
Abstract
Since the first HIV-cured person was reported in 2009, a strong interest in developing highly sensitive HIV and SIV reservoir assays has emerged. In particular, the question arose about the comparative value of state-of-the-art assays to measure and characterize the HIV reservoir, and how these assays can be applied to accurately detect changes in the reservoir during efforts to develop a cure for HIV infection. Second, it is important to consider the impact on the outcome of clinical trials if these relatively new HIV reservoir assays are incorporated into clinical trial endpoints and/or used for clinical decision-making. To understand the advantages and limitations and the regulatory implications of HIV reservoir assays, the National Institute of Allergy and Infectious Diseases (NIAID) sponsored and convened a meeting on September 16, 2022, to discuss the state of knowledge concerning these questions and best practices for selecting HIV reservoir assays for a particular research question or clinical trial protocol.
Workshop Goals
Robert F. Siliciano (Johns Hopkins University) centered his introduction of preclinical workshop goals around the question, “HIV reservoirs: what to measure and when.” A major barrier to cure is a small pool of resting CD4+ T cells that harbor latent HIV. 1 Virus remains latent in these cells because inducible host transcription factors required for HIV gene expression including NF-κB, NFAT, and P-TEF-b are sequestered in the cytoplasm or in inactive complexes. Activation through T cell receptors can reverse latency and lead to viral rebound if treatment has been stopped but it can also lead to proliferation without virus production, generating new latently infected cells. Viral reservoir measurements should not only include productively infected cells, but also proviruses with the potential to produce progeny, as detected in assays, such as the Quantitative Viral Outgrowth Assay (QVOA), Intact Proviral DNA Assay (IPDA), or Quadruplex (Q4)-PCR.
The QVOA provides only a minimal estimate of the reservoir size. Some latent proviruses need more than one round of stimulation for induction. Repeated stimulation with antigen in vivo can lead to the formation of large clones; this has been shown most clearly with cytomegalovirus as a driver of clonal expansion. However, viral rebound can also come from minor, undetected variants because only some viruses in the reservoir can replicate in the presence of autologous neutralizing antibodies (aNAbs). 2 Thus, rebound involves a complex interaction between viruses in the reservoir and the immune response, particularly aNAbs.
This interaction is not captured by any reservoir assay, except the QVOA performed in the presence of aNAbs. Many studies have emphasized the importance of particular T cells subsets of CD4+ T cells (CD32+, Th1, Th17, Tfh, Tscm, central, transitional and effector memory cells, and naive T cells, activated and cytolytic T cells) where HIV is enriched, but there is no clear scientific basis for this claim because only CD4 and coreceptor are required for entry. The Siliciano laboratory has shown that replication-competent virus is present in all memory subsets with more variation between individuals than between subsets. The most important aspect of reservoir assays is distinguishing between intact and defective viruses. Studies by Ya-Chi Ho established that most proviruses persisting in treated patients are highly defective owing to large internal deletions or APOBEC-mediated hypermutation. 3 Single-amplicon PCR assays capture mostly defective proviruses. These findings were the motivation for the development of the IPDA, which provides a better quantitative estimate of intact proviruses. 4
The time points when to make HIV reservoir measurements are also important. The decay of the latent reservoir in resting CD4 cells in the first decade of treatment is slow with a half-life of 44 months, as shown independently by two laboratories. Detailed studies demonstrated that the decay of intact proviruses is not linear and includes four phases with three inflection points at 3 months, 2–3 years, and 7 years. However, in patients on suppressive antiretroviral therapy (ART) for more than 7 years, subsequent reservoir decay occurs more slowly as confirmed by the QVOA and IPDA measurements. Dr. Siliciano also presented decay curves for SIV and SHIV.
Many cure studies in nonhuman primate (NHP) models are conducted early when the viruses are still decaying relatively rapidly on their own and therefore, adequate control populations are necessary. Dr. Siliciano concluded that reservoir measurements should focus on latent, replication-competent proviruses, which represent the most challenging barrier to cure. Latently infected cells can proliferate without producing virus, generating large clones, but rebound can come from minor variants and any CD4 T cell subset. Decay processes complicate reservoir measurements during the first 2 years of ART. Decay processes, selection processes, and infected cell proliferation alter reservoir composition over time, but readily inducible, replication-competent proviruses are present and not declining after more than 20 years of suppressive ART.
Validation of a surrogate marker for ART-free remission will be complex because curative interventions may vary in their mechanisms of achieving ART-free remission. Therefore, each type of intervention may need its own set of biomarkers that require independent validation as surrogate endpoints. Candidate biomarkers are either virological (HIV nucleic acid, inducible virus production) or immunological (CLT, NK, antibody, immune activation, exhaustion markers, etc.) in nature.
Preclinical Session
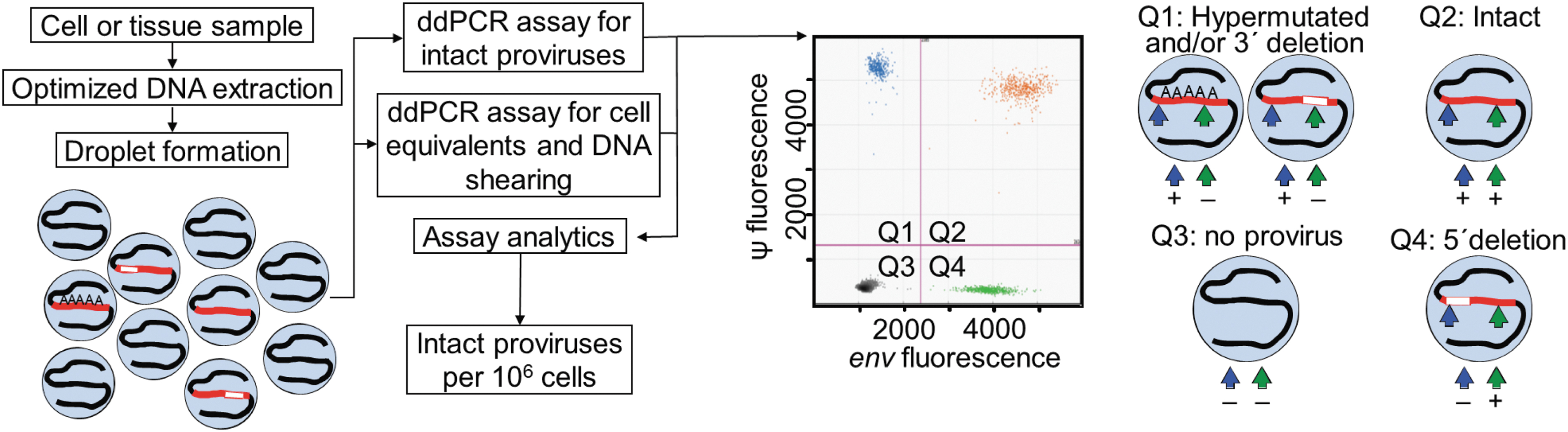
IPDA. DNA is extracted from cell or tissue samples and subjected to digital droplet PCR for intact proviruses using amplicons of the Ψ and env regions. Proviruses that have a 3′ deletion and/or are hypermutated are displayed in the upper left quadrant of the IPDA dot plot (Q1), proviruses that have a 5′ deletion in the lower right quadrant (Q4), and intact proviruses in the upper right quadrant (Q2). IPDA, Intact Proviral DNA Assay; PCR, polymerase chain reaction.
Optimal amplicon placement for other clades may be different and requires analysis of hundreds of full-genome sequences. The impact of DNA shearing on HIV quantification is corrected by amplifying a host gene (RPP30) using two amplicons with the same spacing. The IPDA correlates with QVOA results in a statistically significant manner, although the IPDA gives higher values because it detects proviruses that are not induced in the QVOA [median ratio of intact proviruses/infectious units per million (IUPM) is 100]. Problems with the assay are that some polymorphisms can preclude amplification in ∼7% of the samples, which can be corrected with alternative primers. 7 Second, some defective proviruses with minor deletions are given in the upper right quadrant (Q2, 10%–15%), where intact proviruses appear (Fig. 1). In addition, the IPDA also does not provide information on HIV sequence or clonality.
Other approaches to reservoir measurement involve near-full genome sequencing (nFGS). These include assays such as FLIPS, FLIP-seq, and Q4-PCR, which all use nested PCR to determine the sequence of individual proviruses. In a comprehensive study, Dr. Siliciano evaluated six different published nFGS methods. 8 The outer PCR common to these methods fails frequently and only detects one-third of intact proviruses measured by the IPDA. In addition, less sensitive detection methods are used, such as gel electrophoresis and Q-PCR, and they do not correct for DNA shearing. Although these assays provide ample qualitative information, caution should be used in drawing quantitative conclusions. In conclusion, the IPDA is a scalable assay that allows direct digital counting of intact proviruses.
The IPDA distinguishes populations of proviruses with different in vivo decay rates and susceptibility to shock and kill strategies. The IPDA does not provide information on inducibility, nucleotide sequencing, or clonality. Finally, the QVOA and related induction assays provide a definite minimal estimate of reservoir size but miss a substantial fraction of intact proviruses owing to the stochastic nature of induction. nFGS methods provide information on proviral intactness and clonality but are not quantitative and miss most intact proviruses. The use of multiple assays provides the clearest picture of the reservoir.
As already mentioned, the Q4-PCR measures a lower provirus number than the IPDA, owing to a lower amplification efficiency. The half-life for intact proviruses is 4.8 years and for defective proviruses >50 years, which is in agreement with previous publications. The assay provides additional information on clonality of the reservoir. It was shown that the reservoir increased clonality in both the intact and defective proviruses and decreased complexity over time. 16 One needs to be cognizant that misclassified defective proviruses can artificially extend the half-life of intact proviruses. In a small set of samples, the results of the Q4-PCR and IPDA did not agree; in such cases, the therapeutic effect of an intervention could be underestimated, which is very relevant for small interventional trials. In summary, the IPDA is a rapid high-throughput assay, but cannot fully guarantee the intactness of proviruses.
Q4-PCR is low throughput and far less efficient than the IPDA. The half-life of the intact reservoir measured by Q4-PCR is the same as for QVOA and IPDA in the initial 7 years after ART initiation. The defective reservoir has a much longer half-life than the intact reservoir. Using two amplicons based on IPDA there is a significant and variable misclassification rate of intact proviruses, which may be less sensitive to changes in the intact reservoir. This is especially important in small studies and over relatively short time intervals, such as 1–2 years. The recommendation, therefore, is that for small studies or whenever possible, both digital droplet PCR (ddPCR) and sequence-based assays should be performed.
Of note, the number of identified proviral clones increases steadily as T cells mature. This finding made it clear that naive T cells were truly infected as memory contamination was ruled out because of the distinct sequences identified in naive T cells. To emphasize this point, Dr. O'Doherty presented one example where clonality steadily increased from naive (6% clones) to effector memory (85% clones). In addition, five of the eight largest clones in the same patient originated from naive T cells suggesting the naive reservoir repopulated the memory reservoir. Her team also revealed that the level of naive infection appeared to predict HIV reservoir diversity more accurately than the other subsets. This conclusion was surprising because naive T cells have less HIV DNA than memory cells. 19
Naive T cells have attributes that could make them a formidable reservoir, such as their long half-life, and their resistance to CTL killing. In summary, the strengths of longitudinal sequencing include the ability to identify large clones, and the ability to define the contribution of individual subsets. Limitations include high cost, need for large cell numbers, and slow throughput. Eliminating latently infected naive CD4 T cells will be critical for HIV cure because they constitute a sheltered reservoir, repopulate memory cells, and are long-lived.
Intraindividual polymorphisms can also lead to partial detection failures making the reservoir appear smaller than in reality. These detection failures have more weight in small clinical trials where sequencing of individual samples may be feasible and warranted. He applied some of these approaches to the eCLEAR study in Denmark, 21 which had an enormous diversity of subtypes with 51% not being subtype B (A, C, D, F1, AE, and AG). Therefore, he applied duplex digital droplet 3D PCR by adjusting the nucleotide sequence of secondary primers and probes of the env region, which does not detect hypermutated proviruses. Overall, his laboratory developed 17 custom primers and 25 custom probes for the eCLEAR study. The IPDA correlated well with 3D PCR in individuals where IPDA primers bound to the HIV sequence.
They examined 2,400 full-length HIV sequences (171 subtype A, 1179 subtype B, 719 subtype C, 71 subtype D, 292 CRF01-AE) and identified specific nucleotide positions in which primer/probe binding sites differed in >10% of sequences for any given subtype. In an iterative process, they changed up to six sites per primer, prioritizing the three bases at the 3′ end and at most one position per probe. 24 However, they could not find a primer/probe set for tat that reliably quantified subtypes B and CRF01_AE. Therefore, they chose to move forward with a three-target assay (LTR/gag, 5′pol, and env—Assay 2 of the 5T IPDA), termed CS-IPDA. 24 The CS-IPDA has excellent agreement with absolute quantification by independent gag quantitative PCR (qPCR) assay using JLAT 5A8 cells.
The assay is very sensitive (the absolute limit of detection is 1–3 copies per reaction), and 0/34 negative controls were positive by CS-IPDA. In silico analysis shows good agreement with computational inference of intactness (Pro-Seq-IT) and provides a sense of overestimation of intactness (9.6 for subtype B and 15.5% for subtype C) by CS-IPDA compared with full-length proviral sequencing. The team applied this assay to samples from infants living with HIV in Kenya. 24 In summary, IPDAs are an effective way to estimate the intact HIV reservoir, and thus play an important role in HIV cure studies. The first generation of IPDAs were designed for subtype B and performed poorly on non-B subtypes. The cross-subtype IPDA (CS-IPDA) performs well on all subtypes (including subtype B). CS-IPDAs are already contributing to the study of HIV biology and cure interventions in regions most seriously impacted by HIV.
To develop the assay a cohort of 12 participants was examined by standard QVOA and dQVOA. For the standard QVOA, CD4 T cells from cryopreserved peripheral blood mononuclear cells (PBMCs) were plated in a limiting dilution format in the presence of PHA and gamma-irradiated allogeneic PBMCs and cocultured with allogeneic lymphoblasts at a 1:4 producer-to-target ratio. The dQVOA starts with 50–100 million PBMCs that are cultured in limiting dilution in a differentiation medium (IL-6, IL-7, IL-10, IL-15, TNF-α), which will transition the cells into an effector memory phenotype. After differentiation, the cells are activated with PHA and gamma-irradiated allogeneic PBMCs without redistribution of the cells. Because of the differentiation, the participants' own CD4 T cells are the ideal targets for expansion. Therefore, target lymphoblasts do not need to be added and passaging of cells is not required during the culture.
The IUPM resting CD4 cells using cell dilution and positive HIV-1 p24 frequency are calculated using the maximum likelihood method. Dr. Kulpa found that dQVOA enhanced the sensitivity of viral outgrowth assay; ex vivo differentiation of resting CD4+ T cells followed by mitogen activation significantly increases HIV reservoir measurements over activation alone. dQVOA also allowed for IUPM measurements in two participants that were unmeasurable with the standard QVOA assay. For assay validation, a blinded panel was provided by RAVEN, which demonstrated that the data were reproducible. Evaluation of participants on short (1.8 years), medium (4.3 years), and extended (12.1 years) treatment demonstrated that long-term ART may not result in a smaller reservoir size, but instead CD4 T cells from long-term suppressed PLWH require additional signaling for effective viral reactivation. In summary, the advantages of the dQVOA are more efficient reactivation, major reagent and labor cost savings and that fewer patient cells are required.
The second assay is the matched integration site and proviral sequencing (MIP-Seq) assay. 31 The assay can locate the chromosomal position of genome-intact proviruses and permits to assess their chromosomal location relative to transcriptional and epigenetic features in the host genome. He also proposed that integration sites can be used as a biomarker of selection of viral reservoir cells. In elite controllers, proviruses are integrated into very distinct regions of the genome (heterochromatin). In one person on long-term ART, there was a marked transformation of the proviral integration site profile over time, with a progressive accumulation of intact proviruses in centromeric satellite DNA. 29 Defective proviruses did not undergo the same selection process. The third assay is the parallel HIV RNA, Integration Site, and Proviral Sequencing (PRIP-Seq) assay. 32 The assay is a combination of the MIP-Seq assay with separate HIV RNA extraction and amplification of HIV-1 RNA transcripts.
With this assay, he can determine whether clones are transcriptionally active or silent. He analyzed 701 reservoir cells and found that proviruses integrated in genic regions are more transcriptionally active (∼35%) than in nongenic regions (∼20%) or satellite DNA (∼5%). 32 In summary, the PRIP-Seq assay evaluates transcriptional behavior of single proviruses, reflects the susceptibility of proviruses to in vivo viral reactivation signals, and evaluates the “depth” of latency. The fourth assay, PheP-Seq focuses on the phenotype of the viral reservoir cells. Memory CD4 T cells are isolated from patients, stained with oligonucleotide-tagged antibodies, followed by single-cell encapsulation, single-cell barcoding and multiplex single-cell PCR to amplify strategically important regions of HIV-1 DNA in conjunction with amplification of antibody tags.
Overall, he used 53 selected surface markers and 2 isotype controls, and 18 HIV-1 DNA fragments totaling 4,080 bp, which allowed for a detailed phenotypic analysis and examination of individual clones. In summary, the PheP-Seq assay can evaluate the surface phenotype of infected cells directly ex vivo, can distinguish the phenotype of cells harboring intact versus defective proviruses, can simultaneously analyze >50 markers, and can evaluate the phenotype of individual infected cell clones defined by chromosomal integration sites.
Clones of HIV-infected cells were, in general, much more stable than were clones of uninfected cells taken from the same donors at the same time points. 34 Because the clones of infected cells were derived from cells that were infected before therapy was initiated, all the infected clones are old (>9 years). Even after >9 years on ART, clones of infected cells that are able to produce infectious viruses can grow either larger or smaller over an interval of 2–3 years. The size distribution of the largest clones of infected and uninfected cells was smooth and continuous for the three donors studied. However, the largest clones of uninfected cells were larger than the largest infected clones. For infected cells, the primary negative selection is against cells that have proviruses integrated into highly expressed genes; this selection is stronger for cells in which the proviruses are in the same orientation as the gene.
There is a positive selection for cells with a provirus integrated in particular introns of seven oncogenes: BACH2, STAT5B, MKL2, MKL1, IL2RB, MYB, and POU2F1. The primary mechanism of oncogene activation appears to be promoter insertion. Oncogene activation accounts for only a small fraction (2%–3%) of the clones of HIV-infected cells. The very limited data thus far suggest that most or all of these HIV proviruses integrated in these seven oncogenes are defective, suggesting that this mechanism does not contribute to the reservoir.
There is no difference in the fraction of infected cells with unspliced HIV RNA in noncontrollers, viremic controllers, and donors on ART. The fraction of infected cells with transcriptionally active proviruses is not different in PBMCs and lymph node cells in nonviremic individuals suppressed on ART (∼10% in both). 35 Between 1% and 60% of infected cells within T cell clones have transcriptionally active proviruses. 36
The samples from different patients are barcoded, pooled, and sequenced simultaneously. This approach is efficient with respect to time and sequencing costs and allows accurate comparisons because input cDNA levels are effectively normalized. To reverse latency, transcription and elongation factors have to be activated and epigenetic blockage has to be removed. Because the Tat protein drives cells to maximal transcription, RNA levels are roughly proportional to inducible cell numbers. However, one of the challenges is to establish an accurate calibration curve to convert RNA read numbers into cell equivalents, and this is best achieved using limiting dilution assays. EDITS positive signals roughly correlate with IPDA estimates although the IPDA signals have a wider spread than the EDITS signals.
In this assay, the reservoir in women was slightly smaller than in men (p = .003). In another application, it was shown that women undergo an expansion of the HIV reservoir size during menopause. Similarly, in Ugandan PLWH (clade A1, C, D), the HIV reservoir sizes are smaller than with clade B in the United States but genetically more diverse. 38 Dr. Karn is also developing an EDITS-style assay for SIV barcoded viruses in collaboration with Brandon F. Keele. Perhaps the greatest limitation of the assay is cytotoxicity during induction, which can be very significant for some donors. Detailed validation parameters are available upon request.
Those observations demonstrate that despite plasma viral suppression and early treatment initiation, vRNA+ CD4+ T cells were found in a third of participants while on ART and detected in all at ATI, reinforcing the importance of the GALT as essential site of viral replication during ART interruption. She also collaborated with Dr. Mirko Paiardini, where in situ analysis of the lymph nodes of suppressed NHP helped to identify CTLA4+ PD1− memory CD4 T cells as major contributors to viral persistence. 40 Laser capture microdissection combines laser capture and laser cutting and allows to specifically select cells or regions of interest to then extract DNA (RNA) from formalin-fixed paraffin-embedded or frozen tissues.
More recently, her laboratory adapted an improved assay that overcomes some of the limitations of classic immunohistochemistry named the PhenoCycler-Fusion system (AKA Codex) from Akoya Biosciences. This new approach using barcoded antibody, allows for detection of >40 biomarkers while maintaining tissue integrity, and 70 antibodies for NHPs have been validated. In addition, applying artificial intelligence to tissue analysis, the Tissue Analysis Core in collaboration with the Advanced Biomedical Computing Center at the Frederick National Laboratory, developed a tool called HistomicsK, which allows accurate quantification of RNAscope and DNAscope signals as well as counting of CD4+ T cells with the goal to express the number of virion-infected cells and vDNA+ cells per million of CD4+ T cells.
Furthermore, animal barcode distributions are remarkably consistent over time with the log barcode proportion on day 7 postinfection (pi) highly correlated with day 9 pi. In addition, barcodes identified in various tissue compartments are nearly always found in plasma as well. Only 1.3% of barcodes found in any tissue were not also detectable in blood plasma. The in vivo growth rates are similar even if the challenge doses vary (500–200,000 IU) but the number of detectable barcodes increases with increasing challenge doses. Dr. Keele found that if ART is initiated early, the decay rates are faster compared with when ART is initiated later. Important viral reservoir research questions can be uniquely addressed using barcoded virus in nonhuman primates, for example, the pretherapy phase, decay during ART, reservoir changes over time, clonal expansion during the maintenance phase, and reactivation and rebound.
Dr. Keele reiterated that the reactivation rate is the average time between events with sufficient progeny to be detectable in blood plasma. Individual reactivation events can be readily detected, and the reactivation rate can be calculated using their relative proportion, which is a surrogate marker for the reservoir size. Dr. Keele is currently barcoding some HIV-transmitted founder (T/F) viruses for use in vitro and in humanized mouse models. New viral models containing an antigen tag, including fluorescently tagged and barcoded viruses are also in development.
Clinical Session
In 2014, an improved single-copy assay (iSCA) for quantification of persistent HIV-1 viremia was developed. 47 iSCA improves HIV RNA detection in test panels of donor plasma. RNA extraction from 20 mL of plasma results in a detection limit of <1 HIV-1 RNA copy per milliter. In five donors with HIV-1 RNA below the limit of detection with standard iSCA, four of five became detectable with the mega-iSCA. 47 The iSCA assay was then further improved (iSCAv2) with the 95% limit of detection being 1 copy per 5 mL of plasma. 48 Automation of the SCA was then accomplished through the Hologic Panther platform by testing nine replicates of 0.5 mL of plasma (9 × Panther). For clinical samples, 9 × Panther was more sensitive than iSCAv2. Residual viremia measured by iSCA v2 correlated with IPDA after long-term ART (median 7.1 years). 49 In the PENNVAX study, the Panther 9 × assay in the placebo arm showed that the levels of residual viremia were much lower when ART is given in acute versus chronic infection.
Jonathan Z. Li 50 also found that higher residual viremia was associated with shorter time to rebound. In regard to the clinical trial A5345, iSCA was the best predictor of HIV rebound (vs. CA-RNA, total DNA, IPDA, and QVOA). Collectively, these studies demonstrate that measuring persistent viremia with a high-throughput SCA is promising for assessing the impact of experimental interventions of the expressed HIV-1 reservoir (Table 1).
Low-Level Viremia Assays
ART, antiretroviral therapy; IPDA, Intact Proviral DNA Assay.
Popular Assays to Measure the HIV Reservoir in Clinical Studies
ddPCR, digital droplet PCR; dQVOA, differentiation Quantitative Viral Outgrowth Assay; EDITS, envelope detection by induced transcription-based sequencing; qPCR, quantitative PCR.
In addition, it is not clear which assays are best able to predict the timing of HIV rebound after treatment interruption (Tables 3 and 4). The identification of biomarkers to predict time to HIV rebound will be important in developing and evaluating promising treatment strategies. The A5345 clinical trial aims to determine associations between virologic, immunologic, and host biomarkers and time to rebound. 52 There was a modest delay in viral rebound in early treated individuals who remained off ART for a longer period of time. Laboratory virology assays used were unspliced cell-associated HIV RNA (CA-RNA), total cell-associated HIV DNA (CA-DNA), intact proviral DNA (IPD) assay (Accelevir), residual viremia by the iSCA, and infectious units/million rCD4 cells (IUPM) by the dQVOA.
Key Limitations of HIV Reservoir Assays
Assays Used in Recent and Current HIV Cure Trials
Also referred to as double-positive HIV-1 provirus assay
In addition, antibody levels (HIV-1 + 2 Ab, HIV Combo Ab, LAg-Avidity) were measured by Dr. Michael P. Busch. In A5345, smaller size and lower activity of the HIV reservoir predicted a modest delay in HIV rebound, but the strength of the association was dependent on the timing of ART initiation. The IPDA was the strongest predictor of viral rebound time in chronic-treated and iSCA in early-treated participants although the number of participants was low. Systematic evaluation of reservoir assays and predictors of viral rebound timing is needed.
Two cycles of VRC07-523-LS infusion each followed by 10 doses of Vorinostat were administered to ART-suppressed participants. 55 At baseline and at the end of the study, QVOA, IPDA, and caRNA measurements were performed on resting CD4+ T cells. In three of eight participants, there was a downward trend in IPDA, QVOA, and ca gag RNA. Then, she presented preliminary data from the XTRA study where HIV-specific, ex vivo–expanded T cells were infused in combination with Vorinostat. The primary objective was to evaluate the safety of autologous, ex vivo–expanded HIV-1–specific T cells (HXTC) therapy followed by serial administration of Vorinostat in participants maintained on suppressive combination ART and to evaluate the association of serial Vorinostat dosing and serial HXTC therapy on the frequency of resting CD4 T cell infection (IUPM) via the QVOA and IPDA in participants.
The secondary objective was to explore the ability of combination Vorinostat and HXTC therapy to increase HIV-1–specific immune responses in participants maintained on suppressive ART. In three participants, there was a trend in decline in IPDA and in two participants, a decline in QVOA measurements. Longitudinal analyses confirm small declines in viral outgrowth and intact proviruses, but not in defective proviruses. In summary, successful changes may not have a 1-log10 effect, but these measurable changes may provide important information about the intervention, for instance, whether intensified version of the approach should be explored. Different types of reservoir assays maybe necessary to assess small changes in small clinical trials. Similar patterns observed with multiple types of reservoir measurements may provide more assurance of the relevance of small changes.
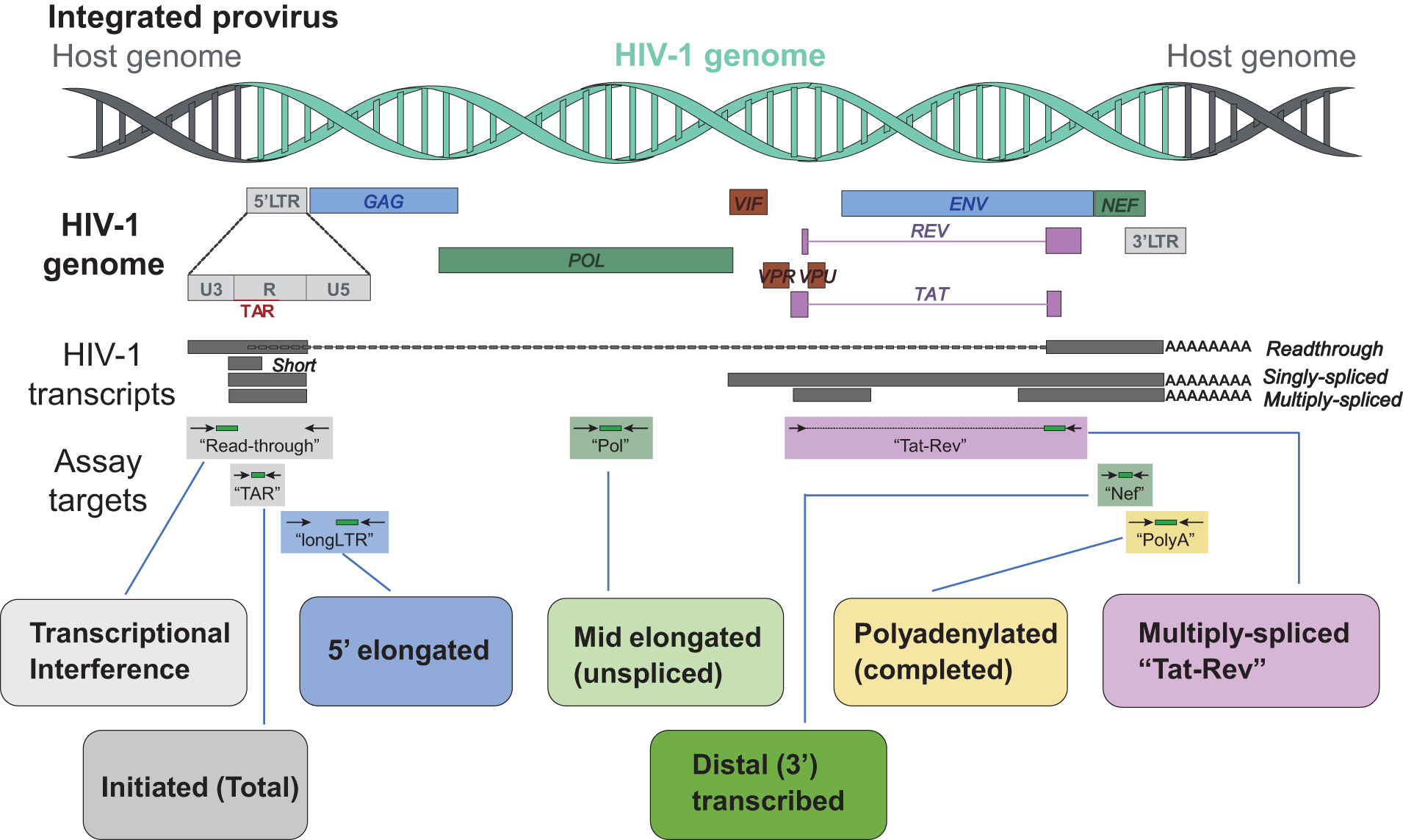
Shown is a map of the HIV genome (representing the proviral DNA), types of HIV transcripts, and the sequence regions detected by different assays. “Read-through” suggests transcriptional interference, “TAR” is found in all HIV transcripts, indicates initiation of transcription, and a more than twofold excess over long LTR suggests inhibition of elongation, “R-U5/Gag” (long LTR) represents elongation, “Pol” represents elongation past Gag and is found only in unspliced HIV RNA, “Nef” indicates that transcription has proceeded almost to the 3′ LTR, “U3-polyA” (PolyA) shows completion of transcription and is a surrogate for HIV protein, and “multiple spliced Tat/Rev” (Tat-Rev) shows completion of splicing with the potential to overcome blocks to initiation, elongation, and export, which is a surrogate for productive infection.
In the blood, most HIV-infected CD4+ cells initiated transcription. The relative order of cell frequencies positive for each HIV transcript mirrored the HIV RNA levels per 106 cells, suggesting that the observed transcriptional blocks operate in most infected cells. In tissues, there is a lower level of HIV transcription initiation than in blood. 57,58
These assays were also applied to study the effects of latency modulating agents and the mechanisms of post-treatment control. Romidepsin increased HIV transcriptional initiation and elongation in vivo, whereas ABX464 decreased HIV transcriptional initiation in vivo (latency-promoting agent). In noncontrollers, viral rebound after ATI was associated with increases in HIV transcriptional completion and splicing, not initiation. Post-treatment control was associated with an early limitation of HIV RNA splicing and a delayed reduction in completed HIV transcripts. A new assay was also developed to quantify “intact” and defective HIV RNA. The 3′ defective HIV RNA (median 86 copies/106 cells, or 94% of HIV RNA) is much more abundant than 5′ defective (2.1 copies; 5.6%) or intact HIV RNA (0.6 copies; <1%). Intact HIV RNA is transcribed by ∼3 in 107 CD4+ T cells, 0.018% of all proviruses, and 2.2% of intact proviruses.
Levels of different HIV transcripts also have implications for understanding clinical disease. First, MS tat-rev RNA is a marker for productive infection. Second, a higher ratio of US/MS HIV RNA before ART correlates with disease progression; a lower ratio is observed in long-term nonprogressors, and a higher ratio of US/MS HIV RNA at 12 weeks post-ART correlates with lower immunologic response and higher T cell activation at 48–96 weeks. Third, US HIV RNA correlates with T cell activation (%CD38+ HLA DR+) and
RAVEN Evaluation Panel Study Assays
HMMC, HIV Molecular Monitoring Core; iSCA, improved single-copy assay; PBMC, peripheral blood mononuclear cells.
Assays used to characterize all RAVEN collections were the 45-replicate Aptima assay for HIV-1 RNA in plasma (VRI), the quantitative RT-PCR (qRT-PCR) for cell-associated (CA) HIV RNA (LTR region) in CD4 cells (VRI), the nested qPCR for CA HIV total DNA (LTR-gag region) in CD4 cells (VRI), the nested qPCR for CA HIV-integrated DNA (Alu LTR sequences) in CD4 cells (VRI), and the dQVOA for replication-competent HIV in resting CD4+ T cells enriched from PBMCs. He highlighted two publications that resulted from the use of the RAVEN panel intra-lab and inter-lab comparison of QVOA, TILDA, and iCARED (inducible cell–associated RNA expression in dilution) assay. 59,60 The RAVEN panel was also involved in standardizing single-copy assays for HIV RNA in plasma. 61,62 At present, RAVEN is focused on completion of analyses and articles based on plasma and PBMC evaluation panels (Table 5).
The limit of detection of five participating ultrasensitive plasma RNA assays has been established using the RAVEN panel. In the meantime, scientific and administrative oversight of the RAVEN program including the repository and additional pedigreed panels have been transferred to the Virology Quality Assessment (VQA) Program, a National Institute of Allergy and Infectious Diseases (NIAID) contract currently held by Duke University.
IMPAACT Network HIV Remission/Cure Clinical Trials (June 2022)
IMPAACT, International Maternal Pediatric AIDS Clinical Trials.
Blinded validation panels were received from the Duke University EQAPOL program, NIAID VQA, and NIBSC, United Kingdom (subtypes A, AE, AG, B, C). Next, Dr. Persaud discussed her clinical trial experience with HIV persistence assays. In IMPAACT P1030, the first trial of lopinavir-ritonavir–based ART for early treatment of infants, the HIV DNA:QVOA ratio ranged from 66 to 148. Of note, 2-LTR circles contributed substantially to total HIV DNA during the first 2 years of early ART in perinatal infection, 65 with implications for newer assays to quantify intact proviruses, such as the FLIP-seq and IPDA discussed previously. In early-treated infants (P1030), there is a stable persistence of unspliced HIV RNA and a decrease in multiple-spliced HIV RNA. Compared with the QVOA, the TILDA assay provides a feasible approach to quantify induced proviral reservoirs in adult and perinatal infection, as fewer cells are needed.
For pediatric studies, a modified TILDA is required (the Enhanced TILDA) to maximally detect HIV reactivation. 66 For IMPAACT clinical trials applications, an IPDA that covers subtypes A1, B, C, D, and F1 is necessary, and for some studies will require CLIA certification, as for IMPAACT P1115. In summary, given the current lack of availability of a validated cross-subtype IPDA, total HIV DNA serves as a reasonable biomarker to assess the efficacy of very early treatment interventions where infected cell frequencies can reach to below the limits of detection of the assay, as well as in transplant settings aimed at treatment for malignancies where HIV eradication has occurred as in the IMPAACT P1107 study.
The addition of the Enhanced TILDA 66 to HIV DNA PCR allows the estimation of induced proviruses and their targeting, which will be greatly enhanced with a cross-subtype IPDA. The incorporation of proviral landscape analyses enables in-depth assessment of proviral dynamics, reservoir composition, diversity/clonality during novel interventions, including immunotherapies. Finally, subtype diversity and rigorous assay validation are key considerations for molecular assay applications in pediatric cure trials.
These findings are consistent with those in Simonetti et al. 22 Using the IPDA, intact and defective proviruses have been detected throughout the body (blood, intestines, liver, spleen, lymph nodes, kidney, prostate, lung, brain, and GALT) in ART-treated PLWH. In new longitudinal studies of HIV reservoir dynamics, IPDA and QVOA measures of the HIV reservoir correlate over a time span of 20 years of ART.
To support growing IPDA demand, Accelevir implemented automation across all sample processing, quality control, and IPDA execution can be further scaled. Analytical performance studies show that the IPDA is accurate and precise across multi-log range using both contrived control samples (JLAT6.3 cells, HIV spiked into negative samples) and real-world samples from PLWH. Such analytical performance data support assay use in early trials and are generally required for primary endpoints and clinical tests (Laboratory developed test and In Vitro Diagnostics). Accelevir is working toward a single, unified group M IPDA design. The IPDA expansion to new subtypes must be supported by both sequence conservation data and robust proviral SGS-based landscape analysis of intact and defective proviruses collected from cohorts with subtype and geographical diversity.
Initial Accelevir data show small but meaningful differences in the ratio between intact and defective proviruses for subtypes B, C, D, and AE, highlighting the importance of sequencing-based proviral landscape analysis in supporting assay design. To ensure that this sequencing-based proviral landscape analysis is accurate, optimal DNA extraction, primer design, and long-distance PCR chemistry are critical. In particular, DNA shearing and long-range PCR efficiency disproportionally impact intact proviruses, resulting in significant skewing of the proviral landscape. 8 In addition to the IPDA, Accelevir offers several other complementary assays for persistent proviral DNA (proviral SGS, env sequencing), virus expression (CA-RNA-dPCR, Aptima 9-rep SCA, standard HIV plasma viral load), and virus induction (QVOA).
Discussion
Two discussion panels were held, one for the preclinical session and one for the clinical session. In the preclinical panel discussion, the following questions were asked: What are the strengths of current assays? How should assays be validated? What are the limitations of current assays? What improvements are needed? Is it important to measure the latent versus the expressed reservoir? Is it important to measure the cell types comprising the reservoir? Do assays performed on blood represent tissue reservoirs?
The panelists Drs. Joel Blankson, John Coffin, Lisa Frenkel, Ya-Chi Ho, Jeffrey Lifson, Frank Maldarelli, and Janet Siliciano made the following points:
There is a strong need for modeling of virus decay before/after ATI.
Precision, sensitivity, and specificity of the assays need to be optimized, but one may then find a small difference in an intervention that is meaningless. Specificity is needed to avoid false positives, sensitivity depends on the question asked, and reproducibility is inversely correlated with the frequency of the target.
Enormous resources are invested into clinical trials (financial, investigator time, patients). The results obtained from clinical samples should be maximized and as many HIV reservoir assays as possible should be performed. For example, DNA and RNA can be extracted from the same cell. The limitation of sample numbers and volumes are the crucial issues.
Data are needed whether the viruses found in these assays are the same found in the rebound virus pool after an ATI. Rebound most likely will come from tissues not blood. The rebound virus could not be reliably detected using DNA methods. An assay for persistent virus in patients that have viremia is also needed. Stopping therapy does not induce the virus, it only reveals the virus that is already present. An ATI should not be performed on patients that still have persistent viremia, which usually consists of infectious virus. The pool of virus producing cells is very small.
Some of the rebound viruses are neutralized by autologous antibody. Pollack et al. have demonstrated that defective proviruses, particularly those with packaging signal defects, can produce viral RNA, viral protein, and virion and serve as a decoy that distracts immune responses against intact proviruses. 67 Simonetti et al. 20 have shown that residual viremia is dominated by defective clones. Measuring residual viremia is critical as a marker but taking aNAbs into consideration is also important.
NHP models and barcoded SIVs and SHIVs have many advantages, for example, for tracking clonal expansion, decay over time, but one cannot follow infected animals on ART for 20 years. Clinicians and animal modelers should work together in an iterative process.
Advancements in single-cell multi-omics enables profiling T cell clone dynamics and HIV reservoir profiling. 67,68 The field should continue to explore new assays to inform mechanisms, so the interventions can be improved.
In the clinical panel discussion, the following questions were asked:
Which reservoir assays are most important to include in interventional studies?
What assay validation is needed before incorporating into clinical studies?
What new assays are needed?
The panelists Drs. Katherine Bar, Marina Caskey, Bonnie Howell, Dan Kuritzkes, John W. Mellors, and Sandhya Vasan made the following points:
For clinical trials, assays are needed that can be used very soon; IPDA is gaining interest because it requires fewer cells. Feasibility is challenging; it needs to be determined what assays can be done with small plasma or cell amounts. The sample volume collected at key time points is critical.
Whether the latent or expressed reservoir should be studied depends on the biological question and the interventions under investigation. The target, pharmacodynamic and proof of mechanism biomarkers should be assessed. The clinical significance of protein expression is not yet understood. QVOA should be performed on all cells, not just on resting cells.
Choice in assay type would also depend on whether the goal is remission or eradication. The expressed HIV reservoir should be measured before an ATI. The assays can be used for predictive algorithms and to study the scientific mechanisms. PET imaging may be useful in assessing changes in reservoir during ATIs and effects of immune response or interventions. Other biomarkers needed are predictors of post-treatment control and predictors of viral suppression off therapy.
The limitations of the assays need to be understood. There are two main issues with the IPDA. First, there is no absolute guarantee that the provirus is intact, and second, HIV sequence polymorphism can abrogate primer binding.
The tiers of assay validation are as follows: (1) validation of the actual performance of an assay, ideally in more than one laboratory, (2) validation in the regulatory sense to show that the assay is a true correlate or surrogate.
Studying HIV clones provides a lot of information; however, rebound can come from minor variants. We do not have knowledge about variation across clones. In clinical studies, at least 1,000-fold reduction of the HIV reservoir is needed. Will the same assays work for clonal populations versus single infected cells?
The role of placebo arms in clinical trials was questioned—it is needed in the absence of validated assays and correlates or predictors of time to viral rebound. The required validation depends on the level of endpoint (primary, secondary, exploratory, etc.). At a minimum, the primary endpoint should be validated.
In-home testing assays to detect early viral rebound are also needed.
Footnotes
Acknowledgments
The authors thank Drs. Katherine Bar, Joel Blankson, Marina Caskey, John Coffin, Lisa Frenkel, Ya-Chi Ho, Bonnie Howell, Dan Kuritzkes, Jeffrey Lifson, Frank Maldarelli, Janet Siliciano, and Sandhya Vasan for serving on the discussion panel.
The authors also thank Drs. Sandra Bridges and Diana Finzi for their support, Dr. Carl Dieffenbach for the workshop opening remarks and for helpful discussions, Dr. Daniel Kuritzkes for the workshop summary, and Cynthia Soriano for her administrative support of the workshop.
In addition, the authors thank NIAID Meet, in particular Reigna Jones, Eric Delaney, and Brian Pinson, for help with execution of the workshop.
Author Disclosure Statement
Dr. R. Siliciano is an inventor on a patent application for the IPDA filed by Johns Hopkins University and licensed by Accelevir Diagnostics. He holds no equity interest in Accelevir Diagnostics.
Funding Information
We did not receive funding for the workshop since it was held virtually.