
Abstract
HIV/AIDS cannot be cured because of the persistence of the viral reservoir. Because of the complexity of the cellular composition and structure of the human organs, HIV reservoirs of anatomical site are also complex. Recently, although a variety of molecules have been reported to be involved in the establishment and maintenance of the viral reservoirs, or as marker of latent cells, the research mainly focuses on blood and lymph nodes. Now, the characteristics of the viral reservoir in tissue are not yet fully understood. In this study, various tissues were collected from SIVmac239-infected monkeys, and the level of total SIV DNA, SIV 2-LTR DNA, and cell-associated virus RNA in them were compared with character of the anatomical viral reservoir under early treatment. The results showed that short-term combination antiretroviral therapy (cART) starting from 3 days after infection could significantly inhibit viremia and reduce the size of the anatomical viral reservoir, but it could not eradicate de novo infections and ongoing replication of virus. Moreover, the effects of early cART on the level of total SIV DNA, SIV 2-LTR DNA, and cell-associated virus RNA in different tissues were different, which changed the size distribution of viral reservoir in anatomical site. Finally, the contribution of nonlymphoid tissues, especially liver and lung, to the viral reservoir increased after treatment, while the contribution of intestinal lymphoid to the viral reservoir significantly reduced. These results suggested that early treatment effectively decreased the size of viral reservoir, and that the effects of cART on the tissue viral reservoir varied greatly by tissue type. The results implied that persistent existence of virus in nonlymphoid tissues after short-term treatment suggested that the role of nonlymphoid tissues cannot be ignored in development strategies for AIDS therapy.
Introduction
The persistence of a viral reservoir, especially latent viral reservoir, is a major obstacle to HIV-1/AIDS cure. Despite long-term antiretroviral therapy, the viral reservoir persists and is a source of re-emerging virus after therapy interruption. 1 –3 Infected cells, such as resting memory CD4+ T cell, macrophages, and astrocytes, contain intact viral genome and can release infectious virus persistently or under special environment. 3 These cells reside in different anatomic site/organizations and have special characteristics. Meanwhile, some anatomical site has a specific structure. They can protect virus against drug or clearing of immune. Both cellular composition and structure of organs increased the complexity of anatomical viral reservoir during therapy. 4,5 Redefining the viral reservoirs under treatment may provide important theoretical basis for a new treatment strategy.
Once infected, HIV-1/SIV rapidly establishes viral reservoir, 6 and rapidly disseminates from the site of inoculation to distal sites. 7 The virus can be detected in almost all tissues at end-stage disease. 8 Now, due to the ineffectiveness of the vaccine prevention and intervention, HIV/AIDS therapies mainly rely on combined antiretroviral therapy (cART). Antiviral therapy, which is usually being initiated at the time that CD4+ T cell number is lower than 400/μL can suppress viremia to lower than the limit of detection. However, nonsuppressible HIV-1 viremia or trace levels of viremia (residual viremia) was found in many of patients that received treatment when using more sensitive assays. 9,10 It implied that there was ongoing virus production in patients receiving cART. Recently, Mohammadi et al. identified the critical host and viral mediators of nonsuppressible HIV-1 viremia. 10 In principle, there are many sources of residual viremia, such as a low degree of ongoing viral replication, the release of virus from latently infected cells that have become activated, or the release of virus from other stable reservoirs. 3 De Scheerder et al. reported that rebound viruses can originate from multiple cellular and anatomical compartments and suggested that cure strategies should take into account the lack of prominent HIV rebound origin. 11 However, due to the limitation of tissue source, the research on the anatomical compartments of the viral reservoir under the cART mainly focuses on lymphoid tissues (such as lymph node, spleen), while the nonlymphoid tissues, such as reproductive and endocrine system, receive less attention. Recently, it has been suggested that suspected cases or high-risk groups should be tested and confirmed cases should get treatment as soon as possible. 12 Some studies also found that the size of viral reservoir in lymph nodes of some patients decreased during the treatment, and long-term virological remission could still be achieved after the interruption of early antiretroviral therapy. 13 Okoye et al. reported that early antiretroviral therapy restricts the establishment of SIV reservoir, thereby delaying or preventing posttreatment viral rebound. 14 However, there are still controversial conclusions on whether early treatment can achieve functional cure. Because early antiretroviral treatment, even as early as 3 days after infection (DAI), only delays viral rebound after treatment discontinuation. 6,14 Although both studies showed that early antiretroviral therapy reduced the size of SIV reservoir in peripheral blood and lymph node (LN), there is still a lack of systemic study of viral reservoir in lymphoid and nonlymphoid tissues.
In this study, we collected a variety of tissues from SIVmac239 acute infected Chinese rhesus macaques (ChRMs), which received or did not receive cART at 3 DAI. Then we detected loads of the 2-LTR (two-long-terminal-repeat) DNA, the total SIV DNA and the cell-associated viral RNA (CA-vRNA), compared the size of viral reservoir and analyzed change and distribution of anatomical viral reservoir to characterize the viral reservoir in anatomical sites under early treatment. Our results suggest that early cART can reduce the size of the viral reservoir, with effects that varied by tissue type, thereby altering the distribution of the anatomical viral reservoir of ChRMs infected with SIVmac239. The results also highlighted the importance of nonlymphoid tissue contribution to the viral reservoir under cART.
Materials and Methods
Animals and infection
Six 8- to 10-year-old adult male rhesus macaques (Macaca mulatta) of Chinese origin were included in this study. They were inoculated intravenously with a thousand 50% tissue culture infective dose SIVmac239 and randomly divided into control group (treatment-naive group) and cART treatment group. Each group has three monkeys. The detailed information of the monkeys are shown in Table 1. In this study, all operations were accorded with the recommendations of the Weatherall report, “The use of nonhuman primates in research” was approved by the Ethics Committee of Kunming Institute of Zoology, Chinese Academy of Sciences (approval number: IACUC18012).
Animal Information
PMPA, emtricitabine; FTC, Emtritabine; RAL, Raltegravir.
cART regimen
The cART regimen consists of two nucleoside reverse transcriptase inhibitors (Emtritabine, FTC, 50 mg/kg/SC, SID; tenofovir, PMPA, 30 mg/kg/SC, SID) and one integrase inhibitor (Raltegravir, RAL, 100 mg/bid, PO). 15 The cART was initiated at 3 DAI and continued until euthanasia.
Sample collecting
EDTA K2 anticoagulated peripheral blood was collected from the leg vein. Plasma and peripheral blood mononuclear cells (PBMCs) were separated. All the animals were euthanized and necropsied as described previously. 16 LNs (mesenteric LN, inguinal LN, axillary LN, submaxillary LN), liver, spleen, lung, kidney, gastrointestinal tract (stomach, ileum, duodenum, jejunum, colon, rectum, and cecum), cerebrum, cerebellum, testicle, bladder, penis, prostate, epididymis, thyroid, and adrenal gland were collected (without PBS perfusion) and cryopreserved in liquid nitrogen immediately after collection.
DNA and RNA extraction from PBMCs and tissues
Total DNA was extracted from PBMCs and tissues with a QIAamp DNA Mini Kit according to the manufacturer’s instructions. Tissues were homogenized using a Tissue Lyser in the presence of TRIzol Plus (TaKaRa). RNA was purified according to the manufacturer’s instructions. Nucleic acid concentration was measured using a NanoDrop 2000. RNA and DNA were frozen in −80°C until use.
Quantitation of plasma viral load, total SIV DNA load, 2-LTR DNA, and CA-vRNA load
As previously reported, 17 plasma viral load (pVL) and CA-vRNA were measured by SIV gag RNA load. Total SIV DNA level was measured by SIV gag DNA load. CCR5 gene, which only has two copies in each cell, was included as internal reference for total SIV DNA load and SIV 2-LTR load. 18 RPL13A RNA was used as internal reference for RNA detection. Total SIV DNA, 2-LTR DNA, and CCR5 gene in 1–5 μg of DNA were quantitated using real-time PCR (ABI7500F) as described by Tian et al. and Lee et al. 16,19 SIV gag probe, 5′-FAM-CTTCTGCGTGAATGCACCAGATGACGC-TAMRA-3′, SIV 2-LTR probe, 5′-Quasar 670 (Cy5)-AGCCGCCGCCTGGTCAACTCG-BHQ-3′, and CCR5 probe: 5′-FAM-AACACAGCCACCACCCAAGTGATCA-TAMRA-3′ were used. Total SIV DNA load or 2-LTR load was normalized as the total provirus copies or 2-LTR copies in 106 PBMCs. CA-vRNA in 1 μg total RNA was detected using primer and probe as pVL above. RPL13A forward primer, 5′-TCACGAGGTTGGCTGGAAGT-3′, RPL13A reverse primer, 5′-GATCTTGGCTTTCTCCTTCCTCTT-3′, and RPL13A TaqMan probe, 5′-Cy3-CCAGGCAGTGACAGCCACCTTGG-BHQ-3′ were used to detect RPL13A. RNA or plasmids with known copy numbers were used as standard sample. For detection of total SIV DNA, 2-LTR DNA, and CCR5 gene, standard samples were diluted in nuclease-free water, ranging from 0.1 to 109 copies/10 μL. The limit of detection was 1 copy/well/10 μL sample. For detection of the CA-vRNA and RPL13A, the standard sample was diluted in nuclease-free water with carrier RNA, ranging from 0.1 to 108 copies/10 μL. The limit of detection (LOD) was 1 copy/well/10 μL. For plasma viral detection, the standard sample was diluted in nuclease-free water with carrier RNA, ranging from 0.37 to 2.96 × 105 copies/8.75 μL. The stable LOD was 2.96 copy/well/8.75 μL. So, the LOD of pVL was 74 copies/mL plasma. CA-vRNA load was normalized as the total SIV gag copies/108 copies of RPL13A. Standard curves of quantification for each viral parameter are shown in Figure 1.

Standard curves of quantification for each parameter. Plasma viral RNA
Statistical analyses
All statistical analyses were conducted with GraphPad Prism 8.0 software. Score of the SIV DNA, SIV RNA, and SIV 2-LTR DNA level of tissues and the center of the data were calculated in R version 4.2.1 using the following function, respectively: scale() and attr(,“scaled:center”). The Mann–Whitney test and unpaired t test with Welch’s correction were used for comparisons within the groups. Values of p < 0.05 were considered statistically significant.
Results
Early cART effectively suppressed viremia in SIV-infected ChRMs
As shown in Figure 2, the pVL of control subjects was around the limit of detection (70 copies/mL) at 3 DAI. Then the pVL increased rapidly, reached peak value within 2 weeks after infection, and then decreased slowly. The peak values of pVL ranged from 5.87 × 106 to 2.66 × 107 copies/mL. The pVL of the patients in early cART group was similar to that in control at 3 DAI, but increased only slowly thereafter, reaching a peak at 7 DAI and then declining. The peak of pVL was slightly above the detection limit while receiving cART. On the day of euthanasia, the pVL of all early cART subjects was below the detection limit. These results suggested that early cART effectively inhibited viremia in SIV-infected ChRMs.

Longitudinal changes in plasma viral load of SIV-infected Chinese rhesus macaques. Six rhesus macaques were challenged with SIVmac239. Three of them were treated with TDF, FTC, and RAL at day 3 postinfection (red), the other three were served as treatment-naive control (black). Viral RNA was extracted from plasma and viral load was determined by Q-PCR and identified as copy number of Gag RNA per milliliter plasma. Three independent experiments were included for each animal and each time point, and each sample was measured three times in the experiment.
Early cART reduced total SIV DNA, CA-vRNA, and 2-LTR DNA levels in PBMCs
Total SIV DNA, also known as cell-associated SIV DNA, was considered to be a marker of the HIV reservoir. 20 Two-LTR circle was used as indicator of recent infection events. 21 In addition, CA-vRNA was cited as a dynamic biomarker of viral persistence. 22 So, we evaluated the level of total SIV DNA, 2-LTR, and CA-vRNA to character the dynamic change of virus replication in PBMCs under cART. In line with the trend in pVL, the level of total SIV DNA and CA-vRNA in PBMCs of control group rapidly elevated during 7–10 DAI and then decreased slowly. The levels of total SIV DNA and CA-vRNA in early treatment group were comparable to those of the control group at 3 DAI (Fig. 3A, B and C). Thereafter, the levels of total SIV DNA and CA-vRNA increased slowly under cART pressure and peaked between 7–14 DAI, and then they decreased slowly (Fig. 3A, B, and C). When accumulative level was shown, the levels of the total SIV DNA and 2-LTR DNA in PBMCs of both the treated groups and the treatment-naive group quickly accumulated during 14 DAI, then declined, whereas the levels in the treated group were consistently lower than those in the untreated group (Fig. 3D and E). Early treatment also significantly reduced CA-vRNA accumulation, but it did not alter the trend of CA-RNA level’s change, as CA-vRNA accumulated to a peak and then remained at high levels in both groups regardless of treatment. (Fig. 3C and F). In summary, early cART reduced the size of the viral reservoir, but there may be low-level de novo infection and the viral reservoir decays slowly under early treatment.
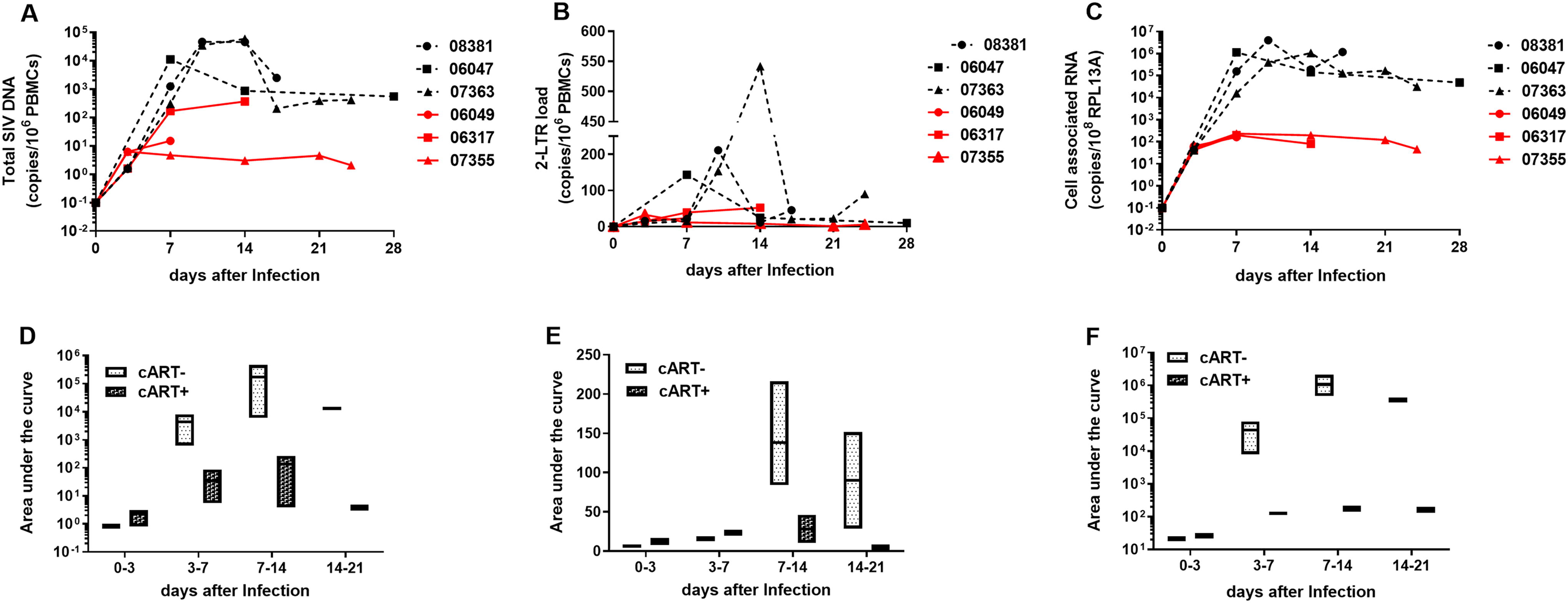
Longitudinal changes of total SIV DNA, 2-LTR DNA, and cell-associated viral RNA level in peripheral blood mononuclear cells from SIV-infected Chinese rhesus macaques. DNA and RNA were extracted from peripheral blood mononuclear cells of SIV-infected Chinese rhesus macaques, and then total SIV DNA
Early cART significantly reduced total SIV DNA, CA-vRNA, and 2-LTR DNA levels in tissues
To investigate the characteristics of SIV replication in tissues under the cATR stress, we collected different tissues from each subject, and assessed the level of total SIV DNA, CA-vRNA, and 2-LTR DNA. It was shown that total SIV DNA (Fig. 4), CA-vRNA (Fig. 5), and 2-LTR DNA (Fig. 6) were detected in all tissues from both control and treated group, and early cART significantly reduced them in most tissues (p < 0.0001; p < 0.0001; p < 0.0001). In total SIV DNA detection, the highest total SIV DNA load in the control group was found in LNs, liver, gastrointestinal tract, and spleen, while the lowest total SIV DNA load was found in the brain and reproductive system. Compared with the control group, early treatment reduced the total SIV DNA load of LN, liver, and gastrointestinal tract by more than 100-fold; the total SIV DNA load of spleen, kidney, adrenal, lung, thyroid, and cerebellum by more than 10-fold; and the total SIV DNA load of bladder, cerebrum, and reproductive system by less than 10-fold (Fig. 4). These results suggested that the effect of early cART on tissues’ viral reservoir depended on type of tissue. Similar to the total SIV DNA level, CA-vRNA level of control group was highest in LNs, liver, gastrointestinal tract, and spleen and was lowestlevel in the brain and reproductive system. After treatment, CA-vRNA level in most tissues changed significantly, with a decrease of 306–20,489-folds in lymph node, small intestine, stomach, kidney, colon, and bladder, and with a decrease of 10–100-folds in liver, epididymis, rectum, brain, thyroid, adrenal, and testicle. In addition, the CA-vRNA level in penis and lung was least affected by treatment (Fig. 5). Therefore, these results suggested that early cART significantly reduced the viral persistence in anatomical site, but its effects varied greatly with the type of tissue. Similar to total SIV DNA detection, the highest level of 2-LTR DNA in control group was found in LN, liver, gastrointestinal tract, spleen, brain, and reproductive system. After the cART, 2-LTR DNA levels in LNs, liver, gastrointestinal tract, and kidney were reduced by more than 10-fold, while 2-LTR level in brain, spleen, lung, adrenal, penis, prostate, and thyroid was only reduced by less than 10 times. Notably, 2-LTR DNA levels in the bladder, epididymis, and brain were comparable between the treatment and control groups, while 2-LTR DNA levels in the prostate were significantly higher in the treatment group than in the control group (Fig. 6). Therefore, these results implied that early cART inhibited infection in most lymph tissue, but recent infection remains, especially in nonlymphoid tissues, such as bladder, epididymis, cerebrum, and prostate.

Total SIV DNA load in anatomical sites. DNA was extracted from tissues of SIV-infected Chinese rhesus macaques. Then SIV gag DNA in 1ug total DNA was quantified by Q-PCR. CCR5 gene was used as internal reference. Total SIV DNA load was indicated as the total SIV Gag copies/106 CCR5 gene or cells. Black, control group. Gray, early cART group. At least three independent experiments were included for each animal and each tissue, and each sample was measured three times in the experiment. All data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001.

Cell-associated viral RNA load in anatomical sites. RNA was extracted from tissues of SIV-infected Chinese rhesus macaques. Then SIV gag RNA in 1ug total RNA was quantified by Q-PCR. RPL13A gene was used as internal reference. Cell-associated SIV RNA load was indicated as the total SIV Gag copies/108 RPL13A RNA. Black, control group. Gray, early cART group. At least three independent experiments were included for each animal and each tissue, and each sample was measured three times in the experiment. All data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001.
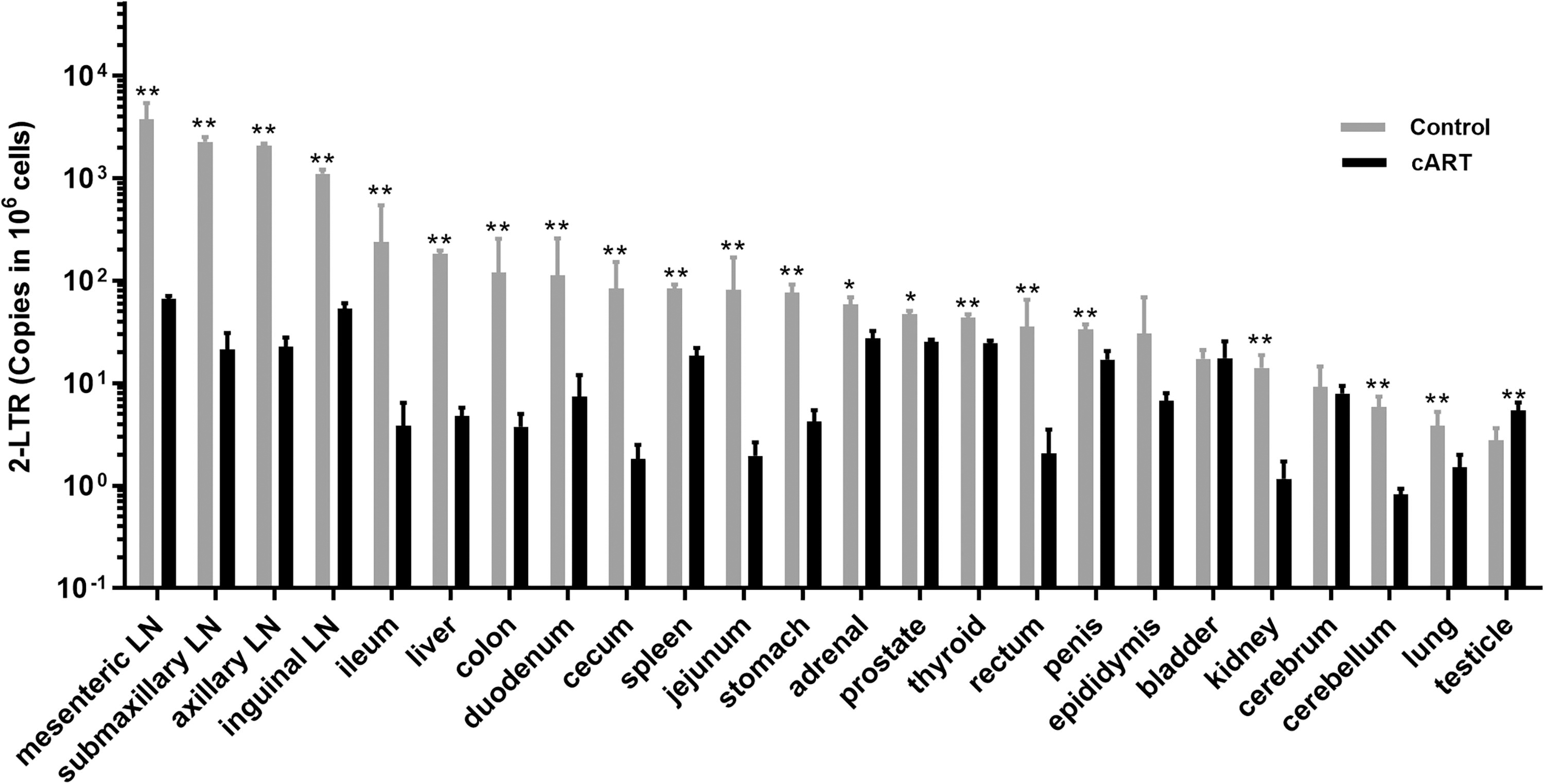
2-LTR DNA load in anatomical sites. DNA was extracted from tissues of SIV-infected Chinese rhesus macaques. Then 2-LTR DNA in 1ug total DNA was quantified by Q-PCR. CCR5 gene was used as internal reference. 2-LTR load was indicated as the total 2-LTR copies/106 CCR5gene. Black, control group. Gray, early cART group. At least three independent experiments were included for each animal and each tissue, and each sample was measured three times in the experiment. All data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001.
In summary, these results suggest that short-term early cART effectively reduced the size of viral reservoir in anatomical sites, with an effect that depends on the tissue type, but cannot completely prevent viral persistence and recent infection in tissue.
Early cART altered the size distribution of anatomical viral reservoir
To further determine the impact of early cART on spatial distribution of viral reservoirs, we calculated the score of the total SIV DNA, SIV RNA, and SIV 2-LTR level in tissues to normalize the data. The scores are shown in Table 2. It was shown that early treatment obviously changed the order of the total SIV DNA load, CA-vRNA level, and 2-LTR level in tissue from cART-treated monkeys (Fig. 7A and B). In control group, the lymphoid tissues (e.g., LNs and spleen) scored highest for both total SIV DNA load (red in Fig. 7A) and CA-vRNA levels (green in Fig. 7A), while most nonlymphoid tissue (brain, testicle, epididymis) scored lowest. With the cART, the score of total SIV DNA level (red in Fig. 7B) in inguinal LN and submaxillary LN was seriously dropped, whereas the scores in liver, epididymis, and cerebrum were significantly increased. Therefore, the highest total SIV DNA loads were found in the liver, mesenteric LN, epididymis, cerebrum, spleen, and testicle in the treatment group (Fig. 4). Consistent with the total SIV DNA level, after treatment, CA-vRNA scores were obviously decreased in the lymph tissue (e.g., LNs and spleen), but increased in lung and rectum. Thus, CA-vRNA level of lung, mesentery LN, rectum, inguinal LN, spleen, axillary LN, adrenal, liver, submaxillary LN, and bladder ranked the top 10 among all tissues in cART subjects (Fig. 5). In a ranking of 2-LTR abundance, the highest 2-LTR levels were found in LNs of the control group, while the lowest levels were found in most nonlymphoid tissues (e.g., brain, epididymis, prostate, penis, bladder, lungs, and kidneys). After treatment, 2-LTR levels in LNs, spleen, adrenal, prostate, thyroid, penis, and bladder were higher than those in other sites (Fig. 6). Thus, the top five tissues with the highest 2-LTR levels were the mesenteric LN, inguinal LN, adrenal, prostate, and thyroid (Fig. 6). When a score of total SIV DNA, CA-vRNA, and 2-LTR DNA levels were joined, the control group had the highest scores in LNs, spleen, and liver, while the treatment group had the highest scores in mesenteric LNs, lung, liver, inguinal LN, epididymis, axillary LN, brain, thyroid, adrenal, and spleen (Fig. 7B). In all, early cART significantly changed the size distribution of anatomical viral reservoir and the contribution of nonlymph tissue to viral reservoir.
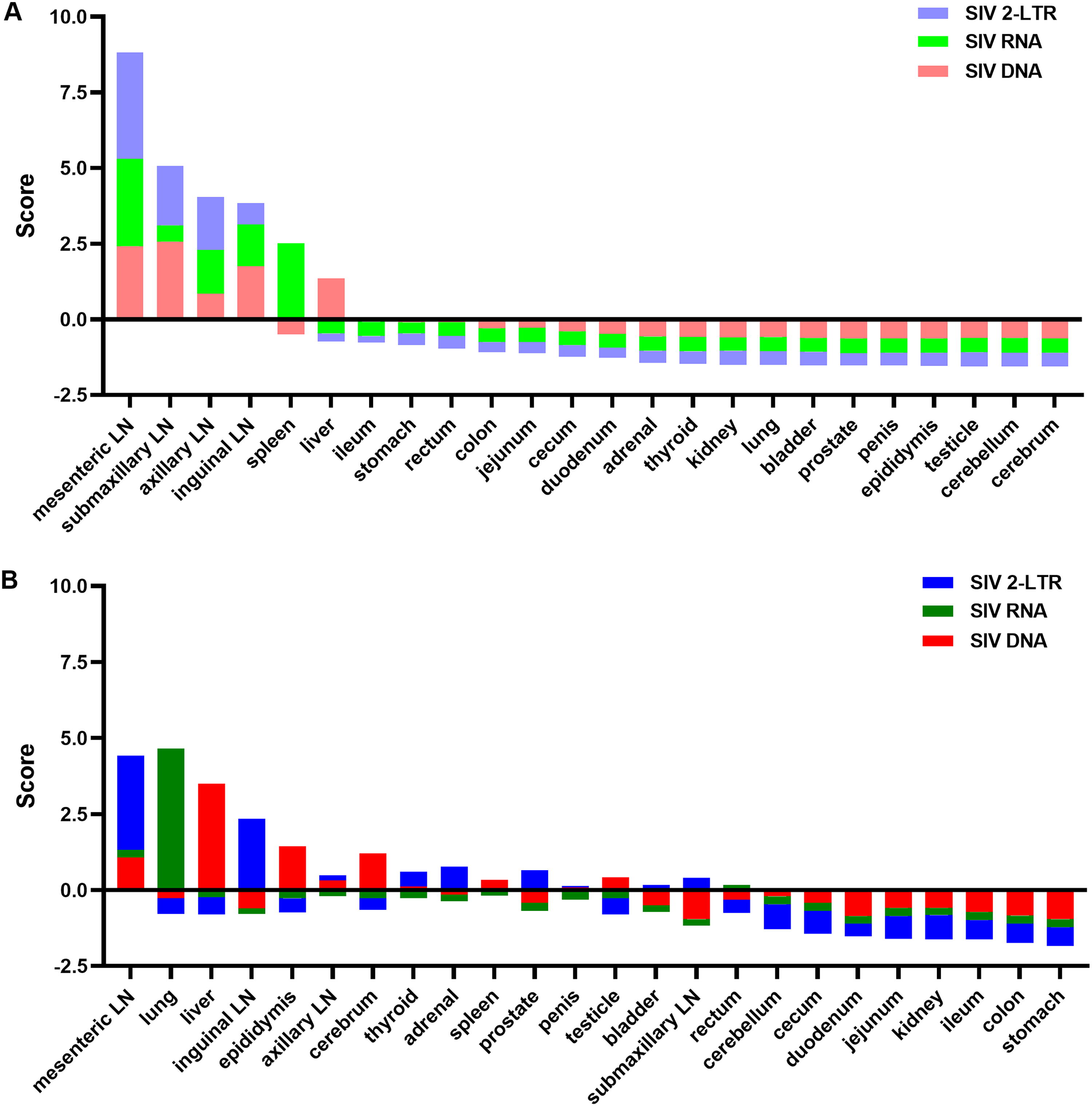
The ranking score of total SIV DNA load, cell-associated SIV RNA load, and 2-LTR DNA load in SIV-infected Chinese rhesus macaques. Scores of SIV DNA level (red), cell-associated SIV RNA level (green), and SIV 2-LTR DNA level (blue) in tissues from control group
The Score of Viral Distribution
The mean of each tissue is shown.
cART, combination antiretroviral therapy; LN, lymph node.
Discussion
Early antiretroviral therapy limits SIV reservoir establishment, thereby delaying or preventing post-treatment viral rebound. 14 In this study, we found that short-term early cART treatment significantly inhibited pVL, and suppressed total SIV DNA load, CA-vRNA level, and 2-LTR level in PBMCs and anatomical sites of SIVmac239-infected ChRMs. Nevertheless, after treatment initiation, total SIV DNA load, 2-LTR level and CA SIV RNA level transiently rose. Then, total SIV DNA load and 2-LTR level decreased slowly, whereas the CA-vRNA level did not change significantly. The impact of short-term early cART on the level of tissue total SIV DNA, CA SIV, and 2-LTR varied depending on the type of tissue, consequently leading to alterations in the distribution of anatomical viral reservoirs. These results implied that short-term early cART effectively inhibited the ongoing infection and the viral transmission, decreased the size of viral reservoir, and altered the size distribution of anatomical viral reservoir in SIVmac239-infected ChRMs.
Following infection, HIV/SIV rapidly establishes viral reservoir and spreads throughout the entire body. 3 The viral reservoir cannot be eradicated by existing treatment and is an obstacle of HIV cure. It is very difficult to accurately quantify tissue viral reservoir. Plasma virus, total SIV DNA virus, 2-LTR DNA, and CA-vRNA are products of viral replication and are associated with the persistence of virus during therapy. 23 –25 The 2-LTR are extrachromosomal products of failed HIV/SIV integration. 24 This episomes has been reported to be useful surrogate markers of viral replication. 25 In our study, we found a slow and delayed decline in pVL, total SIV DNA, and 2-LTR DNA levels in PBMCs during cART. A delayed and slow decline in pVL was also observed in monkeys receiving intravenous SIVmac239X and antiviral therapy. 14 However, Whitney et al. reported that pVL rapidly decreased below the detection limit after early treatment. 6 This difference may be caused by differences in the route of inoculation and the cART regimen. However, we have no evidence to suggest that RAL is less effective than dolutegravir in monkeys. Within 4 days after treatment, pVL increased transiently and remained low thereafter, possibly due to the activation of infected cells, which continued to transcribe viral genes and release new virus. 3 This was also supported by persistent CA-vRNA levels in PBMCs and anatomical sites. Consistent with pVL, total SIV DNA DNA and 2-LTR DNA levels also weakly increased during 3 DAI and then slowly decreased. The elevated 2-LTR level may be due to the accumulation caused by recent infection and long half-life of 2-LTR DNA. 26 However, the increase of 2-LTR DNA level implied de novo replication of virus and incomplete impeding of reverse transcription. Furthermore, given that total SIV DNA represented by SIV gag DNA includes incomplete reverse transcript, circular SIV DNA (1-LTR DNA and 2-LTR DNA) and integration SIV DNA, it is not surprising that total SIV DNA continues to increase. Therefore, combined therapies with multiple drugs targeting different steps of viral replication may have better therapeutic effect. Under the rigorous inhibition of viral replication by multidrug combination therapy and the activation and death of infected cells, the source and production of infectious virus particles decreased, the input of total SIV DNA and 2-LTR DNA decreased, and finally the tissue level of provirus and 2-LTR decreased. Total SIV DNA load, 2-LTR DNA level, and CA-vRNA level in almost all tissues also decreased in the early treatment group. Nevertheless, we do not rule the role of host immune response activated by exogenous DNA in the clearance of cytoplasmic SIV DNA. 27 Therefore, although short-term treatment can greatly suppress the dissemination of virus and decreased the size of the viral reservoir, it cannot delete infected cell and completely block ongoing viral replication. To reduce the existing viral pool, shorten the treatment cycle, and achieve cure, not only early treatment is needed, but also a combination of multiple strategies and long-term treatment are needed.
Tissue viral reservoirs are complex due to the diversity of organ/tissue architecture and cell composition. Some tissues have character of immunological and pharmacological sanctuary and can protect virus against drug or being cleared by the immune system. 28 Lymphoid tissues (LNs, spleen, and mucosa of gastrointestinal tract and genital tract) were rich in HIV/SIV target cells, and most studies on viral reservoir were mostly focused on them. The lymphoid tissues are highly enriched with provirus, CA-vRNA, and 2-LTR DNA (Fig. 4, 5, and 6). Thus, they are not only active site of SIV replication, with high viral transcription and persistent infection, but also the main reservoir of HIV/SIV in treatment-naive patients or monkeys. 29 Although short-term treatment in chronic infected individuals does not reduce total SIV DNA load in LNs, 15 long-term suppression therapies can reduce the total SIV DNA load in LNs, spleen, and gut. 30 However, viral reservoir persisted, and long-term treatment have little effect on total SIV DNA and CA-vRNA load in brain and adipose tissue. 31 –33 Unlike treatment that started at the chronic infection stage, when a stable viral reservoir has been established, short-term treatment in the early stages of infection inhibited viral transmission and reduced the size of the viral reservoir without inducing a virus-specific T and B cell response. (Fig. 4–6). However, the effect of short-term cART on total SIV DNA, CA-vRNA, and 2-LTR DNA level varied by tissue type, leading to a change in the size distribution of anatomical viral reservoir, characterized by a decreased contribution of lymph tissue and an increased contribution of nonlymph tissue, especially lung and liver (Fig. 7). These results may be related to the size of viral reservoir established before cART, 7 drug concentration in anatomical sites, immune response, and decay of the viral reservoir. 3 Although early cART reduced the size of viral reservoir in LNs, it remained the dominant viral reservoir, with highest viral persistence and current infection. These results may be due to the presence of a high level of viral reservoir in the lymph nodes before cART, but high levels of viral production and current infection should not be ignored (Fig. 7). In addition, lower drug concentrations in the lymph nodes have been reported. 34 Except for LNs, CA-vRNA levels were high in both lymph tissue (e.g., spleen, rectum, and duodenum et al.) and nonlymph tissue (including lung, liver, and adrenal), suggesting the presence of a highly active/persistent viral reservoir in these tissues. Higher level of viral reservoir, which was seeded before treatment may also interpret part of the findings. Consistent with persistent transcription of SIV, there was a higher level of 2-LTR DNA in LNs, spleen, duodenum, adrenal, thyroid, prostate, penis, epididymis, and bladder, suggesting higher current infection in these tissues. Although we cannot speculate whether the infection is an abortive infection or not, these results suggest to some extent that reverse transcription is not completely impeded at these anatomical sites. In addition, stable rank of total SIV DNA level in LNs or increased rank of total SIV DNA level in nonlymph tissue (adrenal, thyroid, prostate, penis, epididymis, bladder, cerebrum, testicle) may imply de novo infection in them, which may be caused by low drug concentration in these tissues. 28 Meanwhile, different decay of viral reservoir caused by cell death and immune response may also contribute to more or less. 35
In conclusion, although early treatment reduced the viral reservoir in SIV-infected monkeys, our study highlights the persistence of the virus in anatomical sites, particularly in nonlymph tissues, after short-term treatment. It suggested that the contribution of nonlymph tissue cannot be ignored in the development of strategies for HIV treatment.
Footnotes
Acknowledgments
The authors thank all staff and students from the laboratory of Molecular Immunopharmacology of Kunming Institute of Zoology for their help in macaque anatomy.
Authors’ Contributions
Conceptualization: R.-R.T and Y.-T.Z; Formal analysis: R.-R.T. and T.L.; Investigation: R.-R.T., T.L., M.-X.Z., T.-Z.S., and H.-Y.Z.; Resources: H.-Y. Z.; Writing––Original Draft: R.-R.T and T.L.; Writing––Review & Editing: all authors; Funding acquisition: R.-R.T. and Y.-T.Z.
Author Disclosure Statement
All authors report no potential conflicts.
Funding Information
This work was supported in part by grants from the National Key R & D Program of China (2023YFC2306700, 2021YFC2301703, 2018ZX10301101-002, 2017ZX10202102-001-005), the National Natural Science Foundation of China (32070181, U23A20473, 82202508, 82350710801, 81601808), and Yunnan Applicative and Basic Research Program (202101AT070282, 2016FB134, 202101AU070138).