
Abstract
Human T cell lymphotropic virus type 1 (HTLV-1) is associated with adult T cell leukemia/lymphoma (ATLL), a fetal malignant infection. Recently, HTLV-1 new asymptomatic carriers (ACs) have frequently been reported among blood donors. Reaching the profound concept of HTLV-1-associated molecular pathogenesis could result in finding novel therapeutic strategies. The current study aimed to determine leukemia-related signaling regulation in ATLL. Thirty participants were evaluated in 3 groups, including 10 ATLL patients, 10 ACs, and 10 normal controls. Blood samples were isolated without any chemotherapy history from ATLL patients. Also, blood samples were recovered from ACs and normal individuals. White blood cells isolation was done on the collected blood samples. After this, RNA was extracted from the prepared samples and used for the cDNA synthesis. TAX and HTLV-1 basic leucine zipper factor as viral genes and cellular genes, including MKP-1, EVI-1, JNK-1, FOXO-1, AKT-1, DEPTOR, MTOR, and JUN, were investigated using real-time PCR. The mean age of ATLL patients was 53.2 ± 7.32 years, and 9 (90%) were male. The EVI-1 and FOXO-1 expression levels were significantly associated with ATLL patients compared with the internal control. However, the significant differences in expression of other genes in the remaining groups were not seen. Discovering viral and cellular signaling pathways that regulate HTLV-1 transformation is essential. A novel therapeutic strategy for ATLL-regulating cellular signaling pathways in vivo could be considered. Therefore, clinical trials using activators and inhibitors of related cellular signaling pathways for cell therapy of ATLL are recommended. It is recommended that more investigation be conducted on FOXO-1 and EVI-1 to target these genes and reveal the molecular pathogenesis of ATLL.
Introduction
Concerning the current International Committee on Taxonomy of Viruses (ICVT) taxonomy, the human T cell lymphotropic virus type 1 (HTLV-1) belongs to Retroviridae, Orthoretrovirinae, and Deltaretroviruses. HTLV-1 is a retrovirus belonging to the Deltaretrovirus genus, Oncovirinae subfamily. 1,2 In addition to Iran, 3 HTLV-1 is distributed in Africa (8), South America, the Caribbean, Japan, Australia, and Melanesia, and the world widely infects about 20 million people. 1,4 Mother-to-child breastfeeding, sexual route, and infected blood transfusion are HTLV-1 transmission routes. 5
T cell contamination by HTLV-1 results in immune damage and adult T cell leukemia development. 6 Also, HTLV-1-associated myelopathy–tropical spastic paraparesis is another neurological disorder because of virus infection. 7 Less clinical manifestations of HTLV-1 include uveitis, bronchiectasis, dermatitis, and scabies. Furthermore, HTLV-1 asymptomatic carriers (ACs) are a strongly circulating pool of virus without a manifest symptom. 7
The HTLV-1 TAX (p40) protein has a fundamental role in virus biology. 1 In adult T cell leukemia/lymphoma (ATLL), TAX can prevent DNA repair and induce genomic instability and aneuploidy and causes cell cycle arrest and senescence. 8 –11 HTLV-1 basic leucine zipper factor (HBZ), a transcript from the minus strand of the provirus, downregulates HTLV-1 transcription. 1 By affecting telomerase, HBZ increases replicative immortality in the infected clone. 12 HBZ promotes cellular proliferation. However, it contributes to HTLV-1 persistence and has antiapoptosis and antiautophagy roles in the infected cells. 1 The ATLL’s major clinical features are flower cells and mean multilobulated nuclei of leukemic cells. In ATLL, activated T cells have CD3+, CD4+, CD8–, and CD25+ profile markers. 13 The genetic background is significantly associated with ATLL development. Therefore, investigations on cellular genes are essential. 13 Besides the viral genes, dysregulation of cell cycle factors in HTLV-1-infected cases affects HTLV-1-associated ATLL progression. CREB, NF-κB, AKT, PI3K/Akt, AP1, Cyclidins, BCL-2, FOS, and SMADs are well-known cellular signaling pathways in response to TAX and HBZ proteins, which have critical functions in ATLL cell transformation and immune system failure. 14 In ATLL pathogenesis, TAX oncoprotein interacts with the expression of different signaling pathways genes, cell cycle regulators, transcription factors, and apoptotic proteins. 15 Another HTLV-1 oncoprotein, HBZ, has interactions with AP1 and JunD. This interaction activates the tumor suppressor p53 and promotes T cell proliferation. 15 HTLV-1-infected T cell transformation during ATLL is moderated by numerous viral and cellular factors in a long clinical latency period. 16 –18 The current study evaluated leukemia-related signaling pathways among ATLL, ACs, and normal groups.
Materials and Methods
Population and sample collection
This study was enrolled in the Tehran University of Medical Sciences-affiliated hospitals in Tehran, Iran (ethical code: IR.TUMS.SPH.REC.1400.208). An experienced infectious disease specialist and an oncologist confirmed the diagnosis of HTLV-1-associated ATLL based on clinical and laboratory manifestations. However, the HTLV-1 screening test Enzyme-Linked Immunosorbent Assay (ELISA) and molecular detection Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR) were done. The ACs and healthy groups recovered from the Iranian Blood Transfusion Center by passing serological and molecular detection protocols for HTLV-1 infection. Participants with a history of chemotherapy were excluded because of the bias risk affecting the expression profile of ATLL, ACs, and the healthy included groups. All group members were matched by age range and gender. After written informed consent, 6 ml blood samples were collected from eligible participants using Ethylene Diamine Tetraacetic Acid (EDTA) tubes. White blood cells were isolated from the blood samples using RBC lysis buffer and density gradient centrifugation.
RNA extraction and qRT-PCR
Using the RNA Extraction RNJia Kit (ROJE), total RNA was extracted from whole blood based on the manufacturer’s procedures and recent protocols. 19 The RNA reverse transcription and cDNA synthesis occurred using random hexamer and oligo dT primers guided by the RT-ROSET kit (ROJE). Gene expressions of viral HBZ and TAX and cellular genes including MKP-1, EVI-1, JNK-1, FOXO-1, AKT-1, DEPTOR, MTOR, and JUN were assessed using SYBR Green RT-qPCR (Yekta Tajhiz Azma) and the Qiagen Corbett machine to perform relative standard curve real-time PCR on the cDNA samples. The housekeeping Ribosomal Protein Lateral Stalk Subunit P0 (RRPLP0) internal control gene was targeted for data normalization. Table 1 lists the designed primers used for the current study.
Designed Primers Used for Determining the Cellular and Viral Gene Expression Profile
HBZ, HTLV-1 basic leucine zipper factor.
Statistical analysis
The Graph-Pad Prism statistical software (version 8.0.2.263) conducted all the statistical analyses using the Kruskal–Wallis test. Statistically, the p value < .05 was considered significant to define differences in study groups.
Results
Population data
Totally, 30 participants were included in the current study; 27 (90%) were male, and 3 (10%) were female. The median age of the three groups was 53.46 ± 7.8 years. In total, 10 participants were imported into a separate group.
Viral HBZ and TAX genes expression profile
By using the Mann–Whitney test, a significant difference was observed in TAX (<0.0001) and HBZ (<0.0001) in ACs and ATLL groups. The mean (±SD) expression of TAX (ACs: 0.20 ± 0.35; ATLL: 1.08 ± 0.09) and HBZ (ACs: 0.11 ± 0.03; ATLL: 1.17 ± 0.59) is listed in Table 2 and Figure 1.
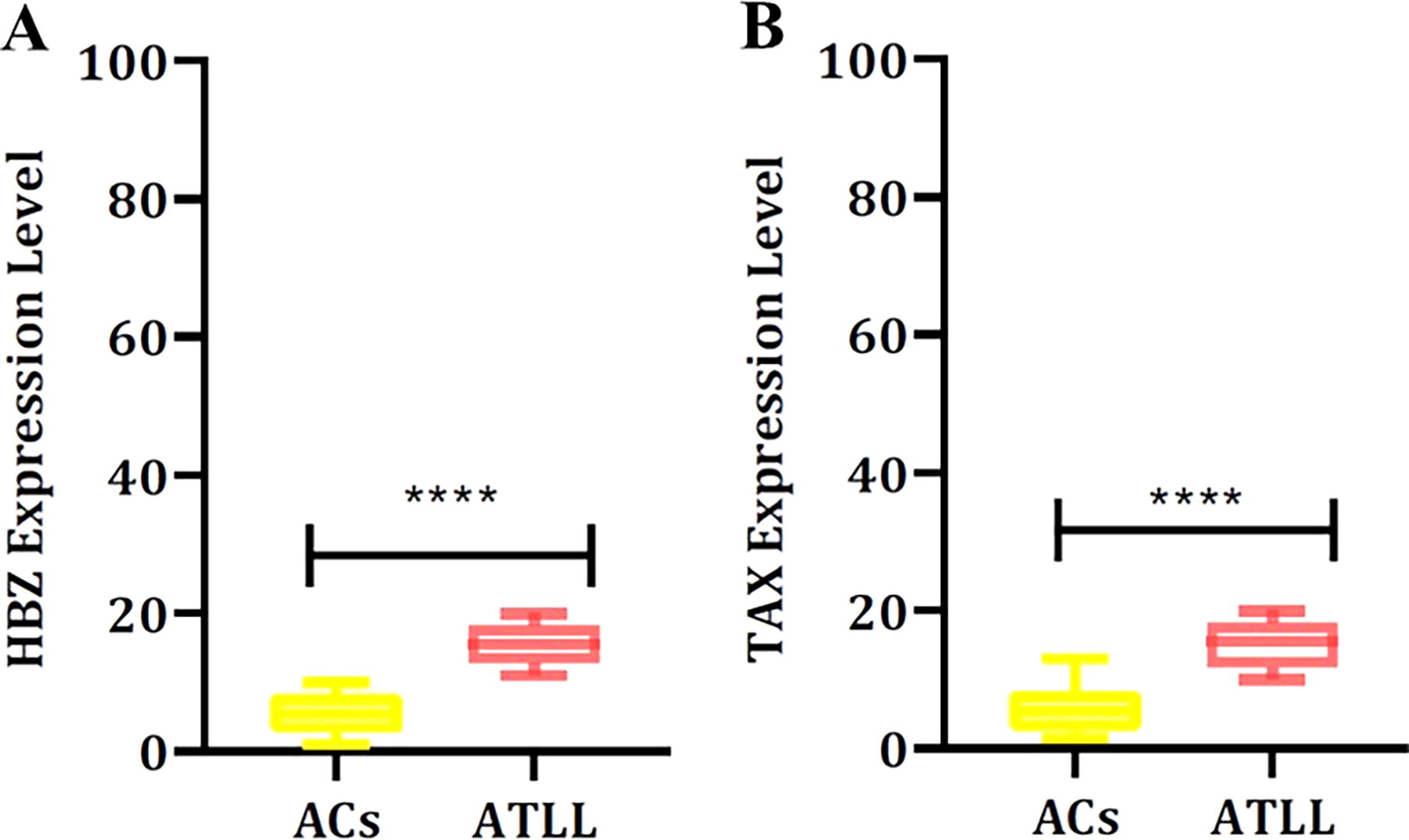
The expression levels of HBZ
Viral Expression Level Among ACs and ATLL Groups Using Mann–Whitney Test
Bold data represents statistically significant upregulation of HBZ and TAX among the ATLL group.
HBZ, HTLV-1 basic leucine zipper factor
Cellular genes expression profile
A significant difference was seen using the Kruskal–Wallis test in EVI-1 (<0.0001) and FOXO-1 (0.0002) expression levels among groups. Analysis revealed the mean (±SD) expression of EVI-1 (normal: 3.04 ± 3.03; ACs: 0.49 ± 0.49; ATLL: 0.08 ± 0.11) and FOXO-1 (normal: 1.27 ± 0.83; ACs: 0.36 ± 0.42; ATLL: 0.07 ± 0.12) (Table 3 and Fig. 2). Dunn’s multiple comparisons tests showed significant differences in EVI-1 and FOXO-1 between normal and ATLL groups with p value < .0001 and p value = .000, respectively (Table 4 and Fig. 2). In addition, the correlation evaluation was done to define the association between different genes. The results revealed a significant correlation between JNK-1 and EVI-1 (p = .007), JNK-1 and FOXO-1 (p = .001), JNK-1 and MTOR (p = .001), JNK-1 and JUN (.002), AKT-1 and DEPTOR (p = .01), AKT1 and MTOR (p = .08), MTOR and DEPTOR (.06), JUN and DEPTOR (.07), and MTOR and JUN (.001) in the ATLL group. Exceptional associations were followed between HBZ and DEPTOR (p = .06), JNK-1 and JUN (0.09), and DEPTOR and JUN (0.054) in the ACs group.
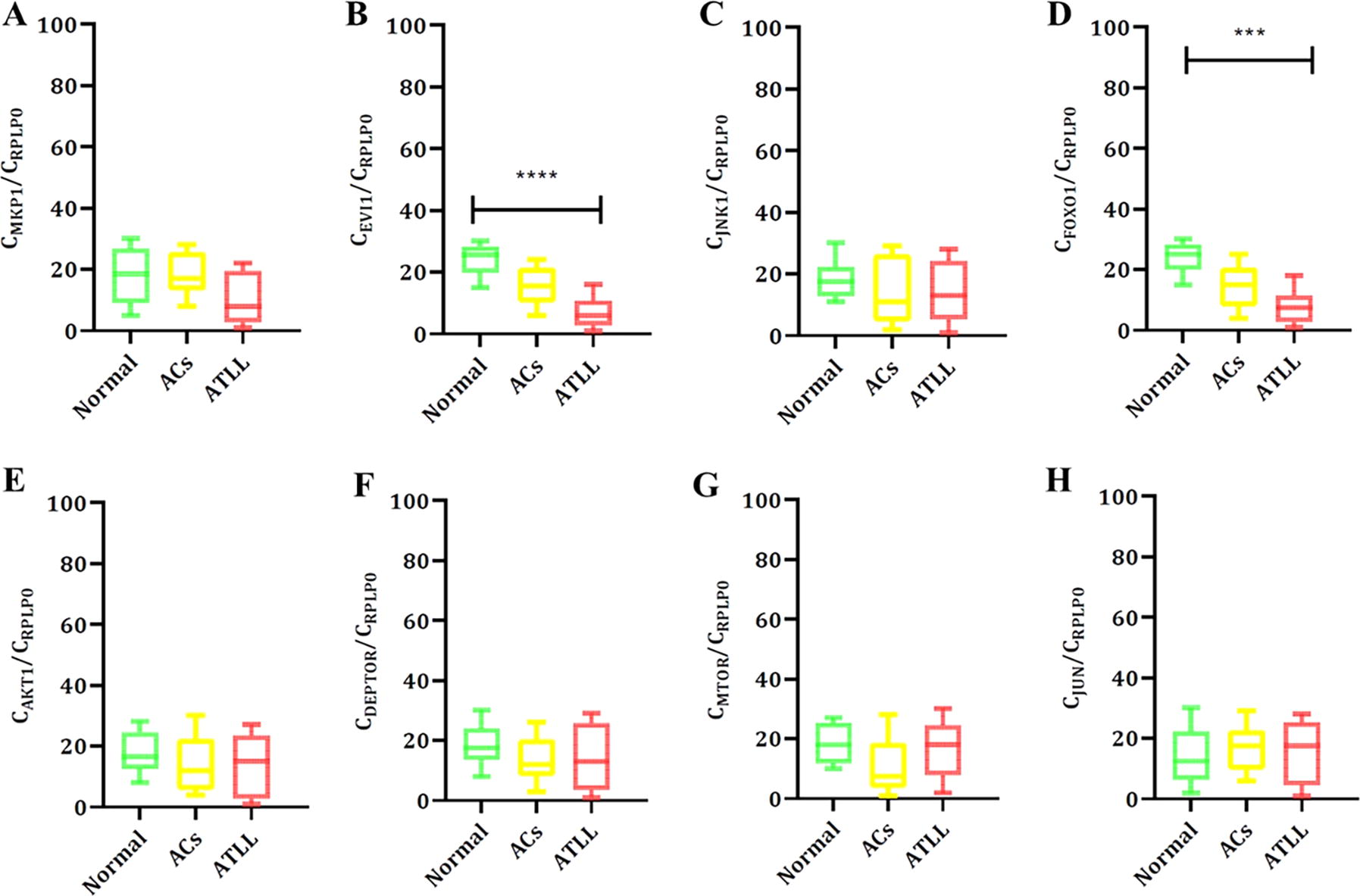
The expression levels of MKP-1
Cellular Expression Level Among Normal and ACs and ATLL Groups Using Kruskal–Wallis Test
Bold data represents statistically significant downregulation of EVI-1 and FOXO-1 among ATLL groups compared to Normal and ACS groups.
Cellular Expression Level Among Normal and ACs and ATLL Groups Using Dunn’s Multiple Comparisons Test
Bold data represents statistically significant downregulation of EVI-1 and FOXO-1 among ATLL groups compared to the Normal group.
p ≤ 0.001.
p ≤ 0.0001.
Discussion
The HTLV-1 as Retroviridae family is enveloped virus and has a single-stranded RNA genome. 20 The HTLV-1 is firstly Retrovirus which was discovered. 21 HTLV-1 as a retrovirus is categorized into different genetic subtypes (a–g) according to the diversity of its long terminal repeat region. 22 Each of the HTLV and HIV retroviruses has similar transmission routes and tropisms for T cells. The coinfection rate of these two viruses is probably high because HIV patients are not going through screening for HTLV infection in clinics. 23 Also, studies have reported that HTLV/HIV-1 coinfection can be evaluated as a new vaccine development strategy for HV-1. 24 Finally, patients infected with HIV-1/HTLV-1 have a higher mortality rate. 25 Currently, studies have been enrolled on HTLV-1-associated ATLL molecular pathogenesis, and different cellular factors have been analyzed. Therefore, the findings and revealed data will be discussed and compared together. The mitogen-activated protein kinases (MAPK) have distinct family members, including c-Jun NH2-terminal kinase (JNK) kinase family 1, whose signaling is deregulated in malignant hematopoiesis. 26 Stress, apoptosis, and growth factors stimulate the JNK signaling pathway. 26,27 The JNK pathway and its role in leukemia pathogenesis are not well understood. However, there are possible relationships between JNK kinases and acute leukemia development, indicating that targeting JNK and its substrate, c-Jun, may be a novel therapeutic strategy in resistance in acute myeloid leukemia. 28 Furthermore, studies investigated the JNK activation pathway’s role in ATLL pathogenesis. 29 The TAX oncoprotein transactivates related genes to HTLV-1 pathophysiology. MAPK kinases are functional in TAX-mediated NF-KB activation. 26 Since the JNK signaling pathway affects ATLL pathogenesis, the JNK kinase could be a target for therapeutic procedures. 26
ATLL development depends on the dysregulation of cellular gene transcription by HTLV-1. Studies showed that HTLV-1 TAX influences activating protein-1 (AP-1) members, including c-Jun. 30 In addition, HTLV-1 HBZ suppresses c-Jun by disrupting DNA-binding function and proteasome-dependent pathway, resulting in cell proliferation dysregulation. 30,31 As we know, TAX has a transactivator role and transforms cells by disrupting various cellular signaling pathways, including SRF, CREB, NF-kB, and AP-1 32,33 However, TAX dysregulates cellular checkpoints and blocks DNA repair, which is the first stage in T cell transformation. 32,33
DUSP-1, inhibiting the MAP kinase signaling pathway, is downregulated in different cancer types and is known as MKP-1. 34 A recent study indicated that dual specificity phosphatase (DUSP-1) could modify drug treatment response. Evidence suggests that the DUSP-1 expression defines the apoptotic threshold in cancer cells. 35 Growth factor and oncogenic kinase signaling affect the expression of DUSP1 in tumor cells modeling leukemias. 36 Tsukasaki1 et al. evaluated DUSP-1 as a progression-associated gene in ATLL and reported that expression levels of DUSP-1 were greater in PHA-stimulated T cells than in resting T cells. 37
The interplay between DNA damage cellular genes and HBZ regulates the mTOR signaling pathway. In addition, the growth arrest process increases dephosphorylation and suppresses mTOR expression. 38 Furthermore, TAX upregulates mTOR in T cells, which is part of HTLV-1-infected T cell growth. 39
DEPTOR in another cellular associating membrane and signaling proteins with two isoforms negatively regulates mTORC1 and mTORC2. 40,41 DEPTOR is functional in leukemia and in T-ALL patients. 42 NOTCH1, in cooperation with AKT, increases leukemogenesis, 43 although, during ATLL leukemogenesis, through transcriptional control of DEPTOR and AKT regulation, NOTCH1 upregulates DEPTOR. 44 Studies showed that control of DEPTOR expression and PI3K-AKT has an essential role in the pathogenesis of different blood cancer types and is a specific therapeutic target in cancer. 41
Antiapoptosis and cell survival are outcomes of the AKT activation pathways. Studies report that in HTLV-1-transformed cells, AKT is activated through TAX and results in cell survival. 45 Furthermore, AKT’s survival role in HTLV-1-transformed cells is related to Bcl-xL expression. 45 However, the AKT inhibition decreases the proapoptotic phosphorylated Bad protein level. 46 AKT signaling pathway regulates caspases and p53 apoptosis in HTLV-1-transformed cells. Since AKT inhibitors prevent NF-kB and p53 activation, AKT could be considered a therapeutic target in HTLV-1-associated ATLL. 46
Our study data revealed a significant relationship between EVI-1 and FOXO-1 cellular genes among the ATLL group, which is shown in Figure 3. EVI-1 is an aggressive oncogene associated with human leukemias. Dysregulation of EVI-1 suppresses the TGF-β pathway, disrupts cellular differentiation, and increases cell proliferation. 47 The levels of EVI-1 transcripts have a significant relationship and are also associated with cancer prognosis. 48 Forkhead box (FOX) proteins are transcriptional factors that significantly function in cell proliferation, differentiation, apoptosis, metabolism, and cellular migration. 49 FOX dysregulation occurs in genetic and malignant diseases such as leukemia. FOXM1 and FOXO family members are notable mediators in leukemia development and pathogenesis. Deregulation of FOXM1 and FOXOs induces leukemogenesis. As a routine, FOXM1 is upregulated, whereas the expression of FOXOs is downregulated because of degradation in leukemia. FOXOs are tumor repressors in leukemia. 50 However, active FOXOs sustain leukemia stem cells and contribute to leukemogenesis. They are valuable diagnostic and prognostic markers in leukemia for clinical applications. 50 The HTLV-1 TAX downregulates FOXO4, related to T cell transformation and ATLL development. 51 Tax, through PIK3CA-AKT1 signaling pathway activation, inhibits FOXO4 in HTLV-1-transformed cells. 51 However, stem cell therapies are a prospective field for ATLL treatment. 52 Also, Bazarbachi et al. showed that different allogeneic stem-cell transplantations of ATLL can influence long-term survival among 40% of patients after more than 5 years. 53
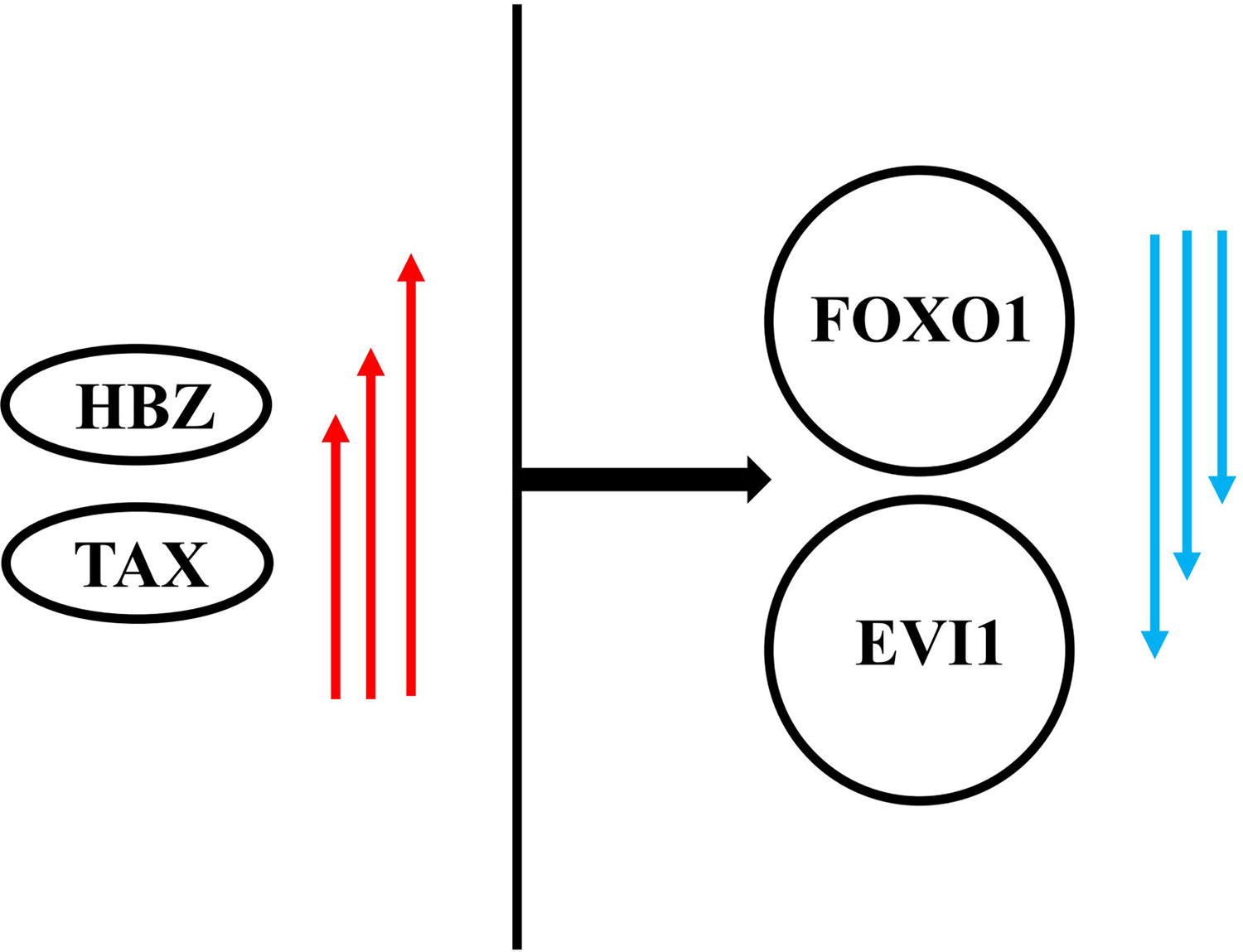
The pattern obtained from the interaction of the signaling pathways of viral and cellular genes among ATLL patients. ATLL, adult T cell leukemia/lymphoma.
Conclusion
The exact cellular factors that regulate Tax and HBZ remain the main question. However, besides TAX and HBZ, many cellular proteins are involved in the HTLV-1 transformation. Since infected T cells in ATLL disease are resistant to drugs, discovering viral and cellular signaling pathways that regulate HTLV-1 transformation through in vivo studies is essential. A novel therapeutic strategy based on regulating the upstream and downstream of oncogenes and tumor repressors could be considered for ATLL treatment in the clinic. Therefore, combining traditional chemotherapy and cellular signaling activator/inhibitor for treating HTLV-1 ATLL clinical trials is recommended.
Footnotes
Acknowledgment
The authors thank the Vice Chancellor for Research, Tehran University of Medical Sciences for supporting the study.
Confirmation Statement
EACH-listed author confirms that their research is supported by an institution that is primarily involved in education or research.
Authors’ Contributions
S.A.S., S.H.M., and M.E.N.: Concept and study design. S.A.S. and SH.M.: Laboratory investigation. S.A.S. and S.H.M.: Data analysis. S.A.S., S.H.M., R.O.R., S.E.H.M., M.H.M., S.E.S., M.F.H., and N.I.D.: Article writing and revised. All authors have read and approved the article’s content and confirmed the accuracy or integrity of any part of the work.
Availability of Data and Materials
All data generated and analyzed during the current study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
This research followed the principles of the Declaration of Helsinki. The Ethics Committee of Tehran University of Medical Sciences approved this study (ethical code: IR.TUMS.SPH.REC.1400.208). Informed consent was obtained from all subjects and/or their legal guardian(s).
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
Tehran University of Medical Sciences funded and supported the study (Grant number: