
Abstract
Adult T cell leukemia/lymphoma is a malignancy with a poor prognosis caused by human T lymphocyte virus type 1 (HTLV-1) infection. Tax and HBZ are two major viral proteins that may be involved in oncogenesis by disrupting apoptosis. Because Bcl-xL plays an integral role in the anti-apoptotic pathway, this study examines the interaction between host apoptosis and oncoproteins. We investigated 37 HTLV-1-infected individuals, including 18 asymptomatic and 19 adult T cell leukemia/lymphoma (ATLL) subjects. mRNA was extracted and converted to cDNA from peripheral blood mononuclear cells, and then gene expression was determined using TaqMan q-PCR. Moreover, the HTLV-1 proviral load (PVL) was also measured using a commercial absolute quantification kit (Novin Gene, Iran). Data analysis revealed that the mean of TAX, HBZ, and PVL was significantly higher among the study groups (ATLL and carrier groups p = .003, p = .000, and p = .002 respectively). There was no statistical difference in Bcl-xL gene expression between the study groups (p = .323). It is proposed that this anti-apoptotic pathway may not be directly involved in the development of ATLL lymphoma. Bcl-xL, TAX, HBZ gene expression, and PVL can be utilized as prognostic markers.
Introduction
Adult T cell leukemia/lymphoma (ATLL) has an extremely poor prognosis due to the progressive proliferation of infected clonal T cells. Hepatosplenomegaly, lymphadenopathy, cutaneous lesions, and a high level of lactate dehydrogenase and calcium in the blood are the most common acute conditions associated with ATLL acute conditions. 1 –3
Tumor cell migration causes damage to various organs, including the lungs, skin, spleen, liver, and lymphoid glands. In addition, patients with acute lymphomatosis are immunocompromised, resulting in opportunistic infection and hypercalcemia, the leading cause of death during treatment. 1 Although the fundamental role of human T lymphocyte virus type 1 (HTLV-1) in ATLL pathogenesis is not fully explained logically, the fundamental role of HTLV-1 in ATLL pathogenesis is aptly demonstrated. 4,5
HTLV-1 is a retrovirus that is the primary cause of two distinct diseases: ATLL and inflammatory disorders such as HTLV-1 associated myelopathy (HAM), which occurred in only a minimal number of infected individuals. 1,6 Around 10–20 million people worldwide are infected with HTLV-1. To the northeast of Iran, particularly Mashhad and Neyshabour are the two most endemic areas in the Middle East. 7
Tax and HBZ are the primary virulence factors of HTLV-1, and they are encoded by the pX region of the HTLV-1 genome. These proteins may be produced by oncogenic transformation via transactivation, deregulation of the apoptosis pathway, and DNA repair disruption in people living with HTLV-1 (PLWHTLV-1). 8,9 Due to the proper immunogenicity of Tax for immune responses, a transcript from the minus strand 3′-LTR HTLV-1 provirus expressed HBZ, a possible immune system escape mechanism. 10 –13 When each cell is integrated, the viral genome supports a high level of virus oncoprotein expression, then an aggressive, uncontrolled proliferation of the virus and infected T cells occur. 14
Apoptosis, also known as programmed cell death, is a series of intracellular proceedings among cell changes that include cell shrinkage, nuclear fragmentation, chromatin compression, chromosomal DNA fragmentation, and membrane blob representations. 15 Tax downregulate pro-apoptotic elements such as BIK, EGL-1, BIM, BMF, NOXA, BID, BAD, BNIP3, and Beclin-1. Moreover, increasing anti-apoptotic proteins such as BCL-2, Bcl-xL, BCL-w, and MCL-1 via upregulation of transcription factors such as NF-κB molecules within the cells may disrupt the cell life cycle and preserve malignant potential. 16 –18
Following caspase-8 activation by external signals such as Fas-Fas Ligand and TNF-TNF receptors, BID could be truncated to tBID, which continues to increase mitochondrial membrane permeability. As a result, releasing cytochrome C into the cytoplasm activates the effectors' caspases, such as caspase-3. 19,20 In addition, Bcl-xL inhibits this process by preventing the attachment of pro-apoptotic proteins such as BIM and BAD from attaching.
Thus, it may inhibit apoptosis initiation within the cell and prolong its life span, resulting in uncontrolled cell proliferation. Several studies demonstrate that the alteration of apoptotic mediators by HTLV viral proteins, particularly TAX and HBZ, alters apoptotic mediators and that the interaction of Tax and BH3-only proteins such as BIM and BID will promote malignant cells survival. 11
The present study examines the expression of TAX, HBZ, and Bcl-xL genes in association with HTLV-1 proviral load (PVL) in two groups of HTLV-1 infected individuals, ATLL and healthy carriers, to determine the diffractions, interactions, and correlations between host cell anti-apoptotic elements and HTLV-1 virulence properties across these populations.
Materials and Methods
Study population
The study population included 19 new cases of ATLL patients, and 18 PLWHTLV-1 referred to the hematology-oncology department of the Ghaem and Imam Reza hospitals in Mashhad-Iran between January 2018 and December 2020. The EDTA whole-blood samples were collected before any treatment proceedings and chemotherapy.
Peripheral blood mononuclear cells and RNA extraction
The peripheral blood mononuclear cells (PBMCs) were isolated using the Ficoll gradient method from the whole blood of the patients and healthy carriers (Cat No. 002 041 600, Lymphodex; INNO-TRAIN, company). According to the manufacturer's instructors, total RNA was purified and extracted from PBMCs using Trizol reagents (TriPure Cat. No. 11667157001; Roche-Germany).
All purified RNA samples were treated with DNase before converting to cDNA to prevent genomic DNA contamination. The integrity of the RNA was determined spectrophotometrically and electrophoretically on 2% agarose gels.
cDNA synthesis
Complementary DNA was generated by random primers and reverse transcriptase (Cat No.: K-2046; Bioneer). A total of 1 to 1.5 μg of RNA was reverse transcripted using the cDNA synthesis kit (Cat No.: K-2046; Bioneer). The manufacturer's instructions specified 30 s at 24°C, 4 min at 44°C, and 30 s at 55°C for the reaction.
These conditions were repeated 12 times, and the reaction was terminated at 95°C after 5 min. GAPDH primers were used to ensure proper synthesis. The following specific primers were used: GAPDH Forward Primer 5′-CAAGGTCATCCATGACAACTTTG-3′ and Reverse GAPDH Primer 5′-GTCCACCACCCTGTTGCTGTAG-3′.
Primer designing
After identifying the desired gene sequences (TAX, HBZ, and Bcl-xL) on the NCBI (Gene bank), primers and probes were designed by Allele ID (version 5) software, which was employed to design primers and probes.
Real-time PCR
Real-time PCR (TaqMan method) was performed using the AccuPower® Plus DualStar™Master Mix 2X (Cat Num: K-6603), according to the manufacturer's instructions with the primer sets. Each sample was duplicated and correlated with the expression of a housekeeping gene (β2 microglobulin). The primers and probes used to amplify the following sequences are listed in Table 1.
TAX, HBZ, Bcl-xL, and β2 Microglobulin Genes Primers and Probes
The PCR was initiated by heating the templates at 95°C for 4 min, followed by 45 cycles of denaturation for 15 s at 94°C and annealing for 20 s at the optimal T m and extension phase for 20 s at 72°C.
PVL measurement
As previously described, PBMCs were isolated from EDTA-treated blood samples to determine the HTLV-1 PVL. The rotorgen Q Real-Time PCR machine performed Real-time PCR using a commercial absolute quantification kit (Novin Gene, Iran), to measure the PVL of HTLV-1 using specific primers and a fluorogenic probe (Qiagen, Germany).
The copy number of HTLV-1 was quantified as an actual amount of cellular DNA using the albumin gene as a reference gene. The HTLV-1 and albumin DNA concentrations were determined using two 5-point standard curves. The HTLV-1 PVL was normalized using the value calculated as the ratio of (HTLV-1 DNA copies number/albumin DNA copies number/2) × 104 and expressed as the number of HTLV-1 proviruses per 104 PBMCs.
Statistical analysis
The experimental results were presented statistically as the mean ± SD. SPSS (version 18) was used for data analysis. As a result of the Kolmogorov–Smirnov analysis, statistical methods such as Mann–Whitney were utilized to calculate the p-values, and spearman analysis was used to determine the correlation between the parameters in the ATLL group and the healthy control group. p-Values <.05 were considered statistically significant.
IRB approval statement
The Ethical Research Committee of Mashhad University of Medical Sciences approved this study (A-910679). No further sampling was done for the patients, and the study investigations were done on routine samples at the commencement of admission after the full consent of patients. All costs of their examination were free for the included patients. All patients' results were kept confidential and delivered to their physicians.
Results
Demographic data and clinical status
The current study examined 19 patients (13 females and 6 males) with ATLL and 18 healthy carriers (12 females and 6 males). The mean age of ATLL and healthy carriers was 53 and 49 years, respectively.
Lymphoid gland involvement was observed in 17 patients; 5 patients had skin lesions, and 6 had immunodeficiency conditions. Two patients simultaneously had lymphadenopathy and skin lesion, whereas only one patient had both an immunodeficiency condition and skin lesions. None of the patients in this study exhibited all three clinical symptoms concurrently (Fig. 1).

Clinical manifestations of patients demonstrated a high degree of lymphoid organ involvement.
Gene expression study
TAX gene expression
This study used the expression of a housekeeping gene (β2 microglobulin) to calculate correlations and normalized target gene expression. The levels of TAX mRNA expression indicate significant differences between the ATLL and healthy carriers (Fig. 2). The mean value of TAX/β2 microglobulin was 0.1993 in the ATLL group and 0.000764 in the control groups. Mann–Whitney analysis revealed a statistically significant difference in TAX gene expression across the study population (p = .002).
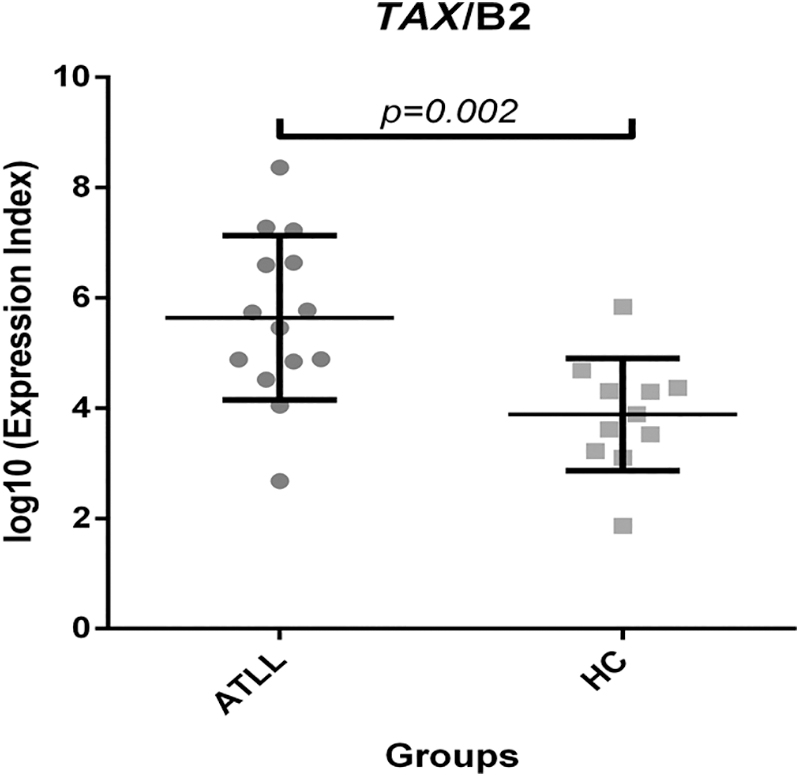
Increased TAX gene expression among ATLL patients in comparison to healthy carriers. ATLL, adult
HBZ gene expression
The levels of HBZ mRNA expression revealed significant differences between ATLL and healthy carriers (Fig. 3). The mean of HBZ/β2 microglobulin was 0.1017 and 0.00012 in the ATLL and control groups, respectively, indicating that the HBZ gene was expressed at a higher level in the ATLL patients' group. Further, this value demonstrated a statically significant difference in HBZ gene expression among the study population (p = .0001).

Increased HBZ gene expression among ATLL patients in comparison to healthy carriers.
Bcl-xL gene expression
Bcl-xL mRNA expression levels indicated no statically significant differences between ATLL and healthy carriers (Fig. 4). The mean of Bcl-xL/β2 microglobulin was 3.01 and 0.37 in the ATLL and control groups, respectively, implying that the ATLL patient group had a higher gene expression level. This value, however, does not establish a statically significant difference (p = .323).
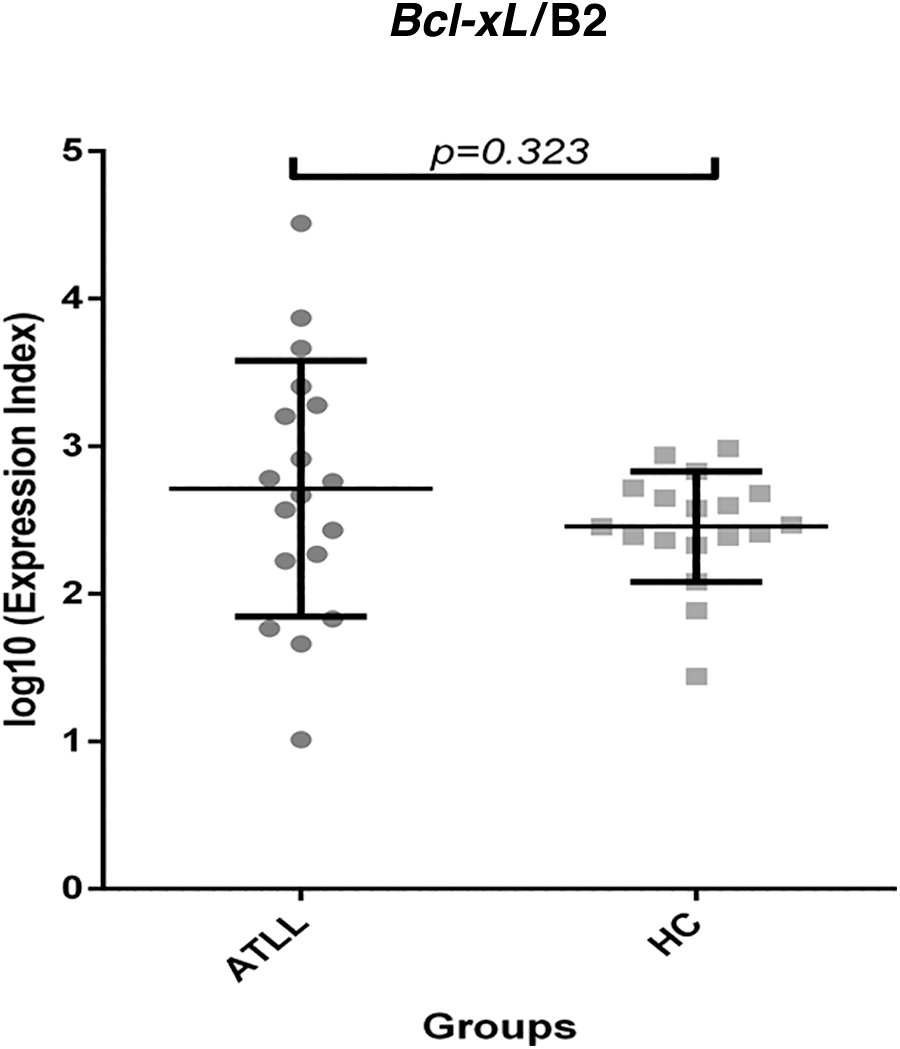
Bcl-xL gene expression level in ATLL versus HC patients. A statistical difference was not observed within the study groups.
PVL level
The mean PVL of HTLV-1 was 11431.7 ± 3774.302 copies/104 PBMCs in ATLL patients (95% CI: 53–58,983). The ATLL group had a percentage of PBMCs infected with HTLV-1 of 114.3% ± 37.7% (95% CI: 0.53–590). This finding indicates that some infected T cells exhibited multiple copies of the HTLV-1 genome.
The average HTLV-1 PVL in Healthy Carriers was 531.25 ± 446.141 (95% CI: 0–1,329) copies/104 PBMCs, with 5.3125 ± 4.46141 (95% CI: 0–13.29) percent of PBMCs being infected (Fig. 5). These values indicated a significant difference in PVL among the study population through the Mann–Whitney analysis (p = .0001).
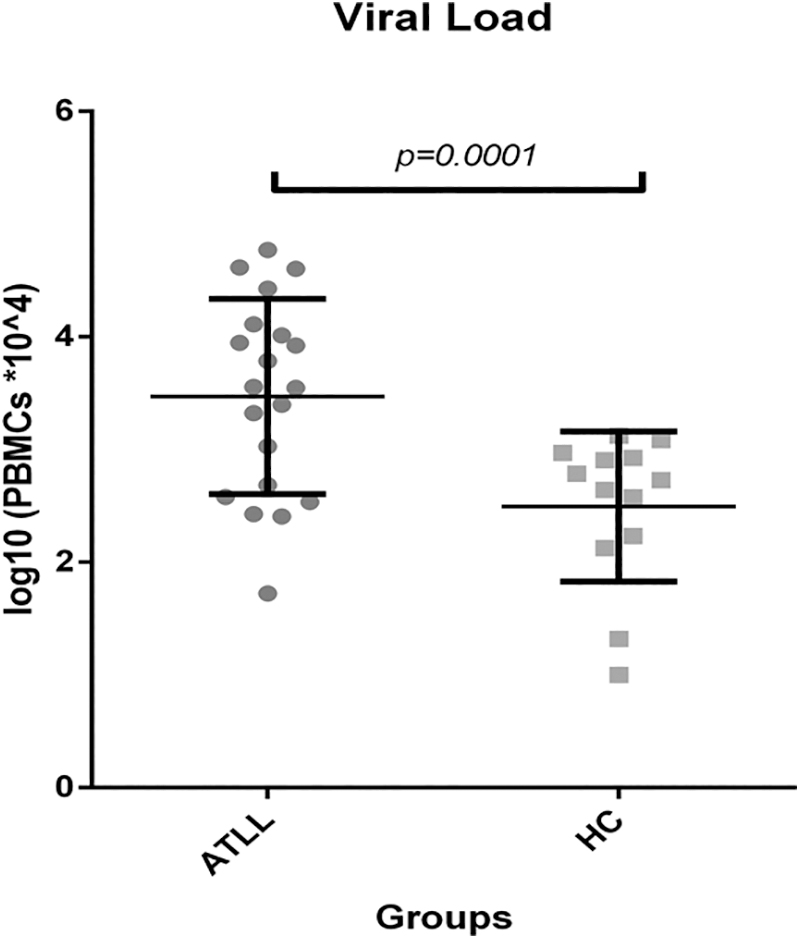
Increased HTLV-1 proviral load among ATLL patients in comparison to healthy carriers.
Correlation of gene expression and clinical symptoms
The results of the Spearman analysis revealed a significant negative correlation between lymphadenopathy and skin lesions (p = .000 correlation coefficient = −0.721). Other clinical manifestations had no significant correlation with themselves. Further, Bcl-xL gene expression was negatively correlated with lymphoid organ trigger (p = .033 correlation coefficient = −0.503).
TAX and HBZ exhibited a strong correlation with each other's gene expression (p = .001 correlation coefficient = 0.636), but only HBZ correlated with HTLV-1 PVL (p = .012 correlation coefficient = 0.469). No correlation between viral gene expression and clinical manifestations was observed. The mean age of patients was significantly correlated with TAX, HBZ, PVL, and immune deficiency (p = .001, .002, .005, and .043 respectively; correlation coefficient = 0.611, 0.537, 0.470 and 0.456). Moreover, no correlations were observed between gender, clinical manifestations, and viral gene expression (Table 2).
Spearman's Rho Correlation Between Clinical Manifestations and Gene Expression
The bold items are statistically significant.
p-Value: two-tailed.
Cor, correlation; ID, immunodeficiency; LA, lymphadenopathy; PVL, proviral load; SL, skin lesion.
Discussion
HTLV-1 is an onco-lentivirus that is responsible for most ATLL, and HAM patients. 21 ATLL is a progressive malignancy, and the majority of diagnostic patient cases are lymphomatous or acute stages. Lymphadenopathy, hepatosplenomegaly, skin lesions, hypercalcemia, and flower-like T cells circulating in the blood are the primary indicators of a poor prognosis and short life span in infected individuals. 1
Our findings directly correlate with increasing age and viral elements progression (Table 2). In addition, a critical health condition such as opportunistic infection demonstrates this correlation. This study discovered that Tax and HBZ had significantly higher PVLs and gene expression (Figs. 3 and 4).
Viral pathogenic elements such as Tax and HBZ may play an essential role in transforming infected T cells into malignant lymphocytes, as HBZ promotes uncontrolled cell proliferation and appears to be required for ATLL. 9,22,23 Although Tax promotes the propagation of infected T cells, its significant molecular weight and complex protein structure made it a prime target for CTL immune response.
Therefore, in ATLL patients, HTLV-1 inhibited TAX gene expression and activated HBZ transcription to evade the immune system. 24 We observed significantly higher HBZ gene expression than Tax expression in this study; further, HTLV-1 PVL correlates positively with HBZ (Table 2). Despite the downregulation of apoptosis in ATLL patients, which results in T cell proliferation, we observed no significant difference in Bcl-xL gene expression (Fig. 5).
Apoptosis can be disrupted either by decreasing pro-apoptotic elements or by increasing antiapoptotic elements. Tax and HBZ have been shown to activate the PI3K pathway, resulting in the phosphorylation of Akt. Then, disrupting the CD95 forkhead, caspase-9 and BAD can cause apoptosis. 25,26 Moreover, other studies demonstrated that TAX and HBZ induce c-Flip inhibiting apoptosis from the external trail. 12,13
Another possibility for the Bcl-xL mRNA expression pattern is that Bcl-2 family member containing Bcl-2-related protein A1 or Blf-1 is promoted, as Bfl-1 expression induced by TAX protein contributes to the survival of HTLV-1 infected T cells. 27 Nicot et al. investigated the level of Bcl-xL expression in the ATLL samples. 28
They stated that TAX could increase Bcl-xL expression in T cell infected cell lines. The study demonstrated that ATLL patients have a higher level of Bcl-xL expression than healthy carriers, but this increased level is not statically significant. This discrepancy in results could be explained by the different methods of investigation used, q-PCR versus Western blotting. Further, they demonstrated that higher levels of Bcl-xL were observed in malignant HTLV cell lines. In contrast, higher levels of Bcl-xL were observed in ATLL samples compared with non-infected subjects or healthy carriers. In the present study, the findings may imply an independent Bcl-2 or Bcl-xL anti-apoptotic pathway in T cells infected with HTLV-1.
Conclusion
Our results suggest substantial Tax, HBZ, and PVL existence in ATLL patients, which can be used for diagnosis, treatment monitoring, and prognosis. In contrast, Bcl-xL appears to not correlate with HTLV-1 pathogenesis.
Footnotes
Acknowledgments
The authors want to express their gratitude to every member of their laboratory team who assisted and completed this project.
Authors' Contributions
All authors contributed to the study conception, and this review was designed by [Z.F.], [M.M.A.], and [S.A.R.]. Data collection and analysis were performed by [M.M.A.], [S.A.R.], [S.R.], and [H.R.]. All clinical data and patient samples, including ATLL and healthy carriers, were selected by [H.R.]. The first draft of the manuscript was written by [M.M.A.], [Z.F.], and all authors commented on previous versions of the manuscript. The first draft was edited by [S.A.R.]. All authors read and approved the final manuscript.
Authorship Confirmation Statement
The experiments were carried out in Immunology Research Lab, Inflammation and Inflammatory Diseases Division, Immunology Research Center, the Mashhad University of Medical Sciences (MUMS), Mashhad, Iran.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financially supported by Mashhad University of Medical Sciences, grant number: 910679. All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript. The authors have no financial or proprietary interests in any material discussed in this article.