Species (ROS), and activating neutrophils. The induction of these receptors in peripheral blood mononuclear cell triggers a response that inhibits HIV replication. This effect is not only achieved exclusively by binding to TLR but also by mechanisms different from those reported in neutrophils, 19 given that the experimental trials show that the neutrophil is activated by nonenveloped HIV-1 even in the face of blockade of receptors 7 and 8. 12
Evidences show that the composition of HIV-1 can modulate neutrophil activation. This is because, in genital tissues, nonenveloped viral particles activate neutrophils dependent on ROS, which does not necessarily happen when exposed to the enveloped virus, whose activation process was independent of the participation of oxidizing agents in the first 30 min. 13 Peripheral blood neutrophils activate when exposed to HIV-1 virions (with or without envelope) in a ROS-dependent manner. 12,18 The presence of this evidence indicates the potential for additional phenotypic variances in neutrophils, beyond those suggested by Smith et al. 20 and discussed in this study. This, in turn, prompts us to formulate hypotheses regarding the existence of alternative signaling pathways. Consequently, we emphasize the necessity for further research to unravel the mechanisms behind neutrophil activation in various tissue locations in the context of HIV infection.
Interestingly, there is evidence that neutrophils also shed their extracellular networks during chronic HIV infection. Mojoli et al. 14 demonstrated this effect when using NET release markers (neutrophil elastase [NE] and extracellular DNA) in the blood of people living with HIV (PLWH) and stimulated it with interleukin 8 (IL-8) in vitro, and verified the release of NETs at different viral loads from undetectable (<40 copies/mL) to high (>100.103 copies/mL). Corroborating these assays, the placentas of HIV+ pregnant women showed significant NETs production measured by immunohistochemistry. 14 It is worth noting that these pregnant women were undergoing Highly Active Antiretroviral Therapy (HAART). Therefore, it is possible to conclude that chronic HIV-1 infection does not extinguish the intrinsic capacity of blood and placental neutrophils to be activated and release extracellular networks rather it is possible that, in the living organism, NETs act as partial protection against the virus dissemination.
It's important to understand that viral activation of NETs is relatively recent, and mechanisms are still being revealed. Most studies in humans are related to respiratory infections. 21 Research regarding HIV in this context is still limited. From what is known, the role of this neutrophil function appears to be dual and pathogen-dependent, probably because the mechanisms underlying NET release differ from one pathology to another. 22 It can act as a protective mechanism to control viral spread and reduce dissemination, or it can favor virus activity, causing extended tissue damage and worsening the infection. 23
From what is known, when exposed to influenza respiratory virus, neutrophils produce NETs in high levels, which are found in patients' plasma. 24 –26 In accordance with this fact, in H7N9 and H1N1 patients, NETs have also been shown to have an impact on the permeability of alveolar epithelial cells, which reflect into acute inflammation. These findings indicate that high levels of NETs correlate with influenza severity and poor prognosis. 26 Despite these findings, a study described that NETs release is triggered only by some specific Influenza strains. While H1N1 stimulates NET production in isolated human neutrophils, the H5N1 strain doesn't. 24 In another scenario, NETs have been associated with immuno-thrombosis and the development of acute lung injury and acute respiratory distress syndrome, which suggests anti-NET therapy as a possibility to avoid exaggerated immune responses, hyperinflammation, immuno-thrombosis, and other complications. 27
There are still few studies that point to the protective role of NETs toward viral infections. Besides the research presented in this review about the importance of NETs in the control of HIV, 12,13 Jenne et al.s' experiments showed that with systemic administration of virus analogs or poxvirus in mice tissue, the release of NETs and the recruitment of both neutrophils and platelets to the liver vasculature, forming large, dynamic aggregates capable of capturing virus particles, protecting host cells from virus infection, and enabling them to be eliminated by macrophages. 22 These findings highlight the role of NETs and early tissuewide responses in preventing viral infection. In accordance with these findings, some viruses present strategies to counter the action of NETs. The dengue virus, for example, impairs NET formation by antagonizing phorbol ester-related glucose uptake. 28
Regulatory Mechanisms of HIV Neutrophil Activation
Objectively, the studies analyzed show solid evidence of the ability of neutrophils to recognize HIV and activate effector mechanisms. However, some constitutive factors can modulate this response (Fig. 2). The microenvironment can influence the innate immune cell's ability to capture HIV. An example of this occurs with neutrophils from the genital mucosa since they can release NETs in a significantly higher amount at the expense of blood neutrophils and capture HIV at a rate four times higher than dendritic cells (DCs). 13 Previously, Smith et al. 20 described the phenotypic differences between genital and peripheral neutrophils in healthy individuals suggesting that the effector response of these cells is influenced by the microenvironment. This finding is reinforced by Barr et al. 13 when exposing evidence of the existence of a site-specific mechanism and self-controlled regulation of neutrophils during viral infection.
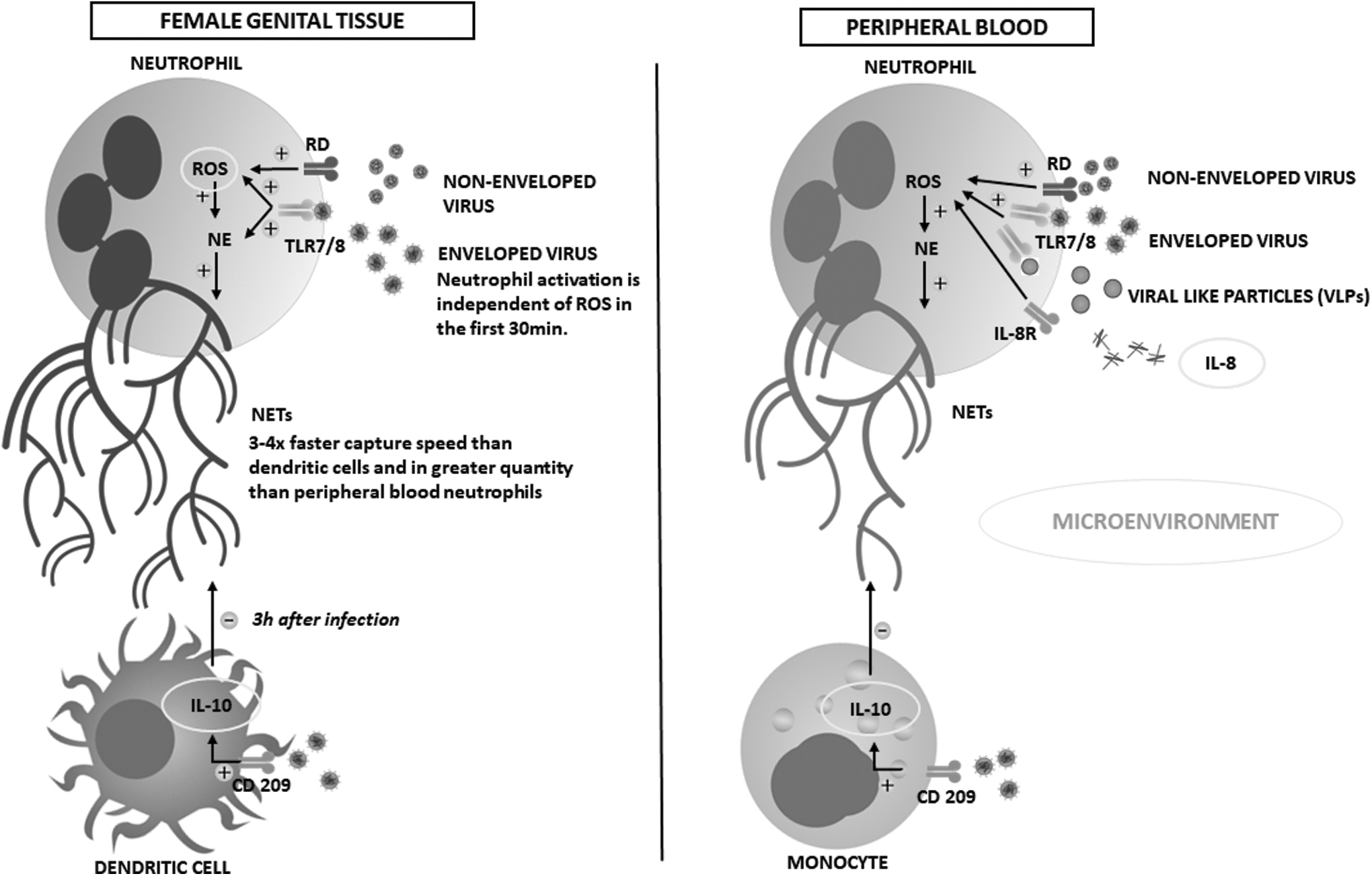
Modulating mechanisms of neutrophil activation and release of NETs after contact with HIV-1 in the genital tract and peripheral blood. Graphic synthesis of the compiled results of the following studies: Refs. 12 –15,17 –20,29 CD209, cluster of differentiation 209; IL, interleukin; ROS, reactive oxygen species; UR, unknown receptor.
HIV itself modifies the microenvironment in which it is inserted, mainly through its mechanisms of immune activation and viral evasion. 29 One of these mechanisms was suggested by Saitoh et al. 12 when verified (by in vitro assays) that HIV can bind to DCs derived from blood monocytes through the cluster of differentiation 2019 (CD209) protein (DC-SIGN), which stimulates the release of interleukin 10 (IL-10), anti-inflammatory cytokine, and inhibitor of NET formation. As a result, the role of DC in HIV viral escape has been extensively studied, in addition to the fact that these cells present antigens to other immune cells, spreading the infection. 30
The IL-10 production mechanism may explain the immunoprotective synergy between preeclampsia and HIV. Theoretically, the greater the amount of extravasated DC at the site of infection, the greater the release of IL-10, decreasing the excess of NETs produced in preeclamptic placentas, thereby reducing mortality as extracellular networks induce a local inflammatory response. 15 However, the experiments by Moodley et al. 15 showed that the group of pregnant patients, HIV+, without preeclampsia, had the highest concentration of NETs in placental tissues, suggesting that other factors counterbalance the performance of IL-10 or that negatively feed DC in this microenvironment.
In other studies, assays with DC from the same genital microenvironment as neutrophils showed that IL-10 release occurred only 3 h after HIV infection, 13,31 evidencing a time gap in which interleukin could not inhibit NETs release. Therefore, the results of these studies converge on the fact that IL-10 is just one piece of this highly complex “puzzle” involving the regulation of NETs production and viral escape. Likely, other interleukins are also involved in this process. An example is IL-8, since its release may stimulate the activation of neutrophils and the production of NETs. 14 Furthermore, recent evidence shows that IL-17, released by neutrophils, can increase the number of target cells for HIV infection and viral spread. 32
In addition to healthy tissues, it is important to investigate the potential of ETs under conditions of homeostatic imbalance, since in certain inflammatory diseases, understanding the participation of NETs is fundamental to elucidating the pathophysiological mechanisms involved. 33 This becomes even more important in a scenario of coexistence between systemic alterations and HIV infection, since interferences may occur in the release of NETs on these occasions, such as in autoimmune diseases induced by Anti-Neutrophilic Antibody Cytoplasmic Antibody (ANCA), in which there is greater production of NETs. 34 In patients with diabetes mellitus, NETs are produced more rapidly, 35 while in preeclampsia, the association with HIV decreases the production of NETs compared to patients living with HIV without preeclampsia. 15 Based on this evidence, we suggest that the presence of systemic alterations in PLHIV can modulate the response of neutrophils, especially in diseases that cause depletion, morphological, and/or functional alteration of these cells.
Furthermore, studies suggest the relationship of NETs with endothelial activation and development of atherosclerosis, 36 as well as with metastatic processes. 37 Cardiovascular diseases and cancer are the leading causes of death. 38 This scenario is worrying given the proposal to use NETs for therapeutic purposes in PLHIV who have concomitant heart disease or malignant neoplasms, given their potential to worsen the prognosis. Thus, future therapies aiming at modulating the expression of NETs may present positive or negative results depending on the systemic conditions of the individuals.
The Anti-HIV Potential of NETs Components
Neutrophils, during HIV infection, carry out the capture, neutralization, and inactivation of the virus. The mechanisms involved in these steps are complex and not fully understood. What is known is that the network of decondensed DNA, which makes up the NETs, forms a mechanical barrier that allows the capture of the virus. 12 In this first step, the NETs must manage to keep the virus fixed to neutralize it, and, for this, the NETs DNA must be intact. 12 –14
Although other factors may be involved, the neutralization of HIV-1 by NETs is due to the histones present in their composition, since they are proteins rich in amino acids (positively charged) and which, by electrostatic forces, can attract and trap virions. 23 Saitoh et al. 12 showed a great contribution of histone H3 in the capture, neutralization, and inactivation of HIV-1. These authors suggest that histones 3 and 4 have this antiviral activity, unlike histones 1 and A2. This diverges from the findings of Moodley et al. 15 since these authors identified high expression of H2A and its immuno-localizing potential of NETs. In addition, Kozlowski et al. 39 showed a greater inhibitory response to HIV-1 in recombinant human histones H1 and H2A. This may indicate the influence of the neutrophil phenotype on the HIV-1 capture efficiency according to the NETs composition.
During the viral inactivation stage, neutrophilic granules enzymes, components of NETs, actively fight HIV. These enzymes are myeloperoxidase, 12,14 α-defensins, 12 and NE. 14 The latter maintains its potential to inactivate HIV-1 even when isolated from DNA, unlike the others. 14 Based on these results, the anti-HIV-1 activity from several other proteins from NETs, such as defensins, cathelicidin, calprotectin, and actin 33,40 –42 have been described. Given the methodological variety of the studies, it is possible to assume that different stimuli and different populations of neutrophils induce, through different biochemical pathways, the proteins that form NETs.
Based on this evidence, we conclude that the anti-HIV properties presented by NETs are intrinsic to their constituents and that, therefore, these structures have the biological potential to prevent the spread of the virus in vivo. However, changes in this process and the involvement of other stimuli can unbalance the antiviral effect, contributing to the mechanisms of viral evasion and infection of target cells.
The Immunoprotective Role of NETs in Target Cell Infection
During HIV infection, one of the biggest challenges is to prevent the compromise of target cells. In vitro assays demonstrated that, upon exposure to HIV-1, the coculture of neutrophils and CD4+ T cells resulted in a lower number of infected lymphocytes than the control group. 12,13 In primary macrophages, the effect of NETs appears to be even more significant, since the phagocytosis capacity of these cells allows the internalization of extracellular networks 43 and consequent acquisition of an antiviral phenotype, through the inhibition of transcription of the provirus and increased production of β-chemokines, which allows for a lower release of viral particles and less infectivity. 14 Therefore, the NETs ability to prevent virus spread is due to their intrinsic antiviral effect, capable of directly killing HIV, and aiding in the defense of macrophages, since after NETs are internalized, they respond to HIV-1 through effector mechanisms that confer immunoprotection of other susceptible cells.
On the other hand, in most cases, viral escape mechanisms overlap with those of immune attack 21 due to several factors, including the ability of activated neutrophils to recruit target cells to the site of infection by the release of cytokines. 32 Since HIV replication is the key to the whole process, it is important to emphasize this duality regarding the role of activated neutrophils to prevent the infectivity of the virus while attracting new susceptible cells to the site of infection, facilitating the natural evolution of the infection, with the dissemination of HIV, development of high viral load in the body, and AIDS status.
As the evidence points to the beneficial activity of NETs toward HIV infection, it is possible to consider this event as a therapeutic approach. However, because of the protagonist role of NETs in chronic inflammation and tissue damage, most efforts are made to discover ways to inhibit it by targeting important components from different pathways that trigger the formation of NETs. 27 A variety of endogenous and exogenous inhibitors have been discovered, such as neonatal NET-inhibitory factor; prostaglandins, like PGE2, which can inhibit NETosis and NET formation through the activation of protein kinase A and cAMP 44 ; the activated protein C inhibits the release of extracellular histone, activation of clotting factors, and NETs accumulation and formation 45 ; antibiotics may also have immunomodulating effects through the suppression of NETosis and NET formation and the release of pro-inflammatory cytokines. 46 Even aspirin inhibits NETosis, platelet activation, and NET formation. 47
However, alternative therapies able to enhance NET formation are slowly emerging for particular situations where NETs are beneficial. In an effort to increase neutrophil activity in cases of chronic granulomatous disease, gene therapy was applied to complement the function of NADPH oxidase, allowing neutrophils to produce NETs and control bacterial infections, a major cause of death for these patients. 48 Further studies must be conducted to better understand how to selectively and adequately promote local or general NET formation without exacerbating the neutrophil response, thus avoiding the establishment of chronic inflammation.
NETs and Their Potential Implications in HIV Associated Comorbidities
It's important to take into consideration that, with the success and effectiveness of antiretroviral therapy associated with the increase in HIV screening and improvement of access to adequate therapy, PLWH now have a better life expectancy and reduced deadly comorbidities. 49 Nevertheless, a substantial number of those individuals are affected by a variety of chronic health issues, which can be either directly or indirectly related to HIV infection and its treatment. 50 Premature aging is noticed, leading to age-related comorbidities at a younger age when compared to non-HIV-infected people, 51,52 possibly due to HIV infection and the side effects of antiretroviral therapy. 53 –55 Also, noninfectious comorbidities account for more than half of deaths in PLHIV, led by cardiovascular diseases and cancer. 56 –58 Other frequent comorbidities are hypertension, diabetes, pulmonary diseases, chronic renal diseases, and osteoporotic bone fractures. 58 –62
Therefore, recent research aims to increase the knowledge about specialized immunoprotective functions of NETs and also appoints their pathophysiological role in many diseases, contributing to acute or chronic pathogenesis mostly by the establishment of chronic inflammation. 63 In cardiovascular diseases, one of the most important type of comorbidities in PLWH, NETs can be associated with a large amount of conditions, such as atherosclerosis, 64 acute coronary syndrome, 65 and many thrombosis-related conditions. 66 Other important complications associated with this neutrophil function are diabetes, 67 –69 respiratory diseases, 70 chronical renal conditions, 71,72 and some type of cancers. 73,74
As we can see, despite the evidence that points to the important role of NETs in HIV infection control and virus replication, the exacerbated production of NETs in PLWH can be an aggravating factor for other conditions, such as the most frequent comorbidities, and must be evaluated and taken into consideration to prevent worse outcomes.
Limitations
Moodley et al. 15 found an immunoprotective synergy between HIV infection and preeclampsia conditions that alone cause high mortality but when together offer immunoprotection and lower mortality. However, it was not possible to clarify which mechanisms modulate this synergy.
Saitoh et al. 12 sought to assess the potential for the interference of histone proteins on the infectivity of HIV-1 virions, but they only analyzed the role of H1, H2, and H3 histones. Although they did not show negative results for histones H1 and H2, they then suggested the antiviral role of histone H4 without neutralization mediated by monoclonal antibodies.
In the research by Barr et al., 13 a limiting factor was the small number of samples in some experimental trials, with a total of six participants; in some assays, the number of tissue samples was lower. Finally, in all studies, there was a need to better investigate the release and microbicidal capacity of NETs derived from different PLHIV cohorts, as suggested by Mojoli et al. 14
Despite the limitations, the analyzed studies evoke advances related to the immunoprotective role of NETs in the infection of target cells by HIV-1, which favors future therapeutic approaches that involve the modulation of the neutrophil effector response. NETs appear to play at least a partial role in containing primary HIV infection in the body, so the approach of conferring an immunoprotective phenotype on cells to prevent viral replication in susceptible individuals may have benefits that outweigh their deleterious potential to tissues. NETs may also have prognostic value since it is possible that their concentration is related to the decrease in viral load and so could be used as biomarkers in the therapeutic course of the patient. These methods may contribute to preventing HIV infection in vulnerable people, reducing vertical transmission, and treating infected patients by reducing the viral load.
Conclusions and Future Perspectives
In vitro and ex vivo experiments show that NETs from different body sites neutrophils can act against HIV-1 infection by capturing, neutralizing, and inactivating the virus. In addition, NETs prevent other cells' infection and provide a protective phenotype in macrophages once they phagocytose HIV-associated NETs. These processes decrease infection spread to the virus target cells (TCD4 lymphocytes). However, these mechanisms are influenced by the tissue's physiological conditions, the cell phenotype, and the virus, making the antiviral action of NETs variable. Despite the small number, the studies have a low risk of bias and good methodological quality, suggesting reasonable reliability in the body of evidence pooled here.
Future research can focus on and clarify the mechanisms of regulation of the production of NETs of different populations of neutrophils, deepening their relationship with HIV. In addition, studies should consider the deleterious potential of NETs evidenced in several pathologies. These issues raise the need for in vivo studies, given that the theoretical basis for the effectiveness of NETs against HIV infection is based on in vitro studies.
Footnotes
Authors' Contributions
N.M.S., N.P.M.L. and M.F.A. conceived and designed the study. N.M.S., I.K.S., T.A.A.M.F. and M.F.A. established the search strategy. N.M.S., I.K.S., J.R.L.P.C., T.A.A.M.F. and M.F.A. contributed software/analysis tools. N.M.S., N.P.M.L., A.E.C., V.D.A. and E.G.C.N. extracted the data. N.M.S., N.P.M.L., J.R.L.P.C., E.G.C.N. and M.F.A. perform the analysis. N.M.S., N.P.M.L., A.E.C., V.D.A., I.K.S., J.R.L.P.C., T.A.A.M.F., E.G.C.N. and M.F.A. wrote the article. All the authors read the manuscript before they have given the final approval for publication.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
The study was funded from internal resources.