
Abstract
Abstract
Tobacco smoking is a risk factor for the development of atherosclerosis. One strategy to reduce the harmful side effects of tobacco smoking is the development of next-generation tobacco and nicotine products. A crucial step in the development of atherosclerosis is the activation and adhesion of monocytes. Remarkably, little is known about the direct effects of cigarette smoking on monocytes. In this study, we developed methods to determine the detrimental effects of aqueous cigarette smoke extract (CSEaq) from 3R4F reference cigarettes on human monocytes in vitro. First, we tested the cytotoxic effects of CSEaq on human peripheral blood monocyte cell line derived from an acute monocytic leukemia patient (THP-1) and primary monocytes. Treatment of THP-1 cells with CSEaq reduced cell viability at dosages ≥40% in a dose-dependent manner. Cell viability of primary monocytes was reduced by CSEaq at dosages ≥10%. CSEaq activated the antioxidative NRF2 system and its target gene NAD(P)H dehydrogenase, quinone 1 in a dose-dependent manner in both cell types. Expression of target gene HMOX1 was increased only in THP-1 cells. In addition, we showed an increased expression of NADPH oxidase subunit NOX2 in response to CSEaq in THP-1 cells, but not in primary monocytes. Furthermore, CSEaq strongly induced adhesion molecule intercellular adhesion molecule 1 in THP-1 cells, but not in primary monocytes. Pro-inflammatory markers CCL2 and CD31 showed a strong induction in response to CSEaq in THP-1 cells, but they remained unchanged in primary monocytes. In conclusion, we have developed in vitro test systems allowing the evaluation of cytotoxic, oxidative, and pro-inflammatory effects of CSEaq on monocytes. These test systems can be considered potential model systems for assessment of next-generation tobacco and nicotine products. Although THP-1 cells were more susceptible to changes of gene expression, primary monocytes showed a stronger reduction of cell viability in response to CSEaq.
Introduction
T
One strategy for reducing the harmful side effects of tobacco smoking is the development of next-generation tobacco and nicotine products. Recently, new products for inhalation of nicotine (e-cigarettes) were launched. 13 Additional innovative and potentially less risky tobacco products are based on heating rather than burning tobacco (also known as “heat-not-burn” products), are more closely aligned with consumer preferences, and are supposed to have less potential to provoke disease compared with conventional cigarette smoking.14–18 Nothing is yet known about the contribution of next-generation tobacco and nicotine products to viability, oxidative stress, and pro-inflammatory phenotype of monocytes.
The aim of this study was to develop methods to determine the effects of smoking on monocytes in vitro. To date, no reference product for next-generation tobacco and nicotine products has been established. We chose CSEaq from 3R4F reference cigarettes (University of Kentucky, Lexington, KY) to develop standardized methods.18–21 The commercially available monocyte cell line THP-1 (ATCC TIB-202) was used as a model for mimicking the function and regulation of monocytes in the vasculature in this project. 22 In addition, primary human monocytes were used as a second model. 23 We assessed the cytotoxic effects of CSEaq by determination of its viability with two different methods (CellTiter-Glo® Luminescent Cell Viability Assay and neutral red uptake assay).
Reactive oxygen species (ROS) play an important role in the development of endothelial dysfunction and atherosclerosis.24,25 Under physiologic conditions, several cellular systems are responsible for maintaining the balance of ROS levels. 26 The transcription factor Nuclear factor, erythroid 2 like 2 (NRF2/NFE2L2) is the main mediator of cellular adaptation to oxidative stress, and it is directly regulated by ROS and cigarette smoke in vascular and blood cells. 27 Activation of the NRF2 antioxidant defense system is an essential step in the response of cells to survive oxidative stress situations by binding to antioxidant response elements in the 5′regulatory regions of detoxifying and antioxidant target genes, such as heme oxygenase 1 (HMOX1) or NAD(P)H dehydrogenase, quinone 1 (NQO1) and modulating their transcription.27–29 NADPH oxidases are a major source of ROS and play a central role in this context. 26 The main source and marker of oxidative stress in monocytes is the superoxide anion-producing NADPH oxidase 2 (NOX2/CYBB). 30 The production of superoxide anions (O2−) by NOX2 contributes to the development of endothelial dysfunction and atherosclerosis, whereas the generation of H2O2 by NADPH oxidase 4 has been shown to play an important role in maintaining endothelial function and in reducing the formation of atherosclerotic lesions.25,31,32
Endothelial inflammation is accompanied by a higher expression of adhesion molecules. 33 Classical molecular markers of vascular inflammation are intercellular adhesion molecule 1 (ICAM-1), vascular cell adhesion molecule 1 (VCAM-1), selectin E (SELE), and platelet and endothelial cell adhesion molecule 1 (CD31/PECAM-1). 34 In the progression of atherosclerosis, pro‐atherosclerotic and pro‐inflammatory cytokines are responsible for the induction of local inflammation and the recruitment of immune cells. 2 C-C motif chemokine ligand 2 (CCL2/MCP-1) is the monocyte-specific surface marker that accelerates atherosclerosis and shows an increased expression in atherosclerotic lesions.35,36
Therefore, the determination of viability, the expression of the NRF2 antioxidant defense system, NADPH oxidase 2, adhesion molecules, and pro-inflammatory genes in response to conventional or next-generation tobacco and nicotine products will provide important information/critical insights into the impact of these products on monocytes, possibly leading to novel insights into potentially harmful effects.
Materials and Methods
Cell culture
Human monocytic THP-1 cells (ATCC# TIB-202) were provided by the Department of Internal Medicine and Cardiology, TU Dresden, Germany. Cells were cultivated in RPMI-1640 (Thermo Fisher Scientific, Waltham MA), which was supplemented with 10% fetal calf serum (Biochrom, Berlin, Germany), 100,000 U/L penicillin (Thermo Fisher Scientific, Waltham MA), and 100 mg/L streptomycin (Thermo Fisher Scientific, Waltham MA). Every 2–3 days, THP-1 cells were centrifuged at 250 × g for 10 min and resuspended in 8 mL culture medium. One milliliter of the cell suspension was transferred into a fresh culture flask, and 11 mL of culture media was added for further cultivation. THP-1 cells were cultured for a maximum period of 8 weeks.37,38 Primary human monocytes were isolated from buffy coats from healthy donors (Institut für Transfusionsmedizin Dresden, Germany) by a two-step density gradient centrifugation and cultured as previously described. 23 Primary monocytes were used for no longer than 96 hours after the isolation procedure. All experiments were conducted in a humidified environment with 5% CO2 at 37°C.
CSEaq production and stimulation
Overall, 3R4F reference cigarettes were conditioned at 22°C and 60% relative humidity for 48 hours and smoked on an RM20H smoking machine (Borgwaldt-KC, Hamburg, Germany) under ISO standard puffing conditions (1 puff/min; 2 s/puff >35/2/60, vents open; ISO 3308:2012), with the exception of only eight puffs taken per cigarette instead of smoking the cigarette to a specified butt length. CSEaq was generated by bubbling the smoke from a single cigarette through a glass impinger containing 20 mL of sterile Phenol Red-free Medium 199 (without supplements; Thermo Fisher Scientific, Waltham, MA). After preparation, the extract was directly frozen at −80°C and was sent to our department on dry ice. The extract was not filtered before use. It contains all water-soluble components from the whole smoke aerosol, consisting of a particulate and a gas phase. This in vitro approach is used for exposing cardiovascular cells to cigarette smoke constituents and is suggested to mimic the in vivo situation in the vascular system of a smoker. After the arrival, fresh CSEaq was thawed just before use and used only once. The remaining extract from an already thawed vial was disposed. Each CSEaq was used for a maximum period of 7 days after extraction. Before treatment with CSEaq, cells were cultivated in Phenol Red-free Medium 199; next, they were supplemented with 10% fetal calf serum, 100,000 U/L penicillin, and 100 mg/L streptomycin for 16 hours. Stimulation with CSEaq was performed in a concentration range from 10% to 88.3% for as long as 48 hours within the same medium. Each sample was accompanied by a control from the same cell passage or cell preparation and was incubated for the same period without stimulation of CSEaq (time-matched controls).
Analysis of cell viability of human monocytes
The amount of adenosine triphosphate (ATP) is an indicator of metabolically active cells and is directly proportional to the number of living cells. 39 By quantification of the amount of ATP using CellTiter-Glo Luminescent Cell Viability Assay (Promega, Mannheim, Germany), the number of viable cells in culture can be determined. Monocytes (25,000 per one 96 well) were seeded and cultured for 48 hours (medium was changed after 24 hours). Cells were stimulated with CSEaq in a concentration range from 10% to 88.3% for as long as 48 hours. Cell viability assay was performed, and the emitted luminescence signal was measured as relative light units by using a luminometer. As a second approach, neutral red uptake assay (#N2889, Sigma-Aldrich, Munich, Germany) for the estimation of cell viability/cytotoxicity was performed. Neutral red is incorporated into lysosomes of living cells, whereas dying cells have a reduced ability to incorporate neutral red. Thus, the loss of neutral red uptake corresponds to the loss of cell viability. Neutral red stock solution (3.3 g/L) was used as a 1:50 dilution in Phenol Red-free Medium 199, which was supplemented with 10% fetal calf serum, 100,000 U/L penicillin, and 100 mg/L streptomycin. After incubation for 2 hours, cells were washed with 37°C pre-warmed phosphate-buffered saline (PBS; 10.0 mM phosphate, 2.68 mM KCL, 140.0 mM NaCl, pH 7.45; Thermo Fisher Scientific, Waltham MA) and the assay was stopped by using 1:1 1% glacial acetic acid solution in 50% EtOH. The incorporated dye was solubilized in the solution. After 10 min of incubation at 22°C, absorption was measured at 540 nm in reference to a blank.
Real-time polymerase chain reaction
Monocytes (700,000 per one well) were seeded in a 24 well-plate. The medium was changed after 24 hours. Cells were cultivated for an additional 24 hours and stimulated with CSEaq in fresh medium for a further 48 hours. Total RNA was isolated by using the High Pure RNA Isolation Kit (Roche Diagnostics, Mannheim, Germany). Reverse transcription of mRNA into cDNA was performed with SuperScript II Reverse Transcriptase according to the manufacturer's instructions (Thermo Fisher Scientific, Waltham MA). Total RNA (500 ng) and random hexamer primers were used. Quantification was performed by a real-time polymerase chain reaction (PCR) (7500 Fast Real-Time PCR System, Thermo Fisher Scientific, Waltham MA) by using GoTaq qPCR Master Mix (Promega, Mannheim, Germany) with specific primers (Sigma-Aldrich, Munich, Germany; for primer sequences, see Table 1). POLR2A was used as a reference gene for cDNA content normalization. Amplification started with an initial denaturation step at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 s, and specific annealing/extension for each gene at 60°C for 60 s. Melt‐curve analysis was performed after every run to ensure a single amplified product in each reaction. Analysis of the raw data was performed with the 7500 Software Version 2.06 (Applied Biosystems by Life Technologies, Darmstadt, Germany). Data were evaluated by using a mathematical model of relative expression ratio in real-time PCR under constant reference gene expression. 40
Statistical analysis
Data are shown as mean ± standard deviation. Statistical analysis was performed by One-Way ANOVA followed by Holm–Sidak post hoc test when appropriate (SigmaPlot 13.0, Systat Software, Inc., San Jose, CA). A value of p < 0.05 was considered statistically significant.
Results
Cytotoxic effects of CSEaq leading to decreased cell viability of human monocytes—development of a test system for analysis of cell viability
An analysis of the cytotoxic capacity of conventional or next-generation tobacco and nicotine products is necessary to estimate the potentially harmful effects of their consumption. To study the cytotoxic effects of CSEaq, THP-1 and primary monocytes were exposed to increasing dosages of CSEaq in a concentration range from 10% to 88.3% for 24–48 hours. First, we used the CellTiter-Glo Luminescent Cell Viability Assay to quantify the degree of cytotoxicity caused by CSEaq. Treatment of THP-1 monocytes with CSEaq reduced cell viability at dosages ≥40% after 24 and 48 hours in a dose-dependent manner. No statistically significant differences could be detected between the incubation for 24 hours and that for 48 hours (Fig. 1A). Measurements using neutral red uptake assay for estimation of cell viability/cytotoxicity showed a similar threshold of 40% CSEaq, which was slightly delayed (48 hours). At dosages ≥60%, no differences between 24 and 48 hours of incubation could be detected (Fig. 1B). In summary, both cell viability assays showed comparable results in THP-1 cells. Treatment of primary monocytes with CSEaq reduced cell viability stronger. This was shown by the CellTiter-Glo Luminescent Cell Viability Assay. Dosages of 10% or higher reduced monocyte cell viability after 24 and 48 hours in a dose-dependent manner. Except for dosages of 40%–60%, no statistically significant differences could be detected between the incubation for 24 hours and that for 48 hours (Fig. 1C).
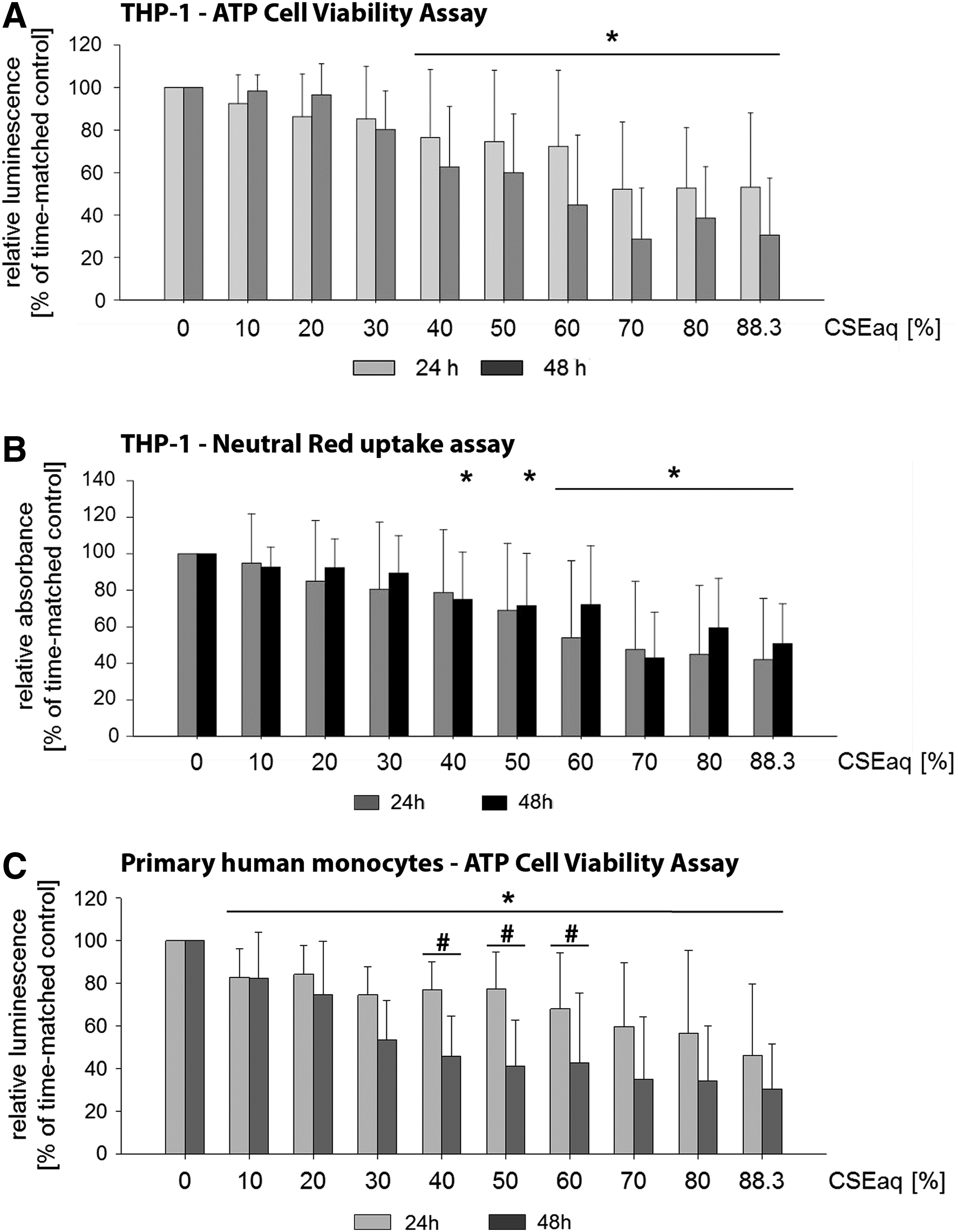
Cell viability of human monocytes after exposure to cigarette smoke extract (CSEaq). Monocytes were cultivated as suspension cultures. Subsequently, 25.000 cells were stimulated with CSEaq in a concentration range from 10% to 88.3% for 24–48 hours.
Activation of NRF2 antioxidant defense system and NOX2 by CSEaq in human monocytes—development of a model to test NRF2 activation and oxidative stress
The transcription factor NRF2 is a main mediator of cellular adaptation to oxidative stress. Activation of the NRF2 antioxidant defense system is a critical response of cells to survive oxidative stress situations. Therefore, the analysis of activation of the NRF2 system can give insights into the induction of oxidative stress by conventional or next-generation tobacco and nicotine products. In this study, we stimulated THP-1 and primary monocytes with 10%-50% of CSEaq for 24 hours. Afterward, mRNA expression of NRF2 and its target genes HMOX1 and NQO1 was quantified. Exposure with CSEaq activated the NRF2 system in both cell types in a dose-dependent manner. This was shown by strongly increased expression of NRF2 target gene NQO1 (Fig. 2C). The expression of HMOX1 was increased only in THP-1 cells (Fig. 2B), whereas transcription factor NRF2 itself remained unregulated in both cell types (Fig. 2A).

mRNA expression of NRF2 and the target genes HMOX1 and NQO1 after exposure to CSEaq in human monocytes. Primary and THP-1 monocytes were cultivated as suspension culture. Afterward, the cells were stimulated with CSEaq in a concentration range from 10% to 50% for 24 hours. Subsequently, mRNA expression was measured by real-time PCR. POLR2A was used as a reference gene.
The generation of ROS plays an important role in the development of atherosclerosis. A main source and marker of oxidative stress in monocytes is the superoxide anion-producing NOX2. Determination of the expression of NOX2 gives important insights into the cellular potential to induce oxidative stress. After stimulation with increasing dosages of CSEaq, NOX2 was upregulated in THP-1 monocytes. In primary monocytes, NOX2 expression remained at a stable level (Fig. 3).

mRNA expression of oxidative stress marker NADPH oxidase 2 (NOX2) after exposure to CSEaq in human monocytes. Primary and THP-1 monocytes were cultivated as suspension culture. Afterward, cells were dose-dependent stimulated with CSEaq in a concentration range from 10% to 50% for 24 hours. Subsequently, NOX2 mRNA expression was measured by real-time PCR. POLR2A was used as a reference gene. Data are shown as x-fold of time-matched controls ± SD. Statistics: One-Way ANOVA, *p < 0.05 compared with static time-matched controls, n ≥ 6.
CSEaq evokes pro‐inflammatory state of human monocytes—a model system to test the susceptibility of monocytes to atherosclerotic risk factors
Adhesion of monocytes to the endothelium reflects an inflammatory state. In the progression of atherosclerosis, pro‐atherosclerotic and pro‐inflammatory cytokines induce local inflammation and recruit immune cells. Therefore, we measured the expression of pro-inflammatory genes and adhesion molecules in response to CSEaq. CSEaq strongly induced adhesion molecule ICAM-1 in THP-1 cells. In primary monocytes, ICAM-1 expression was decreased at higher concentrations of CSEaq (Fig. 4A). Interestingly, VCAM-1 was not detectable in THP-1 cells. In primary monocytes, VCAM-1 showed a very low expression (Ct-values above 32 ± 2). Stimulation with CSEaq downregulated these levels further (data not shown). This expression pattern was confirmed by Western blot (data not shown). SELE was not expressed in both cell types. In contrast, pro-atherosclerotic and pro-inflammatory marker CCL2 (MCP-1, Fig. 4B) and CD31 (PECAM-1) showed a strong induction in response to CSEaq in THP-1 cells, but they remained unchanged in primary monocytes (Fig. 4C).

mRNA expression of adhesion molecules and cytokines after exposure to CSEaq in human monocytes. Primary and THP-1 monocytes were cultivated as suspension culture. Afterward, the cells were stimulated with CSEaq in a concentration range from 10% to 50% for 24 hours. Subsequently, mRNA expression was measured by real-time PCR. POLR2A was used as a reference gene.
Discussion
Cigarette smoking is an important risk factor of endothelial dysfunction and atherosclerosis.1–3 The underlying molecular mechanisms are not well understood. One important in vitro model to study smoking-related cellular changes is the stimulation of cells with CSEaq. 20 So far, little is known about the direct effects of cigarette smoking on the gene expression profile of monocytes. Nothing is known about the contribution of probably less harmful next-generation tobacco and nicotine products to the inflammatory phenotype of monocytes. In this study, we developed methods to determine the detrimental effects of CSEaq from 3R4F reference cigarettes on human monocytes in vitro. 3R4F CSEaq is a widely accepted standard, and it has been used in various research projects analyzing the impact of CSEaq.18–21,41 The monocyte cell line THP-1 and primary human monocytes isolated from buffy coats are well-accepted models for mimicking the function and regulation of monocytes in the vasculature.22,23 In this study, we combined both approaches to develop methods to investigate the effects of next-generation tobacco and nicotine products on human monocytes in comparative analyses.
Cytotoxic effects of tobacco smoking harmfully affect vascular endothelial cells. 42 CSEaq can decrease cell viability and induce a pro-inflammatory phenotype of endothelial cells.43,44 Determination of monocyte viability after stimulation with different tobacco or nicotine products could be of great value. First, we established two test systems for analyzing cell viability (Fig. 1): an ATP-based viability assay (CellTiter-Glo Luminescent Cell Viability Assay) (Fig. 1A) and a neutral red uptake assay (Fig. 1B). Both assays revealed the cytotoxic effects of CSEaq at dosages ≥40%, leading to decreased cell viability in a dose-dependent manner (Fig. 1A, B). Measurements using the neutral red uptake assay showed a delay of 24 hours before reaching the threshold (Fig. 1B). This observation can be explained by the principle of each assay. The CellTiter-Glo Luminescent Cell Viability Assay is mainly based on an enzymatic conversion of ATP, which correlates with the amount of metabolically active and viable cells. The neutral red assay is a colorimetric assay measuring the uptake of the dye by functional lysosomes. In response to specific stimuli, slight differences in the sensitivity of MTT assays and neutral red uptake assays have been shown and discussed. 45
In general, before choosing a viability assay, it is important to test more than one assay with different cell types, incubation times, and concentrations of the test substance. After conducting extensive preliminary tests also with endothelial cells, different CSEaq dosages, and various time points (unpublished data), we recommend the use of ATP cell viability assays for the investigation of the effects of next-generation tobacco and nicotine products. Treatment of primary monocytes with CSEaq reduced cell viability stronger (Fig. 1C). Already, lower dosages of ≥10% reduced primary monocyte cell viability in a dose-dependent manner (CellTiter-Glo Luminescent Cell Viability Assay). Differences in the response intensity of THP-1 and primary cells to atherosclerotic stimuli have been previously described.22,46 Similar dosages of CSEaq were used in recent studies investigating a pro-inflammatory phenotype, autophagic cell death, or apoptosis of endothelial cells.43,47,48 None of these previous studies analyzed the impact of CSEaq on monocytes.
In addition, we have established the stimulation of monocytes by CSEaq as an in vitro model to test the activation of the NRF2 antioxidant defense system and the induction of oxidative stress. Exposure of monocytes with CSEaq activated the NRF2 system in both cell types in a dose-dependent manner. This was shown by the strongly increased expression of the NRF2 target gene NQO1 (Fig. 2C). The increased expression of NQO1 has been shown in human bronchial epithelial cells after stimulation with cigarette smoke condensate from commercial and research cigarettes for 18 h earlier. 49 Interestingly, expression of HMOX1 was increased only in THP-1 cells (Fig. 2B). Even though THP-1 cells have become one of the most frequently used cell lines to investigate the function and regulation of monocytes in pathophysiological changes of the cardiovascular system, 22 differences between THP-1 and primary monocytes were recorded. Especially, changes in the global gene expression pattern have been the subject of intense investigation. 22 After stimulation with PMA (phorbol-12-myristate-13-acetate), 75 genes were regulated in THP-1 cells, 104 genes were regulated in primary monocytes, and only 17 of these genes were altered in both cell types. 50
While comparing both monocytic cell types, THP-1 cells have some advantages compared with human primary monocytes. The homogeneous genetic background of THP-1 cells minimizes the variability, which facilitates reproducible results. THP-1 cells have a high growth rate and can be cultured for several months or stored in liquid nitrogen without losing their morphological and functional properties. 22 In contrast, the availability of primary monocytes is limited. Their isolation has to be performed as soon as possible after blood donation. They can be contaminated with other blood components, and monocytes comprise only 3%–9% of all leukocytes. 37 Since our aim was to establish an easily reproducible, reliable, and simplified test system, we recommend the use of THP-1 cells for screening assays.
Finally, mRNA expression of the transcription factor NRF2 itself remained unregulated in both cell types (Fig. 2A). Recently, in a blood–brain barrier endothelial cell line exposed to 3R4F CSEaq, mRNA and protein expression of NRF2 target genes NQO1 and HMOX1 was induced. mRNA expression of the transcription factor NRF2 itself was not changed, but the translocation of the NRF2 protein into the nucleus was enhanced. 51 Our previous studies have shown a translocation of NRF2 into the nucleus in response to high laminar flow. 52 These data further support an important role of the regulation of NRF2 activity by translocation into the nucleus.
The generation of ROS plays an important role in the development of atherosclerosis.24,25 Particularly, the production of superoxide (O2−) by NOX2 contributes to the development of endothelial dysfunction and atherosclerosis.31,32 For this reason, we analyzed the effects of CSEaq on NOX2. NOX2 is a main source and marker of oxidative stress in monocytes. 30 In THP-1 monocytes, an induction of oxidative stress could be shown by an increased mRNA expression of NOX2 in response to CSEaq (Fig. 3). This is in agreement with a study showing increased superoxide production by NOX2 after stimulation with 3R4F CSEaq and detection of ROS by 2′,7′dichlorofluorescein diacetate (DCFH-DA) in the murine macrophage cell line RAW264.7. 53 In primary monocytes, NOX2 remained stable again (Fig. 3), showing a higher susceptibility of THP-1 cells to CSEaq.
Smoking induces a pro-inflammatory phenotype that is accompanied by adhesion of monocytes to endothelial cells. This is a crucial step in vascular inflammation and atherosclerosis.5–7,9,54 The indicated pro-inflammatory phenotype can be detected by an increased expression of adhesion molecules ICAM-1, VCAM-1, SELE, and CD31 (PECAM-1) of endothelial cells.33,34 CCL2 (MCP-1) is a monocyte-specific surface marker that induces local inflammation, recruits immune cells, accelerates atherosclerosis, and shows an increased expression in atherosclerotic lesions.35,36 We measured the mRNA expression of pro-inflammatory genes and adhesion molecules. CSEaq strongly induced adhesion molecule ICAM-1 in THP-1 cells at dosages ≥50% (Fig. 4A). This is in agreement with previous studies using endothelial cells.55,56 In addition, Adams et al. were able to show an increased expression of ICAM-1 and the adhesion of monocytes to endothelial cells in response to the serum of smokers. 57 An induction of ICAM1 expression in endothelial cells after treatment with supernatants from 3R4F CSEaq-stimulated MM6 monocytes was shown by Poussin et al. 41 This indicates an inflammatory profile of human monocytes that were exposed acutely to CSEaq. In primary monocytes, ICAM-1 expression was decreased at higher concentrations of CSEaq (≥30%) (Fig. 4A). Interestingly, VCAM-1 was not detectable in THP-1 cells. In primary monocytes, VCAM-1 showed a very low expression. SELE was not expressed in both cell types. In contrast, pro-atherosclerotic and pro-inflammatory markers CCL2 (Fig. 4B) and CD31 (Fig. 4C) showed a strong induction in response to CSEaq in THP-1 cells, but they remained unchanged in primary monocytes. Consistently, CCL2 was increased in human endothelial cells by extracts of smokeless tobacco. 58 CCL2 is a central mediator of smoking-induced chronic obstructive pulmonary disease. 59 An activation of CCL2 might be involved in the CSEaq-mediated monocyte adhesion to endothelial cells. 60 Furthermore, the smoking status of patients can correlate with the adhesion of monocytes to endothelial cells. 61
Another important step in the progression of atherosclerosis is the transmigration of leukocytes through the endothelium. 2 Recently, enhanced MCP-1 expression in freshly isolated human monocytes after stimulation with cigarette smoke condensate was shown and seems to be important for the migration of human monocytes. 62 Exposure of human uterine microvascular endothelial cells to cigarette smoke conditioned media leads to alterations in PECAM-1 distribution that may cause endothelial dysfunction and is in accordance with our findings in THP-1 monocytes. 63
The reason for the differences in ICAM-1, CCL2, and CD31 expression in primary monocytes and THP-1 cells (Fig. 4) is not clear. THP-1 cells resemble primary monocytes in many morphological and functional properties as well as in gene expression.64–66 However, a few differences have been reported. Sharif et al. compared the gene expression of several genes in LPS-treated THP-1 cells with primary monocytes from two separate donors and showed a good agreement in the tendency of change (induction or repression) of most genes; however, some differences were observed in the primary monocytes isolated from the different donors. 67 We could detect similar effects in our dataset of primary monocytes where a different tendency of expression between the donors was observed for CCL2 and PECAM-1. Our donors (n = 16) of primary monocytes are anonymous, and we do not know their genetic backgrounds. This might lead to genetic heterogeneity and can influence the basal gene expression. In the case of ICAM-1, CCL2, and PECAM-1, we observed a higher basal gene expression in primary cells compared with THP-1 cells (data not shown). In addition, the expression of extracellular receptors mediating the response to components of CSEaq and the activation of signal transduction pathways could be different between both monocytic cell types, resulting in a higher susceptibility of THP-1 cells to CSEaq. Finally, primary monocytes might have a higher probability to differentiate into macrophages, which could result in a different response compared with primary monocytes and THP-1 cells. 68
In summary, this study analyzed the cytotoxic, oxidative, and pro-inflammatory effects on monocytic THP-1 cells and primary monocytes. Simulation with CSEaq leads to decreased cell viability and changes in the expression of atherosclerosis-related genes of monocytes. Our data support the use of THP-1 cells as a model system to test next-generation tobacco products.
Conclusions
In this study, we developed methods to determine the detrimental effects of CSEaq from 3R4F reference cigarettes on human monocytes in vitro. We showed that the cytotoxic effects of CSEaq decreased cell viability in an in vitro test system. Furthermore, we established the activation of the NRF2 antioxidant defense system by CSEaq in monocytes as a model for the cellular response against oxidative stress. Activation of pro-inflammatory genes and adhesion molecules could be shown and used as a model system to test the susceptibility of monocytes to atherosclerosis. Finally, we showed an increased expression of NOX2 in response to CSEaq, which can be considered an oxidative stress test system. These test systems can be considered potential model systems for the assessment of next-generation tobacco and nicotine products.
Footnotes
Acknowledgments
The authors thank Kathy Eisenhofer for critically reading this article.
Author Disclosure Statement
This work was supported by a research grant from Research & Development, British American Tobacco, Southampton, the United Kingdom. The authors would like to thank Katherine Hewitt and Dr. Natalia Cockcroft from British American Tobacco, Research & Development, Southampton, the United Kingdom, for having kindly provided CSEaq for this study on a weekly basis.