
Abstract
Glycoprotein 41 (gp41) of the human immunodeficiency virus type 1 (HIV-1) protein plays a critical role in membrane fusion. Gp41 binds to proteins in the plasma membrane of CD4+ T cells, particularly the T-cell antigen receptor (TCR). These findings indicate that gp41 is involved in the assembly of HIV-1 at the plasma membrane of T cells and affects the stimulation of the TCR. To control HIV-1, new inhibitors were introduced to target the gp41 protein. However, mutations in this region might reduce their efficacy. The Gp41 region was amplified from the sera of 30 patients using nested polymerase chain reaction. The sequences were analyzed by bioinformatics tools to identify mutations and gp41 structural features. Subtyping and the interaction between fusion inhibitors and gp41 proteins were also examined. As the first report from Iran, docking analysis between fusion inhibitors and Iranian gp41 proteins showed that mutations in gp41 could not reduce the efficacy of the fusion inhibitors. Most of the patients were infected with CRF35-AD. Several post-modification positions, including glycosylation and phosphorylation sites, were identified in the gp41 protein. Our findings revealed no known multinational drug resistance to gp41 inhibitors; thus, fusion inhibitors can effectively inhibit HIV in Iranian patients. In addition, the present study introduced a new gp41 region (36–44 aa), which considerably influences the interactions between gp41 inhibitors and the gp41 protein. This region may play a pivotal role in suppressing gp41 inhibitors in CFR35-AD. Furthermore, gp41 can be considered a good target for subtyping analysis via the phylogenetic method.
Introduction
Infectious diseases inflict a significant burden of morbidity and mortality worldwide, particularly in low- and middle-income countries. 1 –5 One of the most significant concerns in public health worldwide in recent decades has been human immunodeficiency virus type 1 (HIV-1), the virus that causes acquired immunodeficiency syndrome (AIDS). 6,7 The spread of HIV-1 in the human population is influenced by a combination of complex genetic, social, and epidemiological factors, making the management and prevention of the disease more challenging. 8 –12 The HIV-1 genome is a coding RNA encompassing 9 open reading frames (ORFs) encoding 15 distinct proteins. Glycoprotein 120 (gp120) is a subunit on the surface, whereas gp41 is a subunit within the membrane. These proteins are derived from glycoprotein 160 (gp160), which generates a polyprotein divided into two subunits, gp120 and gp41, by a cellular protease. 13 The proteins gp120 and gp41 play a role in membrane fusion, which is the main step in the process of infecting susceptible cells with HIV. 14 Antiretroviral therapy (ART) has revolutionized the fight against the HIV pandemic and has significantly suppressed HIV replication; however, ART strategies require lifelong treatment, as ART interruption leads to rapid viral rebound. 15,16 One of the problems in the suppression of this virus is the presence of HIV reservoirs, which are persistent cellular reservoirs of latent HIV that contribute to viral rebound when ART is interrupted. 17 These reservoirs can be found in myeloid cells such as monocytes and macrophages in the blood and tissues of people with suppressed HIV. 18 Latent HIV can also be detected in CD4+ T cells, which are latently infected cells that can be found in various tissues. 19 The development of methods to detect and remove these latent reservoirs is crucial for achieving a cure for HIV. 20 Moreover, understanding the mechanisms driving HIV latency and latency reversal is important for developing strategies to target and eliminate these reservoirs. 21 Among different fusion inhibitors, Enfuvirtide is the first approved gp41 fusion inhibitor with low toxicity. This fusion inhibitor is a 36-amino acid peptide 22 used for HIV-1 fusion inhibition. It has been shown to have a high resistance barrier and can effectively inhibit the entry and fusion ability of viruses. 23 Resistance mutations to Enfuvirtide have been mapped throughout the envelope protein of HIV-1, including the N-terminal heptad repeat (NHR) and other regions involved in fusion intermediates. 24 Enfuvirtide works by binding to a specific site within the gp41 heptad repeat 2 (HR2) domains, specifically at residues 643–678. This action disrupts the triple structure of gp41 by preventing the HR1 domains from pairing, which, in turn, inhibits the attachment between HR1 and HR2. 25
The C34 peptide, considered an active inhibitor of gp41, contains a pocket-binding area within the C34 peptide that plays a crucial role in preventing the formation of the six-helix bundle (6-HB) found in viral gp41. This region also includes the Nervy homology region 1 (NHR1) domain, which is important for its anti-HIV-1 effectiveness. 26 Several studies have reported HIV resistance to fusion inhibitors and virological response failure. 27,28 VIR-576, an experimental drug, was introduced in 2007 and is still in clinical trials. 29 This synthetic peptide can bind to the HIV1-gp41 protein and prevent the virus from inserting itself into a host cell. Therefore, VIR-576 inhibitors can potentially inhibit HIV infections in their early stages. 30
During the past few decades, new approaches in the field of infectious disease detection, diagnosis, and treatment have been explored in recent research. 31 –33 Bioinformatics has provided various tools for analyzing infectious genomes by predicting the structure and function of proteins and designing a new generation of vaccines. 34 –37 For example, docking analysis may provide findings that are helpful in optimizing and developing HIV-1 inhibitor peptides. The present study aimed to examine drug resistance mutations in the gp41 gene to examine the possibility of using fusion inhibitors in Iranian patients. This is the first study from Iran to investigate significant mutations in the HIV1-gp41 gene among Iranian-infected patients by describing the major physicochemical features of the gp41 protein using bioinformatics tools and analyzing the interaction of three gp41 inhibitors (Enfuvirtide, VIR-576, and C34) with the gp41 protein.
Materials and Methods
Study population
The sera of 30 patients were obtained from the Behavioral Diseases Consultation Center in Shiraz, which is affiliated with
RNA extraction, real-time polymerase chain reaction, nested real-time polymerase chain reaction, and CD4 count
Viral RNA was extracted from a 200 μL serum sample by the QIAamp Viral RNA Mini Kit (Qiagen, Germany) according to the manufacturer’s instructions. Extracted RNA was eluted in 50 μL of DEPC water and stored at −70°C for cDNA synthesis. Full-length cDNA was generated by reverse transcriptase (RT) using an RT kit (CinnaGen Co., Iran) by RNA denaturing at 95°C for 5 min and rapid cooling on ice. After cDNA synthesis, gp41 amplification was performed using a PCR kit (CinnaGen Co.). Each 25 μL reaction mixture contained 6.125 μL of nuclease-free water, 12.5 μL of 2× Master Mix, 0.25 μL of 50 μM forward primer, 0.75 μL of 50 μM reverse primer, 1 μL of 20× RNase Inhibitor mix, and 5 μL of cDNA. The process for nucleic acid amplification involved three thermal cycles, namely, denaturation, annealing, and extension, using the first and second rounds of thermal cycling (Applied Biosystems Thermal Cycler, California, USA) (Table 1). The PCR products were examined on a 2% agarose gel (DNA, Iran) and subsequently sequenced. An Artus HI Virus-1 real-time polymerase chain reaction (RT-PCR) kit (Qiagen) was used to determine the viral loads of the samples. To prepare the master mix, 12 μL, 18 μL, and 2 μL of HI Virus-1 RG Master A, HI Virus-1 RG Master B, and HI Virus-1 RG internal control (IC) were mixed, respectively. Then, 30 μL of the master mix was pipetted into each PCR tube, and 20 μL of the eluted sample RNA was added. Similarly, a positive control consisting of 20 μL of one of the quantitation standards (HI Virus-1 RG QS 1–4) is needed, along with a negative control consisting of 20 μL of water (PCR grade). Three steps were completed to amplify and quantify HIV-1 RNA: Hold1, Hold2, and cycling. Hold 1 at 50°C for 30 min; hold 2 at 95°C for 30 s, 50°C for 60 s, and 72°C for 30 s; and cycling for 50 cycles. Detection of a signal in the cycling green and orange channels showed amplification of the HIV gene and IC, respectively.
The Primers Used in This Study and the Thermal Cycling Conditions Used for PCR
PCR, polymerase chain reaction.
CD4 counts were measured using the BD FACSPresto™ Near-Patient CD4 Counter (B.D. Biosciences, USA), a small portable device that uses multicolor fluorescence imaging and absorbance spectrometry for the direct quantification and enumeration of CD4 absolute count and CD4 percentage in lymphocytes in a single drop of capillary or venous blood. To retrieve the results from the system, an aliquot of well-mixed ethylenediaminetetraacetic acid blood was transferred to the cartridge. The cap of the cartridge was subsequently shut, and the cartridge was placed on the workstation for an incubation period of 18–120 min at room temperature. Following the incubation, the test strip was removed, and the cartridge was placed into the analyzer where the screen displayed individual percentages and absolute CD4+ T lymphocyte counts. This information was then automatically printed within a time frame of 2–3 min.
Amino acid changing and phylogenetic tree
The CLC sequence viewer (Qiagen) 38 was used to investigate the mutations in all sequences, and all sequences were stored and aligned in comparison with reference sequences 1 and 2 (accession numbers AAF74234 and AB703607, respectively) and with 89 selected reference sequences to define the substitutions (Supplementary Table S5). The aligned sequences were used to construct phylogenetic trees via the neighbor-joining method via CLC Workbench 5.
Physicochemical properties
ExPASy’s ProtParam 39 –41 was used to estimate all properties of the gp41 protein, including the theoretical isoelectric point (pI), extinction coefficient, instability index, molecular weight, total number of positive and negative residues, aliphatic index, and grand average hydropathy (GRAVY). Supplementary Table S1 summarizes all software used in the current study.
Post-modification changes
NetPhosK [20, 21], DISPHOS, 42,43 and NetPhos 40,44 were applied to predict the serine, threonine, and tyrosine phosphorylation sites of eukaryotic proteins. NetPhosK was used to determine the kinase-specific phosphorylation sites in eukaryotic proteins. DISPHOS and NetPhos defined the serine, threonine, and tyrosine phosphorylation sites in eukaryotic proteins. N-glycosylation sites were predicted via NetNGlyc and GlycoEP.
Secondary and tertiary structures
To compute the secondary structure, SOPMA software 45 was applied for all sequences; the similarity threshold and window width were 8 and 17, respectively; and the number of conformational states, including helices, sheets, turns, and coils, was 4. To determine the tertiary structure of the selected sequences, the amino acids were submitted to the FASTA format in I-TASSER, 46,47 and the best model was selected according to the highest C-score that was refined via GalaxyRefine 43,48 and 3Drefine. 49 Each program offered five refined structures that were evaluated for stereochemistry, reliability, and quality by the ProSA-web, 50 RAMPAGE, Qmean, 51,52 and ERRAT. 53
Docking analysis
Hex software was used to identify possible interactions between the gp41 protein and three possible inhibitors (C34, VIR-756, and Enfuvirtide). In addition, the docking results were visualized by Discovery Studio to determine the amino acids involved in the binding site.
Subtyping
The subtypes of HIV-gp41 in our samples were identified by phylogenetic tree and tree online software, including Stanford HIVdb version 6.0.10, COMET version 2, and NCBI. To generate a phylogenetic tree, 89 selected reference sequences (Supplementary Table S5) were aligned with our samples using the neighbor-joining method in CLC Workbench 5.
Statistical analysis
A Kolmogorov–Smirnov normality test was performed on each quantitative dataset. As all the data did not have a normal distribution, nonparametric approaches were used. In addition, two independent t-tests were used to compare viral load and CD4 T-cell count with sex. Bivariate statistical tests, such as the Pearson correlation coefficient, were used to analyze the relationships between CD4 and viral load parameters and other quantitative data. A nonparametric Kruskal–Wallis test was used to investigate the difference between groups, the four stages of the disease, the viral load, and the CD4 count in patient samples.
Results
Demographic data
Thirty participants were enrolled in the study, and the demographic information of the study group is outlined in Table 2. According to our findings, there was a negative correlation between the amount of HIV-1 and the HIV-1 load and CD4+ T-cell counts (r = −0.42).
Pretreatment Characteristics of HIV-Infected Patients
ART, antiretroviral therapy; HIV, human immunodeficiency virus; SD, standard deviation; ALT, alanine transaminase; AST, Aspartate transferase.
Amino acid changes
Several mutations were found in the gp41 region, which are not known as resistance mutations in gp41 inhibitors (Table 3). Compared with reference sequence 1, our analysis revealed 27 substitutions. In addition, compared with reference sequence 2, it had 12 high-prevalence substitutions. The gp41 region may play a pivotal role in suppressing gp41 inhibitors in CRF35-AD.
List of Mutations in Comparison with Two Selected References
Physicochemical properties
The findings from the ProtParam software are outlined in Supplementary Table S2. Variations in sequences were identified based on characteristics such as estimated half-life and instability index.
Post-modification regions
The predicted phosphorylation sites can be found in Supplementary Table S3. Our data revealed that gp41 is heavily phosphorylated, and there are several conserved positions in the gp41 protein. Glycosylation prediction using multiple online tools identified specific positions (56, 77, 110, 51, and 77) in the majority of sequences. The findings are presented in Supplementary Table S3.
Secondary and tertiary structure prediction
According to the SOPMA analysis, the alpha helix was the main secondary structure in the gp41 protein (Table 4). I-TASSER examined the 3D structure, and the model with the highest C-score was chosen for refinement by both the Galaxy server and the 3drefine software. Various web servers, including Ramachandran, ERRAT, QMEAN, and ProSA, were used to conduct the validation analysis. The Ramachandran plot, provided by the PROCHECK server, was used to evaluate the stereochemical quality of the protein structure in different proposed models. The qualified model contains fewer residues in the Ramachandran disallowed area and more in the favored Ramachandran region. The ERRAT tool measures the quantity of nonbonded interactions between various atomic types and models, with a threshold value of 85, indicating eligibility for additional analysis. 54 QMEAN evaluates the similarity between the experimental and the proposed model structures, and a Z score of approximately 0 is considered for docking analysis. 55 Finally, the Z score generated by the ProSA-web server was used to assess the overall quality of the model, and a positive score showed the problematic parts in the input structure. 54 The most qualified models (listed in Table 5 as models G3 and G5) were used for the docking analysis. The final tertiary structures of these models are shown in Figure 1. The similarity between the two reference sequences was determined using translation memory (TM) alignment. This method revealed a significant similarity. The results of refining the reference models are displayed in Table 5, and selected models are bolded.

Final structures of two references and the alignment. (A) (AAF74234), (B) (AB703607), and (C) The alignment between two references using TM alignment.
The Secondary Structure Prediction Results for Five Selected Samples. The Majority of the Secondary Structure Was the Alpha Helix
Results of the Evaluation of the Suggested Models for Two Reference Sequences 1 and 2 [AAF74234 and AB703607], by I-TASSER, and the Best Models (G3 and G5) Were Selected. The R1-R5 Models Were the Refinement Results of the 3D Refinement Software, Whereas the G1–G5 Models Were the Refinement Results of the Galaxy Tool
The bold region represents the results obtained from the best models mentioned in the two references.
Docking results
The findings of the docking analysis presented in Table 6 indicate that the highest energy level was found in the bond between C35 and the gp41 protein, whereas the lowest energy level was observed in VIR-576. The specific locations where the selected inhibitors interact with the gp41 protein are listed in Table 6. Analysis of docking interactions between gp41 inhibitors revealed a significant energy value representing interactions between the inhibitors and gp41, which suggests that these inhibitors have the potential to inhibit the function of gp41. The gp41 amino acids located in the interaction sites were conserved among the sequences of our samples, suggesting the ability of fusion inhibitors to suppress the HIV fusion step; thus, structural alterations necessary for gp41 to finalize the fusion procedure will be hindered. Considering the interaction regions of all the inhibitors, it is suggested that the region between amino acids 55 and 78 be investigated further to monitor its effects before the administration of gp41 inhibitors as this region is highly conserved in gp41 proteins. Figures 2, 3, and 4 depict the interaction sites of three specific HIV inhibitors.
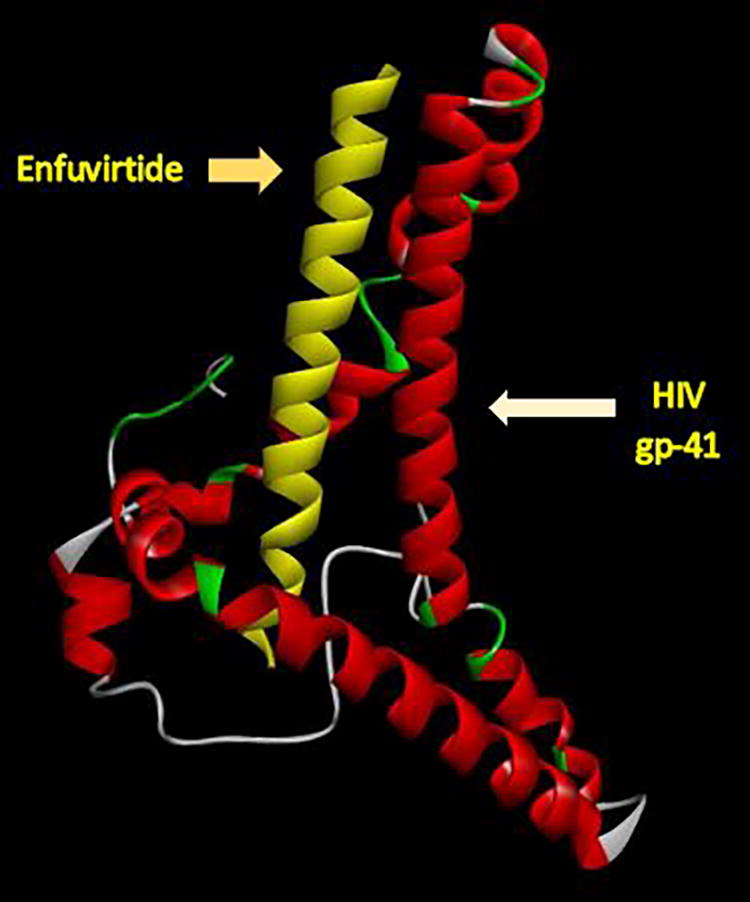
The interaction site of Enfuvirtide and viral gp41 protein docking analysis shows three amino acids in the interaction site (ASN56, LYS57, and GLU74).

The interaction site of C34 and viral gp41 protein docking analysis shows several amino acids in the interaction site.
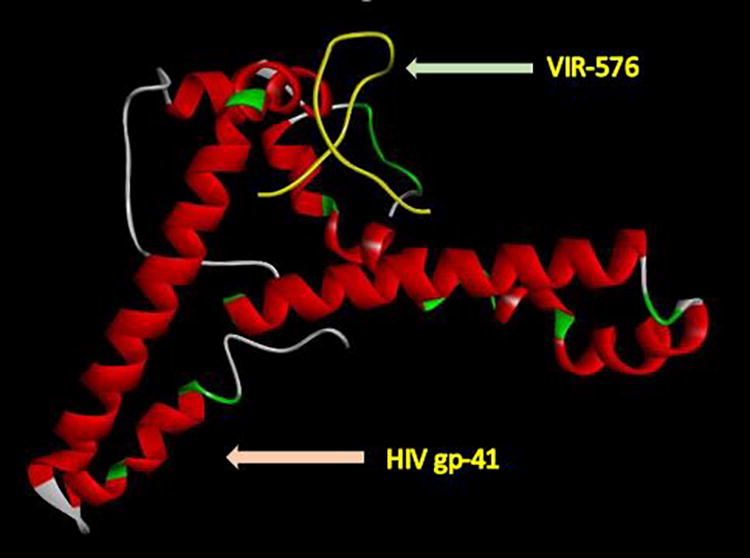
Docking analysis of the VIR-576 and viral gp41 proteins determined one amino acid in the interaction site (ASN 77).
Results of Gp41 Inhibitor Docking Analysis Using Hex and Discovery Studio Software
Subtyping results
The phylogenetic tree showed that the dominant subtype among our samples was CRF35-AD (21/30, 70%), followed by CPXs (6/30, 20%), A2 (2/30, 6.66%), and A1 (1/30, 3.33%). The CPX subtype is a recombinant form of HIV created from the combination of several viral subtypes, including A1. 56 –58 According to our data, three of our samples (13, 18, and 56) are located near the roots of CPXs (Fig. 5). According to the map of the recombination pattern of CRF35-AD, the gp41 sequence is located in the A subtype region (accession number AF095) (Fig. 6); therefore, this gp41 cannot be considered a good target for subtyping analysis via various methods. The results from three online tools are summarized in Supplementary Table S4, which shows that the majority of the sequences belonged to A1.
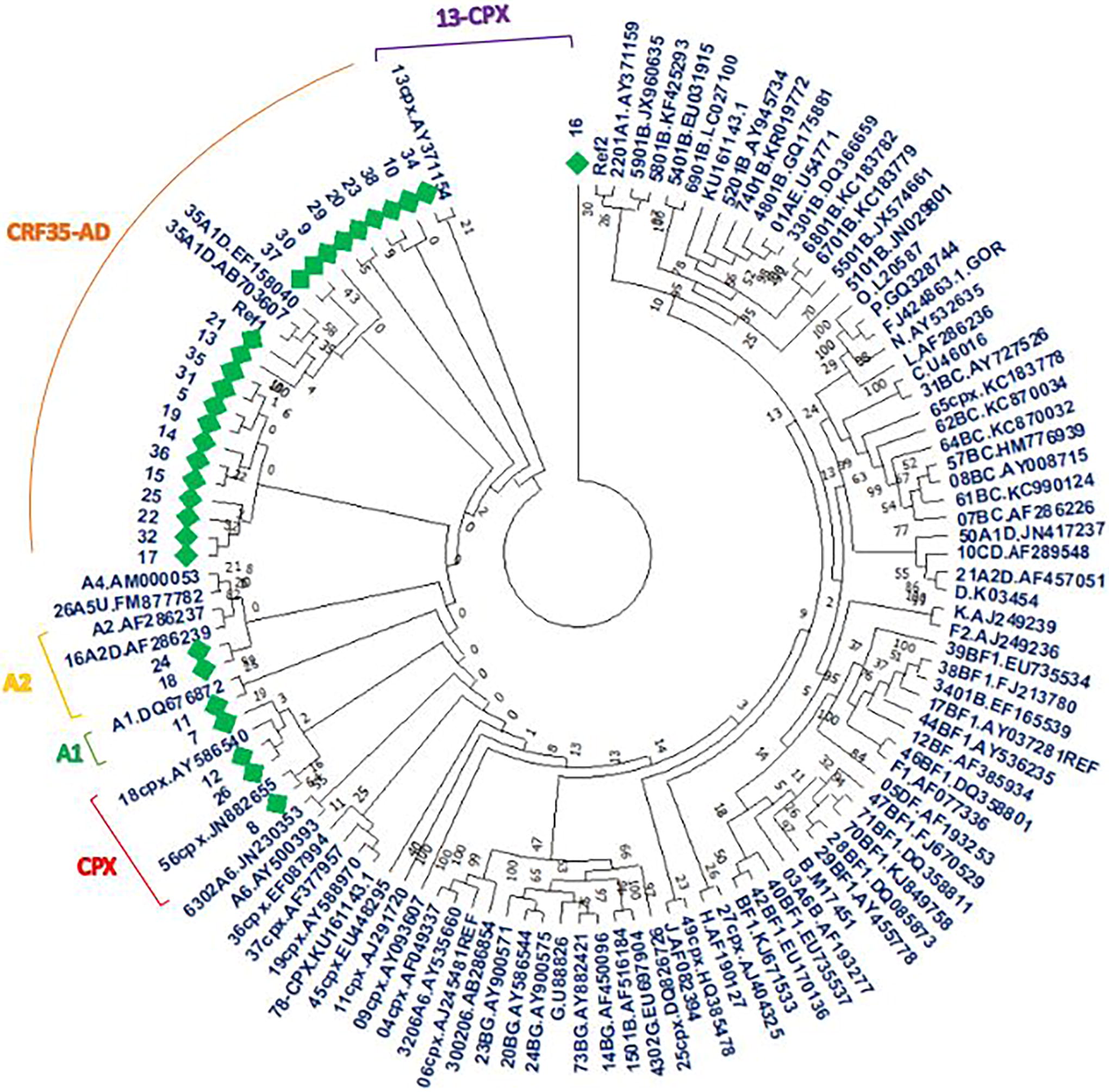
The phylogenetic tree constructed for all samples and several reference sequences, neighbor-joining method, and 1,000 bootstrap replicates. Our samples are shown with green diamonds.

The figure was adapted from https://www.hiv.lanl.gov/content/index, which shows the recombination pattern in the CRF35-AD subtype (accession number AF095). The purple and red colors indicate subtypes D and A1, respectively.
Discussion
This is the first study conducted in Iran examining mutations causing drug resistance to fusion inhibitors. Although there were several mutations in the gp41 gene, none of these mutations resulted in resistance to Enfuvirtide, VIR-576, or C34 in HIV-infected Iranian individuals.
Two specific regions within the gp41 protein, amino acids 36–44 and 55–78, may play a significant role in binding to certain inhibitors of gp41, such as Enfuvirtide, VIR-576, and C34. 59,60 Similar to our study, Joanna Smoleń-Dzirba et al. 61 did not find any mutations in codons 36–45. However, Cardoso et al. reported that G36E, N42T, and N43S induced resistance to Enfuviritide; 62 however, these mutations were not detected in our study.
Aquaro et al. identified V38A/E, which is a particular Enfuvirtide resistance mutation, in 54 Enfuvirtide-treated patients. 63 However, our study did not observe this mutation in our patients, suggesting that Enfuvirtide could effectively control HIV in Iranian patients.
Yu et al. reported that the Q64L mutation results in resistance to the C34 fusion inhibitor. 64 Our docking analysis of C34 with gp41 revealed that the Q64L mutation is located outside of the interaction site. Therefore, this mutation is unlikely to impact the effectiveness of the C34 inhibitor. Therefore, it can be inferred that a C34 inhibitor may be effective in the treatment of HIV infections in Iran.
Overall, the following drug resistance mutations in gp41 against Enfuvirtide are known: G36A/D/E/S/V, 62 V38A/E/K/M, 63 Q40H/K/P/T, 65 N42D/T, 62 N43D/H/K/S, 62,66 L44M, and L45Q/M. 67 None of the mutations discussed were found in our samples, suggesting that Enfuvirtide may be a promising treatment for Iranian HIV patients, particularly those who have not responded well to previous ART. This conclusion is supported by the docking analysis results, which demonstrated that amino acid residue substitutions did not impact the effectiveness of Enfuvirtide, VIR-576, or C34.
Despite several substitutions in our sequences, the tertiary structures of the two selected references (AB703607 and AAF74234) were similar to our results; therefore, the mutations did not have any significant effect on the structure of gp41.
In this study, the structural examination revealed that the two-dimensional structure of the gp41 protein consisted of an alpha helix and a few random coils.
Physicochemical analysis revealed that gp41 is an acidic protein and that mutations did not impact the protein’s pI or its functions, facilitating the HIV fusion process through its hairpin structure. These characteristics could be advantageous in developing HIV vaccines and medications targeting gp41.
The aliphatic indices of all the sequences suggested that gp41 is a stable protein at high temperatures. The negative GRAVY values of the selected sequences indicate that gp41 is a hydrophilic protein.
Exposing the hydrophobic structure of the gp41 fusion peptide can lead to the fusion of HIV to the host cell membrane. The estimated half-life in this study showed that the gp41 protein was stable in all possible hosts. Although these results agreed with the bioinformatic analysis results presented in this study, real-world evidence and valid experiments need to be performed to confirm whether the gp41 protein is stable in Escherichia coli.
Exposing the structure of the hydrophobic gp41 fusion peptide could result in the fusion of HIV with the host cell membrane. The estimated half-life of the gp41 protein in the present study indicated that the protein is stable in various hosts, which is consistent with the results of the bioinformatic analysis in the present study. However, additional real proof, evidence, and valid experiments are necessary to verify the protein’s stability in E. coli.
The reports of Mathew et al., 68 Zhu et al., 69 and Agrawal-Gamse et al. 70 confirmed the high expression of gp41 in yeast, and they suggested that yeast is a suitable host for post-modification processes of glycosylation and phosphorylation, which are essential for gp41 protein function.
Posttranslational modification is an essential step in the activation of viral proteins. According to our research, glycosylation and phosphorylation occur within the gp41 protein. Previous investigations have demonstrated that such modifications play a crucial role in gp41 functions, particularly in triggering signaling pathways in the host and mutations. Our results indicate that different glycosylated and phosphorylated sites were found in conserved regions of our sequences.
Previous studies have demonstrated that the ectodomain of HIV-1 gp41 includes four consensus sites at locations 100, 102, 105, 107, 114, 116, 126, and 128 that play a role in N-linked carbohydrates; however, 71 we did not find any suggested positions in our prediction. Research has shown that glycosylation reduces the area of the gp41 surface that can be targeted by the immune system, which helps HIV to spread through the human body and plays a critical role in HIV pathogenesis.
In this study, subtyping of gp41 was also conducted. The analysis of the phylogenetic tree revealed that 70% of the samples belonged to the CRF35-AD subtypes, whereas new subtypes, namely, CPXs, A2, and A1, were also detected. Nevertheless, the majority of CRF35-AD subtypes were found in distinct clusters, indicating closer evolutionary connections with sequences obtained from France, Coba, Cameron, South Korea, and Australia. There is a plausible theory that HIV outbreaks could have originated in the countries mentioned previously and spread to Iran. Supporting this theory, Eybpoosh et al. demonstrated that the CRF35-AD subtypes found in Iranian patients were closely related to the parental sequences of Kenya and Uganda, suggesting transmission to Iran through Afghan refugees. 72
Subtyping was also conducted via online subtyping tools, including COMET, Stanford, and NCBI. Subtyping revealed that subtype A was identified for all the sequences by the COMET and Stanford serves. In contrast, the NCBI genotyping tool detected novel subtypes, such as CRF09 (nine sequences), CRF03 (one sequence), CRF02 (one sequence), and CRF16 (one sequence). Similarly, several studies 73 –75 showed that subtype A was a significant subtype in Iranian infected patients, which contrasts with the findings of the present study and other studies from Iran. 76 –78
Similar to our phylogenetic results, recent reports have shown that CRF35-AD is the predominant HIV subtype in Iran 79,80 (Fig. 6), and it can be concluded that the COMET, Stanford HIVdb, and NCBI genotyping tools are not appropriate for determining HIV subtypes using the gp41 gene.
Finally, our data showed a negative correlation between the CD4+ T-cell count and plasma RNA level in an HIV-1-infected individual. Although this result is not always absolute, some patients with a high CD4+ T-cell count may suffer from a high viral load and vice versa. 81,82
The limitation of this study is that despite the various advantages of bioinformatic tools, such tools suffer from some drawbacks; for example, they cannot consider the influence of all other factors in living systems that affect the stability or the half-life of the protein.
This study suggested that the most important fusion inhibitors, C34, Enfuvirtide, and VIR-576, may efficiently suppress HIV replication in Iranian HIV-infected patients because the mutations in gp41 did not significantly interrupt the interactions between the inhibitors and gp41. In addition, the subtyping results showed that the gp41 gene is not reliable for HIV subtyping in Iranian patients.
Conclusion
Mutations associated with drug resistance against the fusion inhibitors Enfuvirtide, VIR-576, and C34 were not detected in the samples from Iranian patients, suggesting that fusion inhibitors may effectively control and suppress HIV infection in this population. Moreover, subtyping of HIV using the gp41 sequence, not genotypic tools such as COMET, the Stanford HIVdb, and the NCBI, will be accurate if a phylogenetic tree is applied. Further studies with larger sample sizes are necessary to assist Iranian policymakers in developing an improved treatment approach to control HIV in the country.
Footnotes
Acknowledgments
The authors would like to thank Shiraz University of Medical Sciences, Shiraz, Iran. The authors wish to thank Ms. Susan Craig (Atlanta, Georgia, USA) and Mr. Hosein Argasi at the Research Consultation Center (RCC) of Shiraz University of Medical Sciences for their invaluable assistance in editing this article; Dr. Mohamad Javad Shekiba for her clinical consultancy; Mr. Behzad Rezaei for his support; and Dr. Parisa Keshani for her assistance in the statistical analysis.
Authors’ Contributions
F.G.: Performed the experiments. A.H. and M.A.D.: Designed the study and was involved in the conceptualization, supervision, and resources and funding acquisition. F.G., A.H., and Z.H.: Provided scientific suggestions and controlled the experiments. B.D., F.B., and Z.H.: Collected the data. N.K., F.B., F.G., and Z.H.: Analyzed the data. F.G., Z.H., and N.K.: Statistically analyzed the data. M.A.D. and F.G.: Wrote the original draft; and A.H.: Reviewed and edited the article. All authors have read and approved the final article.
Author Confirmation
All authors are from the Shiraz University of Medical Sciences (Shiraz, Iran), where education and research are the primary functions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by Shiraz University of Medical Sciences (grant numbers 28432 and 97-01-59-18790).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.