
Abstract
Introduction:
As a chemotherapeutic agent, doxorubicin (DOX) is limited because of systemic toxicities. Repeated treatment with DOX reduces the therapeutic response and increases cellular resistance to the drug. Carotenoids are highly efficient in preventing and treating cancer. Coenzyme Q10 (CoQ10) or ubiquinone has a dynamic role in electron transfer within the mitochondrial oxidative respiratory chain. This study aimed to evaluate the synergistic activity of different concentrations of carotenoid lutein (CL) and CoQ10 combined with doxorubicin on the MCF7 cell line.
Materials and Methods:
Evaluation of cancer cell growth inhibition by the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay, free radical scavenging of reactive oxygen species (ROS) test with DCFH reagent, measurement of membrane lipid damage by lipid peroxidation test with thiobarbituric acid (TBA) reagent, and measurement of superoxide dismutase (SOD) and glutathione peroxidase (GPx) enzymatic antioxidants were performed using the ZellBio kit.
Results and Discussion:
Compared with the doxorubicin group, CoQ10 and lutein at high concentrations significantly reduced cancer cell growth by 65% and 60.19%, respectively (p < 0.01). A significant decrease in GPx and SOD enzyme activities was observed with 10 μg/mL CoQ10 (by 66.52% and 63.55%) and lutein (by 64.50% and 66.20%), (p < 0.001 and p < 0.01), respectively. CoQ10 and lutein at 10 μg/mL increased the level of malondialdehyde by 76.66% and 76.80%, respectively (MDA; p < 0.001). CoQ10 and lutein treatment groups at 10 μg/mL significantly increased intracellular ROS by 58.32% and 57.49%, respectively (p < 0.001). Cancer cells' metabolism is higher than normal cells, associated with increased oxidative stress. The increase in oxidative stress is correlated with a decrease in cell viability, which indicates the role of oxidative damage in the cancer cell line.
Conclusions:
This study suggests that using supplements such as CL and CoQ10 may help reduce cancer cell growth as adjunctive therapy with doxorubicin. As shown in this study, the use of high-dose supplements can cause oxidative stress and inhibit the growth of cancer cells by inducing ROS and reducing antioxidants and it benefits the therapeutic process by inhibiting the growth of cancer cells.
Introduction
Breast cancer (BC) is one of the most prevalent types of cancer and a significant reason for death in women globally after lung cancer.1,2 Multiple predisposing factors for BC have been reported till now. Environmental and behavioral factors are more critical in determining the prevalence of cancer among populations, and dietary deficiencies play a significant role in the pathogenesis, progression, and metastasis of BC.1,3 However, many treatment strategies have been identified to treat clinical BC, including cytotoxic agents. Elimination of cancer cells by cytotoxic agents can be performed through various routes, including the interstitial DNA mechanism, impaired DNA repair by inhibition of topoisomerase II, increased intracellular H2O2 and reactive oxygen species (ROS), and injury formation in cell membranes and proteins.4–6
Doxorubicin (DOX) is an antitumor drug from the anthracycline family and is one of the most effective drugs in the treatment of BC.7,8 Several different pathways have been approved for the function of doxorubicin, such as DNA intercalation 9 ; membrane binding7,10; reductive activation 11 ; inhibition of topoisomerase II activity; and increased levels of free radicals in cells. 6
As a chemotherapeutic agent, DOX is limited because of systemic toxicity and cardiac toxicity. Repeated treatment with doxorubicin reduces the therapeutic response and increases cellular resistance to the drug. 12 Resistance to antitumor drugs is often the main reason for treatment failure in cancer patients; therefore, various dietary supplements have been considered by researchers. 13
Due to fewer side effects, using natural compounds is a new therapeutic strategy for treating various chronic diseases, including cancer. Vegetables with green leaves, fruits, and edible seaweed contain carotenoids.14,15
Carotenoids are highly efficient in preventing and treating cancer. They have also been shown to affect cancer cell death's biochemical and molecular events and their anticarcinogenic mechanisms are different. 16 It is thought that chemical and structural properties of carotenoids are involved in antioxidant defense mechanisms and cellular signaling. 17 Numerous studies have reported the various pathways through which carotenoids exert their anticancer effects, including antioxidant and pro-oxidant pathways; cell differentiation; antiproliferation, antiangiogenic, and anti-inflammatory properties; and immune modulation.18–20
Coenzyme Q10 (CoQ10) or ubiquinone is a lipophilic, endogenic, intrinsic, membrane-protective, and potent antioxidant with different biological activities such as immune boosting, radical scavenging, and DNA protection. 21 It has a dynamic role in electron transfer within the mitochondrial oxidative respiratory chain. 22
As a response to reductive oxidative stress in most pathological situations, it is supposed that amount of CoQ10 increases.23,24 Based on some evidence, there is a relationship between CoQ10 deficiency and different types of cancer. 25
CoQ10 improves the anticancer effects of doxorubicin and reduces this drug's cardiac toxicity through various mechanisms, including antitumor and antiangiogenic potential in cancer cells, reduction of ROS activity, and lipid peroxidation (LPO) in normal cells.26–28
This study aimed to evaluate the synergistic, antiproliferative, and anticancer activities of different concentrations of carotenoid lutein (CL) and CoQ10 combined with doxorubicin on the MCF7 cell line.
Materials and Methods
Chemicals
3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT), ethylene diaminetetraacetic acid (EDTA), dichloro-dihydro-fluorescein diacetate (DCFH-DA), thiobarbituric acid (TBA), and dithiobis-2-nitrobenzoic acid (DTNB) were purchased from the Sigma-Aldrich company. All chemicals used were of analytical grade.
Cell lines
The human BC cell line, MCF7, was purchased from the Iran Rangeland Institute in Tehran. Cell suspensions containing Dulbecco's modified Eagle's medium (DMEM), 1% pen-strep, and 10% fetal bovine serum were placed in a 75-cm3 cell culture flask and incubated at 37°C in a humid atmosphere of 5% CO2 to reach their logarithmic growth stage. When the primary culture became almost confluent, one cc of trypsin-EDTA was added to the flask and incubated for 5 minutes to completely separate the cells from the flask. Then, to evaluate the oxidative stress and cell survival tests, 100 μL of cell suspension was added to each well in 96-well plates, and the plates were incubated for 24 hours.
MTT assay
After 24 hours of incubation, the cell line was exposed to a single dose of doxorubicin (IC50 = 3.38 μM) 29 and each dose of 2, 4, 6, 8, and 10 μg/mL lutein and Q10 with incubation for 48 hours. Then, 10 μL of MTT solution was added to each well and incubated for 4 hours. After that the contents of each well were removed and washed with phosphate-buffered saline, then 30 μL of dimethyl sulfoxide (DMSO) was added to dissolve the formazan dye, and absorption was determined at max = 490 and 630 nm using the BioTek ELx800 microplate reader.
Activity of glutathione peroxidase
Glutathione peroxidase (GPx) reduces lipid hydroperoxides to the corresponding alcohols and reduces free hydrogen peroxide to water. Cell lipid compounds are sensitive to free radicals and react to produce lipid peroxide. GPx enzymes use glutathione to reduce peroxides in alcohol and prevent formation of free radicals. In fact, GPxs catalyze the reduction of hydrogen peroxide (oxygenated water) and a wide range of organic peroxides related to alcohol and water using cellular glutathione. 30
GPx activity was assessed using a ZellBio kit (Berlin, Germany). GPx activity is based on the glutathione reductase assay. The experiment's aim is to reduce the hydroperoxide molecule by GPx, which produces the oxidized form of glutathione.
Activity of superoxide dismutase
Superoxide dismutase (SOD) participates in the toxic radical dismutation produced during the oxidation of energy substrates to O2 and H2O2. In this method, xanthine and xanthine oxidase (XOD) are used to produce superoxide radicals that react with INT [phenyl tetrazolium chloride-5-(nitrophenyl-4)-3-(iodophenyl-4)-2] to form formazan red. It can be measured at 420 nm. In the presence of the SOD enzyme, superoxide radicals, it is converted to hydrogen peroxide and O2. Finally, the formation of formazan red color is prevented, and the activity of the SOD enzyme is determined by the degree of inhibition of this reaction. Briefly, the sample was first mixed with WST solution (containing xanthine and INT), then the SES solution containing XOD was added and incubated at 37°C for 20 minutes. Absorbance of the samples was then measured at 450 nm. Finally, enzymatic activity was calculated in units per mg of protein. One unit of SOD is the amount of enzyme that reduces the rate of reduction or reduction by the enzyme XOD by up to 50%. 30
LPO assessment
LPO was evaluated by determining the production rate of thiobarbituric acid-reactive substances (TBARS) and expressed as malondialdehyde (MDA) equivalents.
TBA reagent, 0.1 mL, and phosphoric acid, 0.1 mL, were added to the cell suspension and placed in a warm water bath (100°C) for 30 minutes and then placed on ice for 5 minutes. After cooling, 0.2 mL of n-butanol was added, shaken well, and then centrifuged at 3500 rpm for 10 minutes. Absorbance of the supernatant was measured at 532 nm using the BioTek ELx800 microplate reader. 31
ROS assessment
Intramitochondrial ROS was measured by the DCFH-DA reagent. Dichloro-dihydro-fluorescein diacetate (DCFH-DA) was used to check the amounts of ROS; 20 μL of DCFH-DA was added to each well and incubated for 15 minutes and absorbance was measured at a wavelength of 485–530 nm, using a fluorescence microplate reader [23].
Statistical analysis
Prism, ver.3, software was used to perform statistical tests. One-way analysis of variance, followed by Tukey's test, was used to determine the differences among groups (p < 0.05). SPSS, V21, software was used to evaluate and obtain the appropriate mathematical equation.
Results
Combinatorial therapy of CL and CoQ10 with DOX decreases MCF7 cell proliferation
As shown in Figure 1, treatment (48 hours) of MCF7 with different concentrations of lutein or CoQ10 combined with a single dose of doxorubicin reduced the growth of cancer cells. Treatment of cancer cells with a single dose of doxorubicin inhibited the growth of cancer cells by 51%. In addition, treatment of cells with a single dose of lutein and Q10 at a 10 μg/mL concentration inhibited cell growth by 14.31% and 13.12%, respectively (Table 1).

The effect of Q10 or CL in combination with DOX on cell viability in MCF7 cells. High doses of lutein and Q10 at about 15% and doxorubicin at a concentration of 3.38 μM show 51% growth inhibition. *p < 0.05, **p < 0.01, and ***p < 0.001 compared with the doxorubicin group. CL, carotenoid lutein; DOX, doxorubicin. Color images are available online.
Mean and Standard Deviation of the Effect of Lutein or Q10 in Combination with Doxorubicin
CL, carotenoid lutein; CoQ10, coenzyme Q10; DOX, doxorubicin; GPx, glutathione peroxidase; LPO, lipid peroxidation; MTT, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide; ROS, reactive oxygen species; SOD, superoxide dismutase.
Combination treatment of lutein with doxorubicin at concentrations of 2–10 μg/mL reduced the growth of cancer cells. The growth inhibition rate was 51.10% at the lowest concentration of lutein (2 μg/mL) and 60.19% at the highest concentration (10 μg/mL). Compared with the doxorubicin group, treatment with doxorubicin and lutein at concentrations of 8 and 10 μg/mL significantly reduced cancer cell growth (p < 0.01). In other words, the cell growth inhibition rate was 55.85% at a concentration of 8 μg/mL and 60.19% at a concentration of 10 μg/mL.
Exposure of BC cells to coenzyme CoQ10 and doxorubicin reduced growth by 52.58% at 2 μg/mL and 65% at 10 μg/mL. Compared with the doxorubicin group, treatment of MCF7 cells with doxorubicin and coenzyme CoQ10 at concentrations of 4 and 6 μg/mL (p < 0.05) and 8 and 10 μg/mL (p < 0.01) showed a significant difference. Inhibition of cell growth was 58.76% at a concentration of 6 μg/mL, 63.11% at a concentration of 8 μg/mL, and 65% at a concentration of 10 μg/mL.
Combinatorial therapy of CL and CoQ10 with DOX activity of GPx
Figure 2 shows the activity of the enzyme GPx. Treatment with different concentrations of both supplements, CL and CoQ10, combined with DOX, was done for 48 hours.
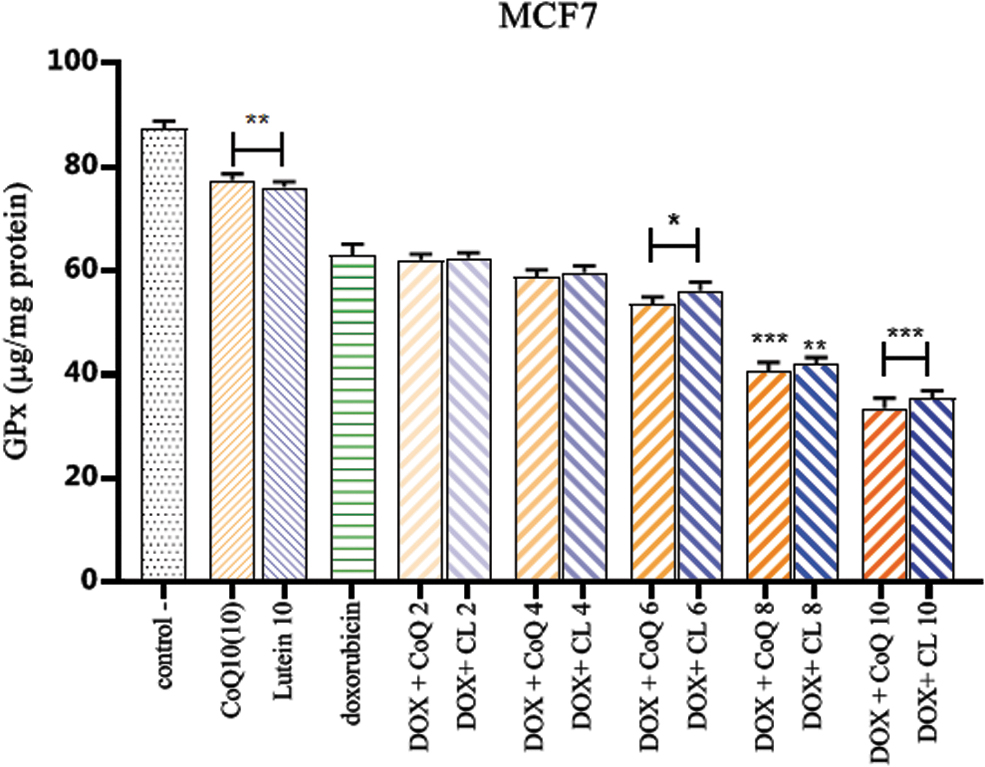
The effect of Q10 or CL combined with DOX GPx activity in MCF7 cells. The activity of the GPx enzyme with exposure to high doses of lutein and Q10 decreased by about 24%, and addition of doxorubicin at a concentration of 3.38 μM resulted in a 36.89% decrease in enzyme activity. *p < 0.05, **p < 0.01, and ***p < 0.001 compared with the doxorubicin group. GPx, glutathione peroxidase. Color images are available online.
As shown in the diagram, doxorubicin reduced the enzyme activity by 36.89%. In cells exposed to a single dose of lutein and Q10, the GPx activity decreased by 23.94% and 22.55%, respectively (Table 1).
Exposure of cells to combination therapy with lutein and doxorubicin at different concentrations further reduced the activity of the GPx enzyme by 37.59% at a concentration of 2 μg/mL and 65.50% at the highest concentration, and combination therapy with coenzyme CoQ10 inhibited GPx activity by 37.9% at the lowest concentration and 66.52% at the highest concentration.
Compared with the doxorubicin group, we observed significantly decreased activity of the enzyme with CoQ10 and lutein at a concentration of 6 μg/mL (p < 0.05), and the rates for lutein and CoQ10 at concentrations of 8 μg/mL (p < 0.01) and 10 μg/mL (p < 0.001) were significantly different.
Combinatorial therapy of CL and CoQ10 with DOX activity of SOD
The effect of carotenoid and CoQ10 treatment with the anticancer drug DOX on SOD activity of MCF7 is shown in Figure 3. The different concentrations of lutein and CoQ10 combined with DOX significantly depleted the SOD activity.
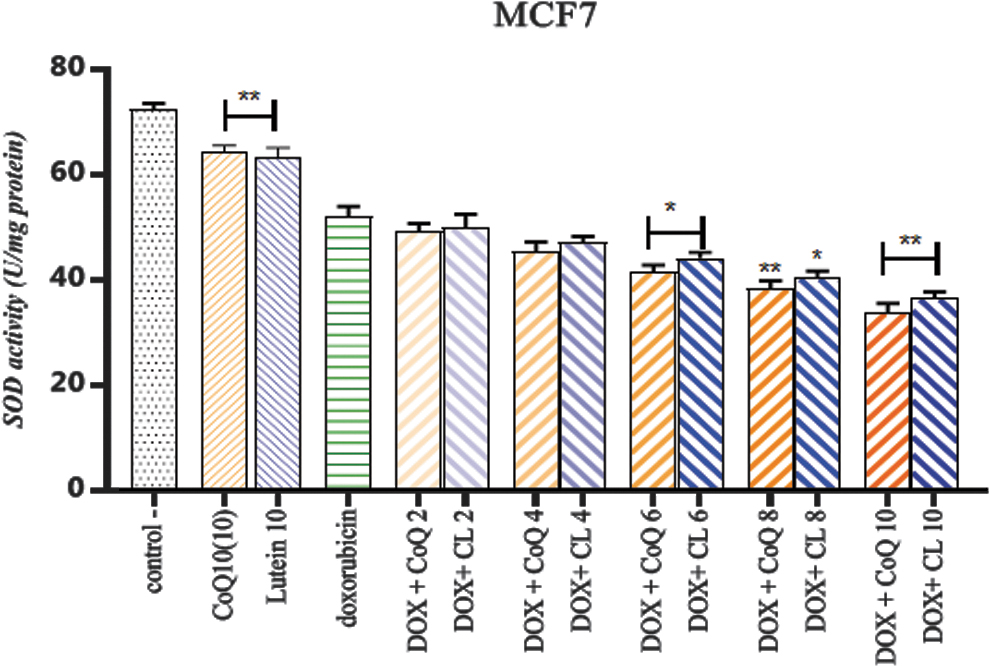
The effect of CL or CoQ10 combined with DOX on SOD activity in MCF7 cells. SOD activity decreased by about 37% in the presence of high doses of lutein and Q10 and doxorubicin at a concentration of 3.38 μM. *p < 0.05 and **p < 0.01 compared with the doxorubicin group. CoQ10, coenzyme Q10; SOD, superoxide dismutase. Color images are available online.
Treatment of cells with doxorubicin reduced SOD activity by 47.89%. The decrease in SOD activity in cells exposed to a single dose of lutein and Q10 was 36.6% and 35.8%, respectively (Table 1).
However, combination treatment with doxorubicin and lutein reduced this amount by 50.19%, 52.89%, 52.84%, 59.71%, and 63.55% at different concentrations of 2–10 μg/mL, respectively. A significant decrease was observed in the enzyme activity with lutein at concentrations of 6 and 8 μg/mL (p < 0.05) and 10 μg/mL (p < 0.01) compared with the doxorubicin group.
Exposure of MCF7 cells to CoQ10 and doxorubicin at the lowest concentration of 50.77% and the highest concentration of 66.20% resulted in a decrease in SOD activity. Compared with the doxorubicin group, SOD activity at a concentration of 6 μg/mL (p < 0.05) and concentrations of 8 and 10 μg/mL (p < 0.001) showed a significant difference.
Combinatorial therapy of CL and CoQ10 with DOX and evaluation of LPO
MDA was a final product of LPO, resulting from oxidative stress. It is used for the evaluation of oxidative stress. A significant increase was observed in MDA concentration in MCF7 cells treated with DOX/lutein and Q10 for 48 hours compared with the DOX group (Fig. 4).
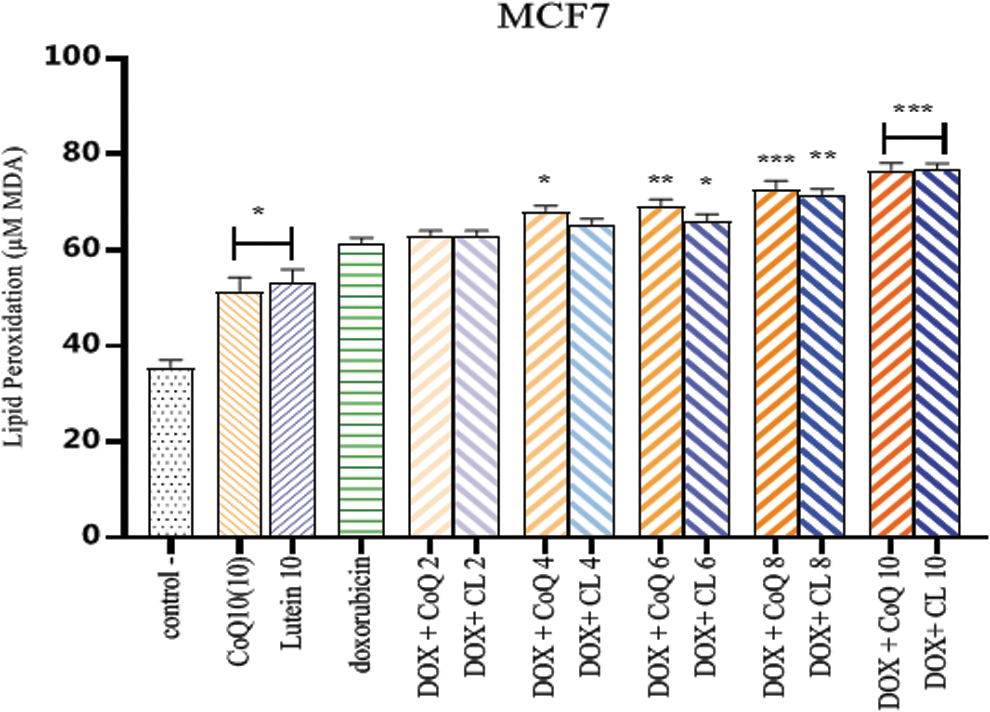
The effect of lutein or Q10 in combination with DOX on MDA levels in MCF7 cells. The amount of MDA produced in MCF7 cells was about 53% with high doses of lutein and Q10 and 61.47% in combination with doxorubicin at a concentration of 3.38 μM. *p < 0.05, **p < 0.01, and ***p < 0.001 compared with the doxorubicin group. MDA, malondialdehyde. Color images are available online.
Doxorubicin increased the MDA content of cancer cells by 61.47%. A single dose of lutein and Q10 at a concentration of 10 μg/mL induced MDA in dead cancer cells, 53.20% and 51.20%, respectively (Table 1).
Combination therapy with DOX+CL at concentrations of 2–10 μg/mL increased the LPO of cancer cells from 62.8% at 2 μg/mL to 76.80% at 10 μg/mL. Compared with the doxorubicin group, lutein showed a significant difference at concentrations of 6 μg/mL (p < 0.05), 8 μg/mL (p < 0.01), and 10 μg/mL (p < 0.001).
Cell exposure to CoQ10 and doxorubicin also increased LPO from 62.98% at 2 μg/mL to 76.66% at 10 μg/mL. Compared with the doxorubicin group, LPO levels at concentrations of 4 μg/mL (p < 0.05), 6 μg/mL (p < 0.01), and 8 and 10 μg/mL (p < 0.001) were significantly different.
Combinatorial therapy of CL and CoQ10 with DOX and evaluation of ROS
Intracellular ROS concentrations were assessed by changing the intensity of DCFH-DA fluorescence. As shown in Figure 5, the combined treatment of doxorubicin with lutein and Q10 increased intracellular ROS formation.

The effect of lutein or Q10 in combination with DOX on ROS levels in MCF7 cells. The amount of ROS produced in MCF7 cells was about 38% with high doses of lutein and Q10 and 42.84% in combination with doxorubicin at a concentration of 3.38 μM. *p < 0.05, **p < 0.01, and ***p < 0.001 compared with the doxorubicin group. ROS, reactive oxygen species. Color images are available online.
The amount of ROS formation in cancer cells in the presence of doxorubicin was 42.84%. However, treatment of cells with a single dose of lutein and Q10 at a concentration of 10 μg/mL induced ROS formation by 38.56% and 38.04%, respectively (Table 1).
Adding lutein at a concentration of 2 μg/mL with doxorubicin did not change the amount of ROS formation (42.89%), but with increasing concentration, the amount of ROS formation increased from 46.10% to 57.49%. The doxorubicin and lutein treatment groups at concentrations of 6 and 8 μg/mL and 10 μg/mL were significantly different from the doxorubicin group, (p < 0.01) and (p < 0.001), respectively.
With the addition of CoQ10 and doxorubicin, intracellular ROS increased from 43.53% at a concentration of 2 μg/mL to 58.32% at a concentration of 10 μg/mL. ROS levels at concentrations of 4 μg/mL (p < 0.05), 6 and 8 μg/mL (p < 0.01), and 10 μg/mL (p < 0.001) were statistically different from the doxorubicin group.
A mathematical model to conclude the synergistic effect of doxorubicin+CoQ10/CL
Based on data obtained from the MTT test and using SPSS software, a mathematical model was determined to explain the synergistic effect of doxorubicin and CoQ10/CL (Tables 2 and 3).
Statistical Parameters of Coenzyme Q10 and Carotenoid Lutein
Mathematical Model of Doxorubicin+Coenzyme Q10/Carotenoid Lutein
As shown in Figures 6 and 7, with increasing CoQ10 concentration (on the x-axis) of 1.76% per unit and increasing lutein concentration of 1.68% per unit, the survival rate of BC cells significantly reduced. In other words, for each increment on the x-axis (concentration) by a factor of b, a significant decrease in MCF7 cell growth is observed.

Mathematical model to conclude the synergistic effect of doxorubicin+CoQ10.

Mathematical model to conclude the synergistic effect of doxorubicin+CL.
Based on the mean comparison test, lutein from a concentration of 8 μg/mL onward and CoQ10 from a concentration of 4 μg/mL onward were significantly more effective than the use of doxorubicin alone. Doxorubicin added in the concentration can inhibit cell growth by 50%. Concomitant use of doxorubicin with two supplements at a 2 μg/mL concentration increased the effect of doxorubicin. However, CoQ10 from 4 μg/mL onward and lutein from 8 μg/mL onward have a synergistic effect. It is suggested that the concentration of these two substances could be increased to 50 μg in future studies.
Discussion
Combination therapy is one of the newest and most important strategies for treating chronic and complex diseases such as cancer. We use combination therapy to reduce drug resistance, side effects, and drug toxicity and achieve a synergistic effect to treat or prevent disease progression. We investigate the growth inhibitory effect of different concentrations of CL and CoQ10 and the potential biological effect of the combination of CL+DOX and CoQ10+DOX in BC cells. Due to reduction in the antitumor side effects and resistance of doxorubicin, combination therapies have been considered.
In general, metabolism of cancer cells is higher than normal cells and is associated with increased oxidative stress. In other words, the increase in oxidative stress is associated with a decrease in cell viability, which indicates the role of oxidative damage in the cancer cell line. 32
Taking supplements with anticancer drugs is very helpful in limiting side effects. The present study results showed that lutein and CoQ10 with doxorubicin reduced the growth of cancer cells compared with a single dose of doxorubicin (Fig. 1).
Combination therapies increase intracellular ROS, which are the most crucial cancer cell proliferation factors.33,34 In this regard, studies have revealed that the cytotoxic event mediated by ROS is a potential route of anticancer drug activity at the site of malignancy.35,36 By increasing ROS production in cancer cells, the ability of endogenous antioxidants decreases, which leads to cell death due to oxidative damage. 32
The combination of carotenoids with anticancer drugs such as tamoxifen and oxaliplatin created a synergistic effect in cancer cells. 37 Due to the oxidative status and amount of intracellular ROS, administration of doxorubicin and different concentrations of carotenoids increases cytotoxicity by increasing the level of intracellular ROS, followed by increased apoptosis and inhibition of topoisomerase II. 32 More than two decades of research have focused on elucidating carotenoids' anticancer activity in in vitro and in vivo models. These mechanisms include increased ROS and intracellular LPO, followed by an increase in apoptosis. 38 Exposure of MCF7 cells to lutein has also modulated epithelial-mesenchymal-transition expression, reduced NOTCH signaling, and inhibited cancer cell migration and metastasis. 39 The results of Gong et al.'s experiments show that catenoids have been implicated as anticancer agents in various tumors. Lutein plays a role in inducing apoptosis and stopping the cancer cell cycle. Their experiments showed that lutein-exposed triple negative breast cancer cells produced higher levels of ROS, which inhibited the growth of cancer cells, whereas breast epithelial cells did not produce ROS when exposed to lutein. Lutein induces ROS production in macrophage cells and mitochondrial-mediated ROS generation, which induces apoptosis in cancer cells. 40
Several studies have focused on improving the effects of CoQ10 on BC patients. Some of them report that CoQ10 deficiency increases the risk of BC.41,42 On the other hand, CoQ10 acts as a powerful antioxidant against MMP-2 activity along with the pro-oxidant capacity of the MCF7 cell in a dose-proportional manner.21,43 Therefore, the use of CoQ10 is recommended to prevent cancer or cancer progression. 1 A study showed that exposure of cells to high doses of CoQ10 induces oxidative stress because the enzyme can freely cross the cell membrane and affect the activity and expression of MMP. 23 Various studies have reported that exposure of prostate cancer cells to CoQ10 reduced the growth of cancer cells while having no adverse effect on noncancer cells. A clinical study reported that administration of CoQ10 in combination with temozolomide and radiation therapy increased the sensitivity of cancer cells' DNA to radiation, which in turn increased the effectiveness of the drug.44–46
In addition, supplemented CoQ10 has been shown to consist of a fraction of mitochondrial CoQ10. Besides, Q10 supplementation is part of the Q10 fraction in mitochondria. Two hypotheses have been proposed for the anticancer effects of CoQ10: first, oxidative stress increases due to exposure of cells to higher exogenous doses of Q10 because the enzyme is lipophilic and passes freely through the membrane, disrupting the signaling pathways of anion superoxidase; and second, this enzyme has pro-oxidant properties and can help maintain the cytoplasm and membranes' physiological oxidation potential. Accordingly, superoxide anion production as a second messenger in signaling systems is affected. 21
The results of a study suggest that inhibition of the growth of BC cells with CoQ10 therapy may be due to inhibition of the inflammatory factor, NF-κB, which is one of the important factors in the cancer process. 38 Another study confirmed that CoQ10 with anti-inflammatory effects inhibited the growth of cancer cells. 21
Reducing the expression and response of GPx to oxidative stress factors in cancer cells increases tumor sensitivity to the drug. By increasing the production and accumulation of ROS in cancer cells, the cells progress to apoptosis. 6 Furthermore, the other reason for cancer cells' resistance to anticancer drugs is the accumulation of MDA due to LPO.6,47 One theory is that increase in the level of LPO and oxidation proteins and the imbalance between oxidant and antioxidant enzymes increase oxidative stress factors and cytotoxic effects. 47
Hence, regulation of ROS for treatment of cancer is very significant. In this regard, our study results showed the effect of carotenoids and CoQ10 with DOX on increase of ROS-mediated cell death.
As was found from the results of the present study, exposure of cancer cells to doxorubicin with lutein or CoQ10 increased intracellular ROS and subsequently increased LPO. Previous studies confirm these findings (Figs. 3–5).
Conclusions
Based on the reasons mentioned above, we hypothesize that using two antioxidant supplements, in addition to balancing oxidant and antioxidant levels, and exerting cytotoxic effects on cancer cells may prevent drug resistance and contribute to the better and more profound effect of anticancer drugs. The present study results have shown that the combination therapy had better results and achievement in reducing cancer cell growth.
Previous studies and this study suggest that using supplements such as CL and CoQ10 may help reduce cancer cell growth as adjunctive therapy with doxorubicin. As shown in this study, the use of high-dose antioxidants can cause oxidative stress and inhibit the growth of cancer cells by inducing ROS, which in turn reduces in vivo antioxidants and benefits the therapeutic process by inhibiting the growth of cancer cells.
Further studies are suggested to evaluate possible side effects of CL and CoQ10 and the role of oxidative stress in regulation of gene expression by lutein and Q10 in synergism with DOX to understand its mechanisms of action.
Footnotes
Acknowledgment
The authors thank Dr. Sayyed Kamaleddin Allameh (Isfahan Research Institute for Agricultural Science and Natural Resources—Animal Science, Biotechnology Unit, Doctor of Philosophy) for his guidance and advice in the mathematical modeling section.
Author Disclosure Statement
All authors declare that they have no conflicts of interest.
Funding Information
This study was supported by a grant from the Research Council of Mazandaran University of Medical Sciences, IR.MAZUMS.REC.1398.1113.