
Abstract
Multifaceted natural killer (NK) cell activities are indispensable for controlling human immunodeficiency virus (HIV)-1 transmission and pathogenesis. Among the diverse functions of NK cells, antibody-dependent cellular cytotoxicity (ADCC) has been shown to predict better HIV-1 protection. ADCC is initiated by the engagement of an Fc γ receptor CD16 with an Fc portion of the antibody, leading to phosphorylation of the CD3 ζ chain (CD3ζ) and Fc receptor γ chain (FcRγ) as well as downstream signaling activation. Though CD3ζ and FcRγ were thought to have overlapping roles in NK cell ADCC, several groups have reported that CD3ζ-mediated signals trigger a more robust ADCC. However, few studies have illustrated the direct contribution of CD3ζ in HIV-1-specific ADCC. To further understand the roles played by CD3ζ in HIV-1-specific ADCC, we developed a CD3ζ knockdown system in primary human NK cells. We observed that HIV-1-specific ADCC was inhibited by CD3ζ perturbation. In summary, we demonstrated that CD3ζ is important for eliciting HIV-1-specific ADCC, and this dynamic can be utilized for NK cell immunotherapeutics against HIV-1 infection and other diseases.
Introduction
Natural killer (NK) cells exert diverse effector functions that are critical for human immunodeficiency virus (HIV)-1 control. 1 –6 Specifically, antibody-dependent cellular cytotoxicity (ADCC) is a major contributor of HIV-1 control. 1,7,8 Others have elucidated that in the RV144 and VAX003 vaccine trials, individuals with better control of HIV-1 infection following vaccination exhibited higher levels of HIV-1-specific immunoglobulin G3 (IgG3) antibodies, which are potent inducers of NK cell ADCC. 7,9 Similarly in nonhuman primate (NHP) models, simian immunodeficiency virus (SIV) protection is strongly correlated with robust SIV-specific ADCC activities. 8 To trigger ADCC, the Fc portion of the antibody binds to CD16, a low-affinity Fc γ receptor IIIa, on NK cells, which induces phosphorylation of CD3 ζ chain (CD3ζ) and Fc receptor γ chain (FcRγ) adaptor molecules. 10 This event initiates subsequent signaling activation, which induces NK cell ADCC as a consequence. 11 –21
Due to the association of CD16 with both CD3ζ and FcRγ, 10,22 it has often been thought to trigger similar activation signals for NK cell ADCC. However, multiple lines of evidence indicate that NK cells preferentially utilize CD3ζ to exert more robust ADCC. 11,23 –26 One subset of human and rhesus macaque (RM) NK cells without FcRγ expression, referred to as g- or Δg NK cells, 11,23,25,27,28 is specialized in ADCC responses, and it primarily depends on CD3ζ for relaying signals from CD16. 11,29 Recently, Tuyishime et al. illustrated that the magnitude of CD3ζ phosphorylation positively correlates with NK cell ADCC potency in humans. 30 People experiencing systemic lupus erythematosus also exhibited lower CD3ζ expression in their NK cells, and their NK cell ADCC activity is positively associated with the levels of CD3ζ. 26 A recent study performed by Liu et al. documented that FcRγ knockout in primary NK cells using clustered regularly interspaced short palindromic repeats knockout technique did not perturb NK cell responses via CD16 stimulation. 31 While these reports imply the importance of CD3ζ in NK cell ADCC, few studies have directly elucidated its contribution to overall NK cell ADCC through modulation of CD3ζ expression by small interfering RNA (siRNA) gene knockdown. Specifically in HIV-1 infection, the link between CD3ζ and potent HIV-1-specific ADCC activities is yet to be definitively established. Hence, in order to fill this knowledge gap, we investigated whether the reduction of CD3ζ expression by siRNA knockdown can diminish HIV-1-specific NK cell ADCC.
Materials and Methods
NK cell expansion
Healthy human blood specimens were obtained from ZenBio Inc. and under Duke IRB protocol 00000873. Peripheral blood mononuclear cells (PBMC) were purified from healthy donor blood by Ficoll–Hypaque density gradient centrifugation. NK cells were then isolated from PBMC using a human NK Cell Isolation Kit (Miltenyi Biotec) following the manufacturer’s recommended protocol. NK cells were then resuspended in NK cell expansion medium comprised of NK magnetic-activated cell sorting medium (NK MACS medium; Miltenyi Biotec) supplemented with 5% human AB Serum (Thermo Fisher Scientific) and 30 ng/mL of recombinant human interleukin (IL)-15 (Miltenyi Biotec). Five days after culture, an additional NK cell expansion medium was supplemented, and cell numbers were counted. NK cell expansion medium was then added to the cell culture to maintain a cell concentration of 0.5 million cells/mL. Every 2–3 days, cell numbers were monitored, and the NK cell expansion medium was supplied so that the cell concentration was kept under 0.5 million cells/mL up until 17 days after culture initiation.
Single NK cell clones
NK cells were isolated from human PBMC by negative selection (human NK Cell Isolation Kit, Miltenyi Biotec). As feeder cells, we mixed fresh allogeneic PBMC and 8866 cell lines that are in an exponential proliferation phase (Sigma-Aldrich, St. Louis, MO) at a PBMC:8866 cell ratio of 10:1 and irradiated with the dose of 60 Gy. NK cells and feeder cells were cocultured in a cloning medium consisting of RPMI-1680 (Thermo Fisher Scientific), 5% human serum (Sigma-Aldrich), 1× minimum essential medium non-essential amino acids (MEM NEAA, Gibco), 1× sodium pyruvate (Gibco), 100 μg/mL kanamycin, 500 U/mL Roche recombinant human IL-2 (Sigma-Aldrich), and 1 μg/mL phytohemagglutinin (Fisher Scientific) and plated in 96-well plates so that 100 μL of culture in each well contained one NK cell. Fourteen days after culture, wells with proliferated NK cell clones (NKCL) were transferred to a 48-well plate, and additional cloning media were supplied every 2–3 days. The phenotype of NKCL was characterized by flow cytometry using the following antibodies: anti-CD3-AlexaFluor 700 (BD Pharmingen, clone SP34.2), anti-CD56-BV605 (BD Pharmingen, clone NCAM16.2), and anti-CD16-APC-Cy7 (BD Pharmingen, clone 3G8). To minimize the potential batch effects on mean fluorescence intensity (MFI) during our experiments, we also employed multiple mitigation strategies. We first purchased the antibodies in bulk in order to use a single lot of antibodies for the entire duration of this study. We also ensured the quality control of the BD FACS Symphony, including the use of calibration beads, 32 daily to ascertain that the voltage, filter, and laser measurements were within consistent and acceptable values.
siRNA knockdown for CD3ζ
Expanded NK cells were pelleted and nucleofected with 5 µM Accell nontargeting siRNA #1 (Horizon Discovery) or 5 μM SMARTpool Accell CD3ζ siRNA (Horizon Discovery) using a P3 Nucleofection Kit (Lonza) and Amaxa 4D nucleofector (Lonza, pulse code CM137). 33,34 Nucleofected cells were then cultured in an NK cell expansion medium. For NKCL, cells were incubated in NK MACS medium with 5% human AB serum, 500 U/mL recombinant human IL-2 (R&D Systems), and 1 ng/mL of recombinant human IL-15 (Miltenyi Biotec) following nucleofection. Two days after nucleofection, the efficiency of CD3ζ knockdown and the levels of FcRγ expression were analyzed via flow cytometry by the staining of the following surface and intracellular markers: anti-CD3-BV786 (BD Pharmingen, clone SP34.2), anti-CD14-BUV737 (BD Pharmingen, clone M5E2), anti-CD20-BUV395 (BD Pharmingen, clone L27), anti-CD56-BV605 (BD Pharmingen, clone NCAM16.2), anti-HLADR-APC-Cy7 (BD Pharmingen, clone G46-6), anti-CD16-BUV496 (BD Pharmingen, clone 3G8), anti-Fc receptor γ chain (FcRγ)—Alexa Fluor 700 (conjugated in our laboratory from anti-FcεRI antibody γ subunit, EMD Millipore, rabbit polyclonal), and anti-CD247 (CD3ζ)-fluorescein isothiocyanate (FITC) (BioLegend, clone 6B10.2). A representative gating strategy is summarized in Supplementary Figure S3.
Calcein acetoxymethyl–based NK cell ADCC and killing assay
CEM.NKR.CCR5 cell lines (ATCC) were spinoculated with p24 90 ng of HIV-1 bronchoalveolar lavage (BaL, National Institutes of Health [NIH] AIDS reagent) per 2 × 105 cells at 1,200 g for 2 h; infected cells were cultured for 6 days in RPMI-1640 with 10% fetal bovine serum (Thermo Fisher Scientific) and 100 U/mL penicillin–streptavidin (Thermo Fisher Scientific) (R10). One day prior to the killing assay, a fraction of BaL-infected cells was stained with anti-HIV core antigen-FITC (Beckman Coulter, clone KC57), and the percentage of HIV+ cells was analyzed by flow cytometry. We confirmed that >99% of CEM.NKR.CCR5 cells are CD4+, and, following spinoculation, we observed that 30%–60% of cells became BaL+ the day before the experiment. BaL-uninfected or BaL-infected cells were then stained with 10 μM calcein acetoxymethyl (CAM) (Invitrogen) for 1 h at 37°C. Stained cells were washed twice and coated with 10 μg/mL human anti-HIV-1 envelope-specific antibody PGT121 or anti-human CD4 antibody (clone CD4R1) 11 for 15 min at room temperature. Following the incubation, nontargeting siRNA or CD3ζ siRNA-nucleofected expanded NK cells or NKCL pool were cocultured to achieve a 10:1 Effector:Target (E:T) ratio in a total volume of 200 μL R10 per well. Four hours after coculture, supernatant from the culture was harvested and CAM signals were measured using Nivo plate readers (PerkinElmer). To measure spontaneous CAM release, target cells without NK cells were prepared. Four hours after coculture, cells were pelleted, and the supernatant was collected, and the levels of CAM release (excitation 480 nm, emission 535 nm) were quantified by Nivo Plate Reader (PerkinElmer). ΔCAM was calculated by subtracting fluorescence from the condition without NK cells. We confirmed that data from expanded NK cells and NKCL are comparable (data not shown), and both datapoints were plotted in the same figure as a separate symbol.
Annexin V-based NK cell ADCC and killing assay
CEM.NKR.CCR5 cell lines were stained with 50 nM carboxyfluorescein succinimidyl ester (CFSE, Thermo Fisher Scientific) for 20 min. After staining, cells were incubated with anti-human CD4 antibody (clone CD4R1, NHP reagents) at room temperature for 15 min. Following antibody coating, nontargeting siRNA or CD3ζ siRNA-nucleofected expanded NK cells were added and cultured for 4 h. Following coculture, the cells were stained with anti-Annexin V-APC (BD Biosciences, clone), and the % Annexin V+CFSE+ cells were measured by flow cytometry. The %ADCC was calculated as follows: %ADCC = (%Annexin V+ cells with an antibody) − (%Annexin V+ cells without an antibody). For an HIV-1-specific antibody titration, BaL-infected CEM.NKR.CCR5 cell lines were treated with 5-fold serial dilution of human anti-HIV-1 env antibodies PGT121, VRC01, 10E8, and 35022 starting at 100 μg/mL. Cells were then incubated with primary human NK cells for 4 h, and %Annexin V+CFSE+ cells were measured by flow cytometry.
Statistical analysis
GraphPad Prism 9.0 software (GraphPad Software) was used to evaluate statistically significant differences. p-Values <.05 were considered statistically significant. Wilcoxon signed-rank tests were used to assess statistically significant differences for paired analyses.
Results and Discussion
To delineate the contribution of CD3ζ in NK cell ADCC, we aimed to establish a CD3ζ knockdown system in expanded primary human NK cells or NKCL using siRNA. We first investigated a transient CD3ζ knockdown approach using CD3ζ siRNA. Nucleofection of CD3ζ siRNA was able to reduce CD3ζ expression, whereas FcRγ expression was only minimally altered (Fig. 1A–C, p < .05). The levels of CD3ζ and FcRγ were not strongly correlated following CD3ζ knockdown, further supporting the specificity of this strategy (Supplementary Fig. S1). We also measured the levels of surface CD16 expression; while there was a statistically significant difference, the downregulation of CD16 expression was limited by CD3ζ knockdown (Fig. 1D). Altogether, we confirmed that CD3ζ knockdown by siRNA can specifically assess the contribution of CD3ζ in NK cell function with limited off-target effects.

CD3ζ knockdown specifically downregulates CD3ζ expression in primary human NK cells. Expanded primary human NK cells or pooled NKCL were nucleofected with scrambled siRNA (si-Ct) or CD3ζ siRNA and the levels of CD3ζ in live CD3− CD14−CD20−CD56+CD16+ cells were measured by flow cytometry,
Using this siRNA knockdown strategy, we investigated whether CD3ζ reduction will impair NK cell ADCC responses. First, to evaluate the effect of CD3ζ knockdown on NK cell ADCC against abundantly expressed antigens, we used uninfected CEM.NKR-CCR5 cell lines 35 as target cells and coated cells with anti-human CD4 antibody to elicit NK cell ADCC. 11 We demonstrated that CD3ζ knockdown significantly decreases NK cell ADCC activity against target cells as measured by intracellular CAM release (Fig. 2A, p < .05). While it was not statistically significant, our Annexin V-based NK cell ADCC assay also revealed an analogous reduction of NK cell ADCC activities by CD3ζ knockdown (Fig. 2B). In summary, we illustrated that CD3ζ knockdown specifically perturbs NK cell ADCC against an antigen with high endogenous expression.
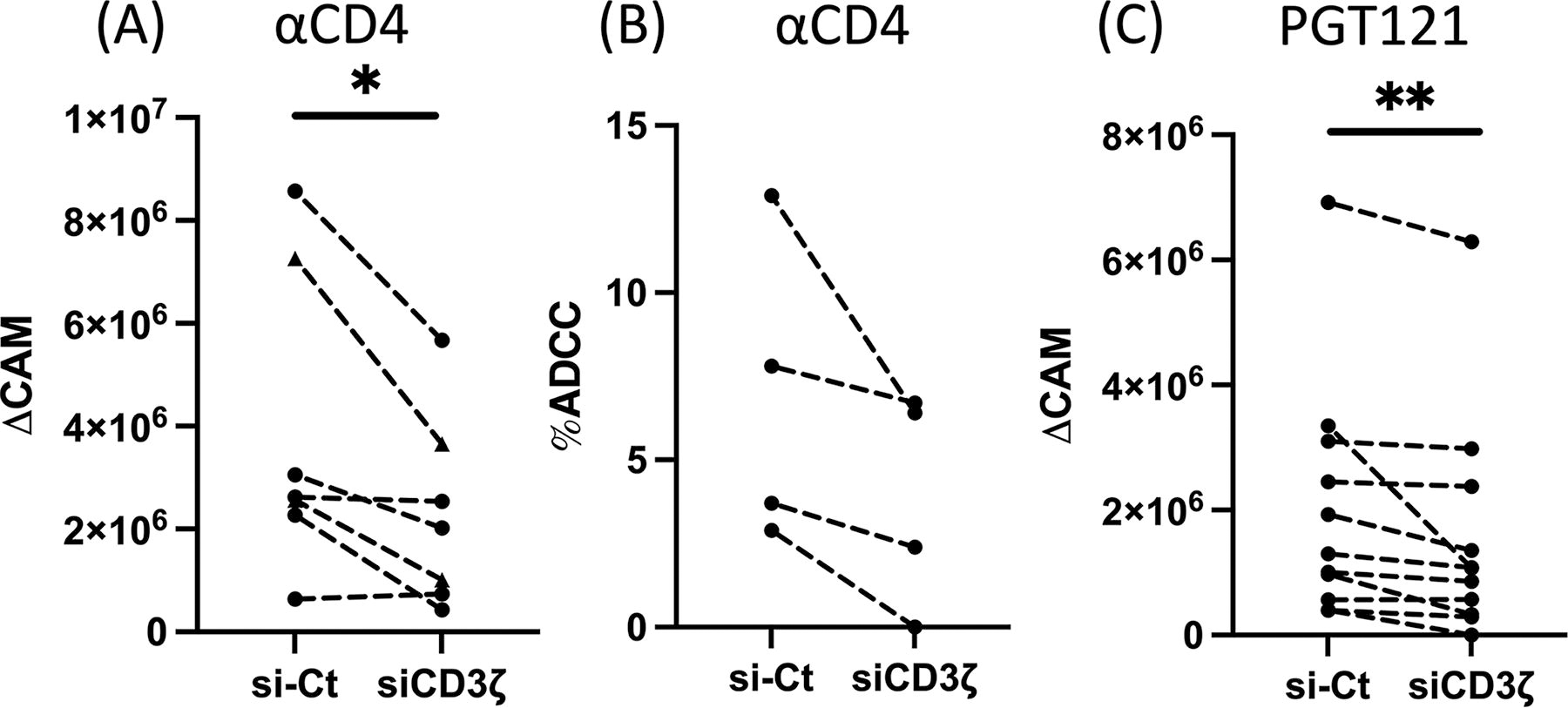
CD3ζ reduction diminishes HIV-1-specific NK cell ADCC. Expanded primary human NK cells (triangle) or the mixed NKCL (circle) were nucleofected with scrambled siRNA (si-Ct) or CD3ζ siRNA for two days. Cells were then cocultured with BaL-uninfected CEM.NKR.CCR5 cells coated with anti-CD4 antibody
In order to evaluate the impact of CD3ζ knockdown on HIV-1-specific ADCC, we next tested the potency of HIV-1-specific ADCC by HIV-1 BaL-infected CEM.NKR-CCR5 cells. First, we investigated which of the commonly used HIV-1-specific antibodies (PGT121, VRC01, 35022, and 10E8) 36 elicited the most robust HIV-1-specific NK cell ADCC at the optimal concentrations. Our ADCC assay elucidated that PGT121 triggered the most potent HIV-1-specific ADCC within the concentration of 5–20 μg/mL (Supplementary Fig. S2). Using PGT121, we next tested if the knockdown of CD3ζ abrogated HIV-1-specific ADCC. 36 Consistent with Figure 2A, HIV-1-specific NK cell ADCC was significantly diminished by loss of CD3ζ expression (Fig. 2C, p < .01). Taken together with our prior findings, these data illustrate that CD3ζ contributes to eliciting HIV-1-specific NK cell ADCC.
Having developed a CD3ζ knockdown strategy in primary human NK cells using siRNA, we were able to confirm that CD3ζ siRNA does not significantly alter FcRγ expression. Using this technique, we illustrated that HIV-1-specific NK cell ADCC was perturbed by decreasing CD3ζ expression. Our findings represent a critical addition to our understanding of NK cell biology, specifically for ADCC activity. Previous studies indicate the association between robust ADCC responses and CD3ζ levels in NK cells, 11,23 –26 but the importance of CD3ζ in ADCC has never been characterized by knockout/knockdown techniques. Our data support the argument that CD3ζ is a crucial molecule in harnessing NK cell ADCC by directly modulating CD3ζ expression.
While our findings provide novel insights into NK cell ADCC, we acknowledge that this study has several limitations. First, HIV-1-specific ADCC potency was limited in our experiments, presumably due to the antibody clones that were used in this study. Recently, it was demonstrated that the combination of neutralizing and non-neutralizing antibodies against HIV-1 can further ameliorate HIV-1-specific ADCC responses. 37 Therefore, the role of CD3ζ in HIV-1-specific NK cell ADCC can be validated in an in vitro system with the optimal combination of antibodies against HIV-1. Moreover, the roles of CD3ζ in NK cell ADCC were mostly demonstrated in vitro in this study. Thus, follow-up studies using samples from people living with HIV (PLWH) or NHP models are necessary to validate our findings ex vivo and in vivo.
Our presented data also provide a novel insight into which method can potentially be applied for NK cell immunotherapeutics. To date, numerous NK cell-based strategies, including chimeric-antigen receptor NK cells, bispecific killer engagers, or trispecific killer engagers, demonstrated their effectiveness against cancers and multiple pathogens. 38 –41 The potency of NK cell therapies is also increasingly appreciated in HIV-1 infection. 41 These approaches are further augmented by gene editing strategies to enhance NK cell activities. 42,43 Our results imply that the modulation of CD3ζ expression by siRNA knockdown or lentiviral transduction could be implemented as an additional tool to specifically augment NK cell-mediated ADCC in these strategies to kill HIV-1-infected cells more effectively. Altogether, we highlighted that NK cells prefer to utilize CD3ζ for inducing HIV-1 ADCC. These results can be translated to develop more potent NK cell ADCC for HIV-1 cure strategies and other disease treatments.
Footnotes
Authors’ Contributions
S.S., R.K.R., and S.J. conceptually developed the initial project, and S.S. performed a majority of experiments. E.L., M.A.C., A.P., S.B.G., and H.B. generated NKCL, and M.A.C. and A.P. helped the expansion of primary human NK cells. K.K. and C.M. developed the protocol for CEM.NKR-CCR5 cell line infection with BaL, prepared the laboratory stock of BaL, and performed HIV-1-specific antibody-titration experiments. Y.L. provided anti-HIV-1 Env antibodies. S.S. and G.W. drafted the article, and all the authors contributed to the final version.
Data Availability
The data generated for this study are available upon request to the corresponding author.
Author Disclosure Statement
All authors report no financial conflicts of interest.
Funding Information
This research was supported by NIH grants:
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.