
Abstract
Introduction:
This study aimed at evaluating the anticancer activity of selenium and zinc in human colorectal adenocarcinoma (HT-29) cell line exposed to sulfasalazine (SSZ).
Methods:
Lipid peroxidation through the thiobarbituric acid reagent, oxygen-free radicals with the dichloro-dihydro-fluorescein diacetate reagent, glutathione (GSH) reserves through the 5,5’-dithiobis-(2-nitrobenzoic acid) reagent, and mitochondrial activity with the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide reagent were evaluated in the HT-29 cell line. The activity of superoxide dismutase (SOD) and glutathione peroxidase (GPx) enzymes and the activity of caspase-3 were determined using an ELISA kit. Genetic toxicity was also assessed by Comet assay.
Results and Discussion:
Combination therapy of SSZ with zinc or selenium in HT-29 reduced their growth, GSH, and the activity of SOD and GPx enzyme and increased reactive oxygen species and lipid peroxidation and the activity of caspase-3. The amount of tail moment in the comet test in the presence of these substances has also increased, indicating damage to cancer cells' DNA. In general, the best effect of selenium and zinc at concentrations of 100 and 200 μM (p < 0.05) (p < 0.01) was observed.
Conclusion:
Due to zinc and selenium's mechanism of action in inhibiting cancer cells' growth, these two substances can be used as a supplement to improve the effect of SSZ in treating diseases (e.g., colorectal cancer) and inhibiting drug toxicity.
Introduction
Colorectal cancer (CRC) is one of the often diagnosed malignancies and is recognized as the third leading cause of cancer death for both sexes around the world.1,2 However, surgery and resection of the bowel in cancer's early stages is a recommended treatment path. Chemotherapy continues to be an essential treatment pathway for patients diagnosed at an advanced stage. Hence, most cancer patients eventually develop drug resistance despite their initial response to anticancer drugs. Therefore, it is necessary to develop strategies to prevent drug resistance and increase its clinical efficacy. 3 The researchers proposed various mechanisms that make anticancer drugs resistant, such as detoxification, inhibition of the apoptotic process, and increased DNA repair. 4
Sulfasalazine (SSZ) inhibits cysteine transport, selectively inhibiting cancer cell growth. SSZ stimulates cell death by decreasing glutathione (GSH) and increasing intracellular reactive oxygen species (ROS). Clinical studies in cancer patients treated with SSZ have shown reduced GSH levels in cancer cells. SSZ is an anti-inflammatory drug used to treat ulcerative colitis or inflammatory bowel disease (IBS). SSZ performs apoptosis in liver cancer cells by inducing ROS. In CRC, it causes cancer cells to become sensitive to chemotherapy drugs. 5 Since GSH is an intracellular antioxidant, SSZ can inhibit GSH production by inhibiting cysteine uptake, thereby increasing ROS. As a result, the mechanism of action of SSZ is to inhibit GSH production and induce ROS. 1
The signaling of oxidative stress is essential in the pathogenesis and progression of the disease. Although its exact roles and mechanisms are not entirely understood, it is accepted that oxidative stress is involved in the onset and progression of the disease. So, antioxidants may have potential therapeutic effects, especially in combination with commonly prescribed treatments.6,7 GSH is a vital antioxidant that protects cells against ROS and acts as an essential detoxifying agent. Previous studies have shown that the binding of GSH to cisplatin forms a Pt (GS) 2 compound released by multidrug resistance-associated proteins out of cancer cells. Thus, the cell level cisplatin decreases. 8
Selenium (Se) is an essential trace element for humans and other organisms, especially for many cellular processes considering the numerous selenoproteins with essential biological functions. 9
Some selenoproteins are selenoenzymes such as thioredoxin reductase and glutathione peroxidase (GPx), representing basic antioxidant systems for maintaining cellular redox homeostasis. Thus, Se acts as an antioxidant in the body, thyroid hormone metabolism, oxidation reactions, reproduction, and immune function. 9 Selenium is a part of the selenoenzyme GPx, reducing oxidative damage. GPx converts hydrogen peroxide to water by oxidizing glutathione. Glutathione reductase is a flavin-dependent enzyme that uses nicotinamide adenine dinucleotide phosphate (NADPH) (as an electron transferor) to revive the reduced form of GSH. Selenite supplementation enhances the accessibility of GPx among other selenium-affiliate enzymes.10,11
There are several proposed mechanisms to explain the effect of Se on the cell cycle and apoptosis, and it is well recognized that Se plays a crucial role in these processes. However, the mechanisms of action of Se are very complex and not fully understood. They include signaling protein kinases, activation of caspases, phosphorylation of p53, and production of ROS. 12
Zinc is a rare element that is essential for cells' normal functioning. It is a cofactor for the structure and function of various cellular proteins, including enzymes, transcription factors, and structural proteins. Recent studies have shown that zinc plays a role in inhibiting various cancers. Zinc has systemic effects such as regulation of the immune system and direct cellular effects, leading to the regulation of gene expression, bioenergetics, metabolic pathways, signal transduction, and cell invasion. Zinc has also been reported to regulate cell proliferation and growth. 13
Zinc is known to modulate oxidative stress. Oxidative stress is a momentous agent in developing multiple chronic human diseases and dysfunctionalities such as vascular disease, mutagenicity, cancer, neuronal destruction, immune disturbance, and aging processes.14,15 OH•, O2•−, and H2O2 are known as ROS and are constructed unceasingly in vivo under aerobic status. NADPH oxidases utilize NADPH (as an electron transferor) to catalyze the production of O2•− from oxygen and inhibit NADPH oxidases' activity.
The transformation of O2• to H2O2 is catalyzed by the superoxide dismutase (SOD), which includes zinc and copper. It also produces metallothionein, a cysteine-rich component used to clear OH•. Iron and copper ions also catalyze the production of OH• from H2O2. Zinc competes with iron and copper in binding to cell membranes, reducing the production of OH.16,17
This study aimed at evaluating zinc and selenium's synergistic effects with SSZ in inhibiting the growth of colon cancer cells.
Materials and Methods
Ethical committee code: IR.MAZUMS.REC.1398.5966.
Material and reagents
SSZ, Selenium, Zinc, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), thiobarbituric acid (TBA), 2’,7’-dichlorofluorescin diacetate (DCF-DA), and 5,5’-dithiobis-(2-nitrobenzoic acid) (DTNB) were purchased from Sigma-Aldrich.
Cell lines and cell culture
The human CRC cell line (human colorectal adenocarcinoma [HT-29]; ATCC® HTB-38™) was purchased from the Pasteur Institute of Iran. Cell suspensions containing Dulbecco's modified Eagle's medium, 1% pen-strep (penicillin G/streptomycin), and 10% fetal bovine serum were placed in a 75-cm3 cell culture flask and incubated at 37°C in a humid atmosphere of 5% CO2 to reach their logarithmic growth stage. When the primary culture became almost confluent, one cc of trypsin-ethylene diamine tetra acetic acid (EDTA) was added to the flask and incubated for 5 minutes to completely separate the cells from the flask. Then, to evaluate the oxidative stress and cell survival tests, 100 μL of cell suspension was added to each well in 96-well plates, and the plates were incubated for 24 hours. 18
Viability test (MTT)
After 24 hours of incubation, cells were treated with different concentrations of Selenium and Zinc (10, 50, 100, and 200 μM) with a single dose of SSZ (IC50 = 130 μM) 16 and incubated for 48 hours 1, 17. After that, cells were treated with 10 μL MTT solution, incubated for 4 hours, and washed twice with phosphate-buffered saline to remove all remaining materials. Then, 30 μL of dimethyl sulfoxide was added to dissolve the formazan dye; the absorption was read at 630 nm by the Biotek ELx800 Microplate Reader. 19
Reactive oxygen species
Intracellular oxygen-free radicals were measured using the dichloro-dihydro-fluorescein diacetate (DCFH-DA) reagent. When DCFH-DA enters living cells, it is converted to DCFH by intracellular esterase. When it reacts with oxygen radicals, it converts to DCF, which can be detected using a fluorescence microplate reader. Twenty microliters of DCFH solution was added to each well and incubated in the dark for 15 minutes. The absorbance was then read as 485 and 530 nm. 20
Measurement of lipid peroxidation
To measure lipid peroxidation, 100 μL of cell suspension, 100 μL of TBA reagent, and 100 μL of phosphoric acid were mixed well and incubated in a warm water bath for 30 minutes after cooling. Overall, 0.2 mL of n-butanol was added, shaken well, and then centrifuged at 1370g (3500 rpm) for 10 minutes. The n-butanol layer is separated for measurement at 532 nm. 20
Measurement of intracellular GSH
In the falcons' tube containing the studied cells, which were pre-treated with different concentrations of substances, 1.5 mL of trichloroacetic acid and EDTA (10%) were added to precipitate the proteins in the next step. The samples were centrifuged at 3500 rpm for 15 minutes. Then, 1 mL of supernatant was removed, and 2.5 mL of Tris buffer was added at pH = 8.9. Next, 0.5 mL of DTNB (40%) was added and incubated for 15 minutes. The tube was shaken well to obtain a uniform yellow color to finish the reaction. Finally, the absorbance was measured by spectrophotometry at 412 nm. 20
The activity of GPx
GPx reduces lipid hydroperoxides to the corresponding alcohols and reduces free hydrogen peroxide to water. Cell lipid compounds are sensitive to free radicals and react to produce lipid peroxide. GPx enzymes use GSH to reduce peroxides in alcohol and prevent the formation of free radicals. GPxs catalyze the reduction of hydrogen peroxide (oxygenated water) and a wide range of organic peroxides related to alcohol and water using cellular GSH. GPx activity was assessed via A Zell bio kit (Berlin, Germany). GPx activity is based on glutathione reductase assay. The experiment's principle is to lessen the hydroperoxide molecule by GPx, which produces the oxidized form of GSH. 19
The activity of SOD
The SOD participates in the toxic radical dismutation produced during the oxidation steps of energy to O2 and H2O2. In this method, xanthine and xanthine oxidase (XOD) are used to produce superoxide radicals that react with INT [phenyl tetrazolium chloride-5-(nitrophenol-4)-3-(iodophenol-4)-2] to form formazan red. It can be measured at 420 nm. In the presence of the SOD enzyme in the sample of superoxide radicals, it is converted to hydrogen peroxide and O2. Finally, the formation of formazan red color is prevented, and the activity of the SOD enzyme is determined by the degree of inhibition of this reaction.
Briefly, the sample was first mixed with water-soluble tetrazolium solution (containing xanthine and INT); then, SES solution containing XOD was added and incubated at 37°C for 20 minutes. The absorbance of the samples was then measured at 450 nm. Finally, enzymatic activity was calculated in units per mg of protein. One unit of SOD is the amount of enzyme that reduces the rate of reduction or reduction by the enzyme XOD by up to 50%. 19
Measurement of caspase-3 activity
The activity of caspase-3 protease was measured by using a caspase-3 colorimetric assay kit according to the manufacturer's instructions.21,22
Comet assay
For the comet test, the slides were coated with 1% normal melting agarose. The cells exposed to different concentrations of the substance were mixed with 1 mL of 1% low melting agarose, and A layer was placed on the coated slides and kept at 4°C for 10 minutes. The slides were first immersed in an alkaline lysis buffer (pH = 10.0) to remove proteins and cell membranes. They were then placed in an electrophoretic alkaline buffer (pH >13) until DNA was allowed to unwind for 40 minutes. Then, electrophoresis was performed with the same buffer at 300 mA and 25 V for 40 minutes.
Next, the slides were removed and submerged in neutralizing tris buffer solution (pH = 7.5) for 15 minutes and stained with ethidium bromide (20 μg/mL) for 10 minutes. The slides were examined with a fluorescence microscope with magnification × 400, with an excitation filter of 510–560 nm and a barrier filter of 590 nm. DNA damage was measured using the comet score software and expressed as (tail length, % DNA in Tail and Tail Moment), and results were given as mean ± standard error of the mean. 22
Statistical analysis
Statistical analysis was performed by Prism version 3 Software nonlinear regression method, and data were compared with one-way analysis of variance and related post-test (Tukey–Kramer multiple comprehension tests). The same graphic program was used to make the graphs.
Mathematical models were analyzed by SPSS Software, and graphs were made by software Table Care 2D version 5.01.
Results
The effect of selenium and zinc, along with SSZ, on inhibiting the growth of colon cancer cells (HT-29)
Colon cancer cells' treatment with SSZ reduced cell growth by 53.4%. Combination treatment with selenium or zinc and SSZ has further inhibited cancer cells' growth, which is directly related to increased concentration. Combination treatment by “Sel” reduced the cell viability by 53.92% in lower concentrations and 79.31% in higher concentrations. Treatment with “Zn” at 10 μM inhibited cancer cell growth by 55.35%, and by increasing the concentration, in the highest dose, this rate reached 79.41%.
Statistically, concentrations of 100 and 200 μM selenium and zinc significantly differed from the SSZ group (p < 0.05—p < 0.01) and all treatment groups. In comparison between the two supplements, selenium and zinc, at concentrations of 100 and 200 μM, zinc at a concentration of 200 μM had a better effect in inhibiting cell growth (p < 0.001). Compared with the control group, all groups had significant differences (p < 0.0001) (Fig. 1).
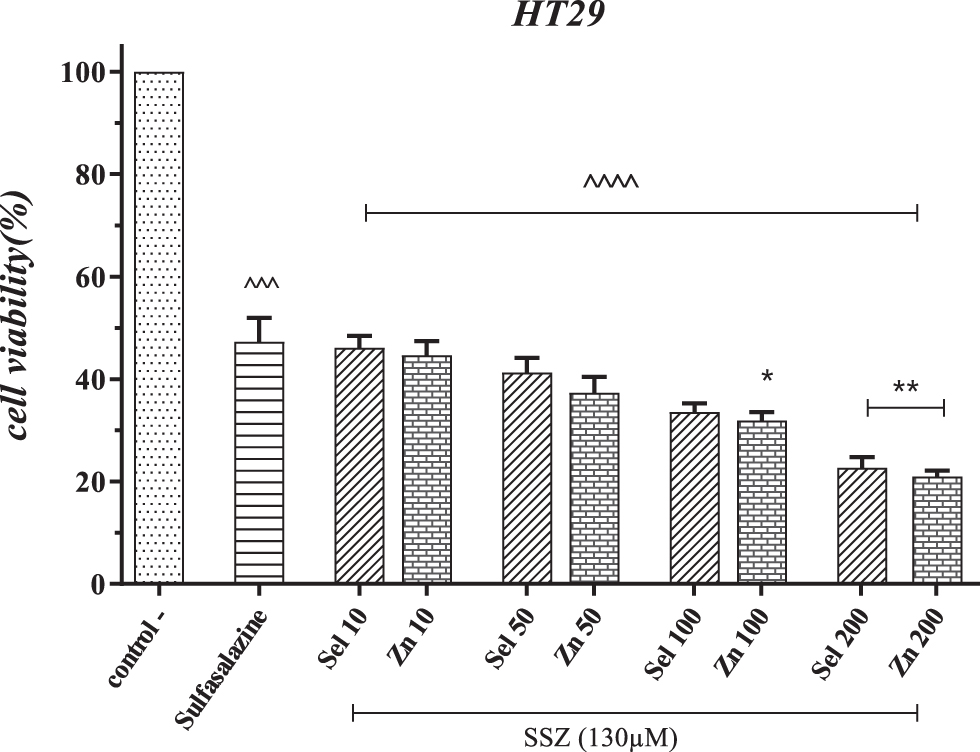
The effect of Sel and Zn in combination with SSZ on inhibiting the growth of HT-29 cells. ^^^p < 0.001, ^^^^p < 0.0001 compared with the control group. *p < 0.05, **p < 0.01 compared with the SSZ group. HT-29, human colorectal adenocarcinoma; Sel, selenium; SSZ, sulfasalazine; Zn, zinc.
The effect of selenium and zinc together with SSZ on the production of ROS in colon cancer cells (HT-29)
The production of ROS has increased in the presence of SSZ, which causes the death of cancer cells. SSZ increased ROS formation by 30.20%. In contrast, concomitant SSZ treatment with zinc or selenium supplements prevents the drug from being expelled by cancer cells, which, in turn, has a more substantial effect on cancer cells and accelerates their death. Oxidants are part of the enzymes' SOD and GPx. Selenium and zinc with high concentrations increased ROS formation by 41.73% and 39.18%, respectively. In contrast to the SSZ group, this amount increased by 11% and 9%, respectively.
Statistically, concentrations of 100 and 200 μM selenium and zinc significantly differed from the SSZ group (p < 0.05). Compared with other treatment groups, concentrations of 100 and 200 μM were also significantly different. Compared with the control group, all groups were significantly different: SSZ group (p < 0.05), Sel and Zn at 10 μM (p < 0.01), and other groups (p < 0.001) (Fig. 2).
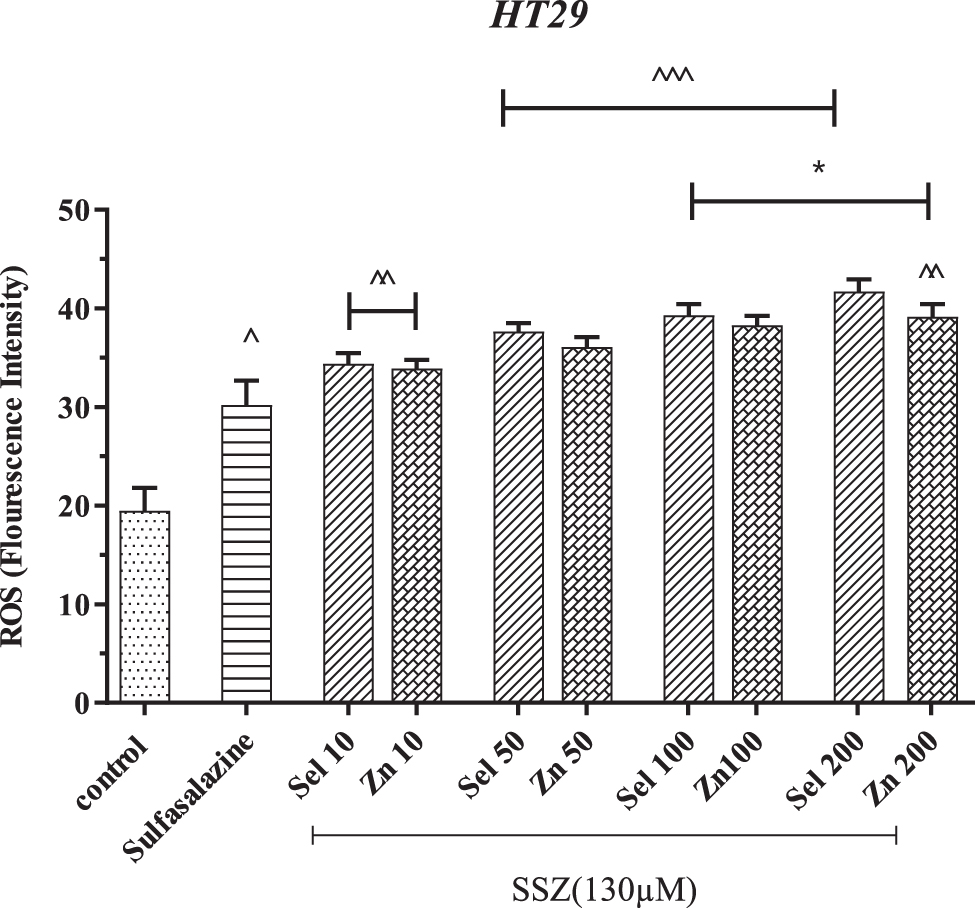
The effect of selenium and zinc together with sulfasalazine on the production of reactive oxygen species in HT-29 cells. ^p < 0.05, ^^p < 0.01, and ^^^p < 0.001 compared with the control group. *p < 0.05 compared with the SSZ group. HT-29, human colorectal adenocarcinoma; ROS, reactive oxygen species.
The effect of selenium and zinc on SSZ-induced lipid peroxidation in the colon cancer cell line (HT-29)
Lipid peroxidation (malondialdehyde [MDA] level) is one of the consequences of oxidative stress that causes chain damage along the body's membranes and macromolecules. Figure 3 shows the increase in SSZ-induced lipid membrane damage in cancer cells. As previously mentioned, zinc or selenium supplementations are co-administered with SSZ as an adjunct to prevent the drug's expulsion by the defense mechanisms of cancer cells. Therefore, we see an increase in lipid peroxidation and cancer cell death by adding these supplements to SSZ-exposed cells.
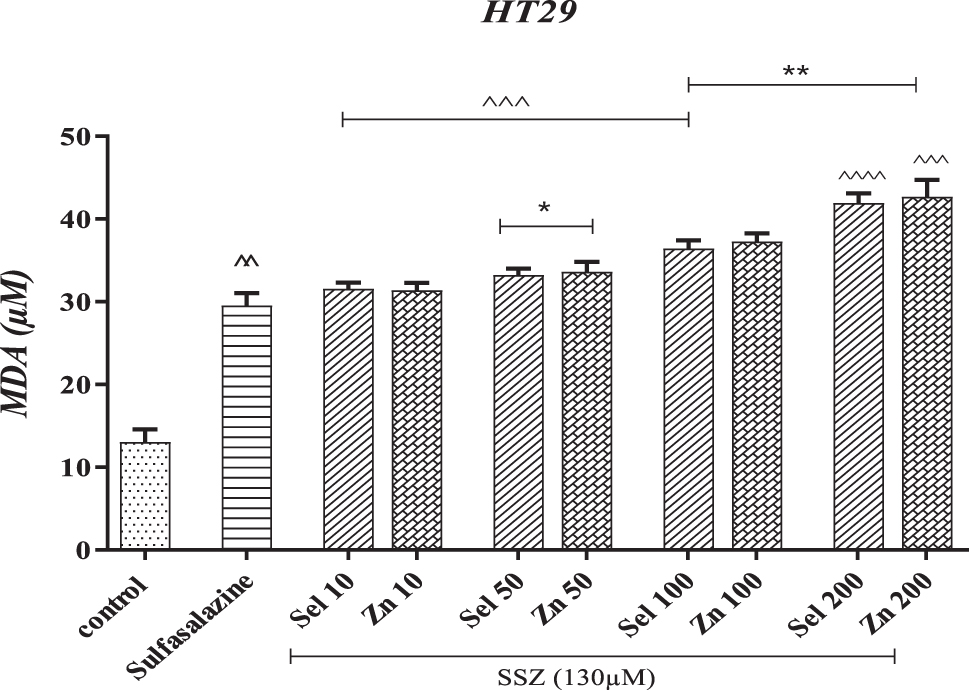
The effect of selenium and zinc on sulfasalazine-induced lipid peroxidation in HT-29 cells. ^^p < 0.01, ^^^p < 0.001, ^^^^p < 0.0001 compared with the control group. *p < 0.05, **p < 0.01 compared with the SSZ group. MDA, malondialdehyde.
In other words, SSZ increased lipid peroxidation by about 17% compared with the control group. However, by adding zinc and selenium along with SSZ, the amount of MDA produced increased from about 32% at a concentration of 10 μM to 43%. With the concentration reaching 200 μM, a 14% increase in lipid induction in HT-29 cell peroxidation was shown compared with the SSZ group.
Statistically, the treatment groups of selenium and zinc at concentrations of 100 and 200 μM differed significantly from the SSZ group (p < 0.01). Concentrations of 50 μM of both substances were also significantly different (p < 0.05). Compared with the control group, all groups had significant differences (p < 0.001) and SSZ (p < 0.01).
The effect of selenium and zinc on intracellular GSH reserves exposed to SSZ in colon cancer cells (HT-29)
As shown in Figure 4, selenium had the most significant effect on SSZ activity when it came to reducing GSH in cancer cells compared with zinc. Decreased GSH causes the gradual death of cancer cells. SSZ reduced the GSH stores of cancer cells by 32.16%. In the groups treated with selenium, the amount of GSH reserves in the highest concentration is 51.5%, and in the groups treated with zinc, this amount is 47.53%.

The effect of selenium and zinc on intracellular GSH reserves exposed to sulfasalazine in HT-29. ^p < 0.05, ^^p < 0.01, and ^^^p < 0.001 compared with the control group. *p < 0.05, **p < 0.01, ***p < 0.001 compared with the SSZ group. GSH, glutathione.
Statistically, the 100 and 200 μM selenium and zinc concentrations were significantly different from the SSZ group (p < 0.05) (p < 0.01). We also see a significant difference in all treatment groups compared with the control group (Fig. 4).
The effect of selenium and zinc on GPx and SOD enzyme activity exposed to SSZ in colon cancer cells (HT-29)
The activity of SOD and GPx enzymes in colon cells decreased in the presence of SSZ. Exposure of cells to selenium and zinc along with SSZ has severely reduced the activity of enzymes so that the activity of enzymes at a concentration of 200 has reached <50% (Figs. 5 and 6). All treatment groups showed a significant difference (p < 0.05) compared with the SSZ group.

The effect of selenium and zinc on GPx enzyme exposed to sulfasalazine in HT-29. ^p < 0.05, ^^p < 0.01, and ^^^p < 0.001 compared with the control group. *p < 0.05, **p < 0.01, and ***p < 0.001 compared with the SSZ group. GPx, glutathione peroxidase.

The effect of selenium and zinc on SOD enzyme exposed to sulfasalazine in HT-29. ^p < 0.05, ^^p < 0.01, and ^^^p < 0.001 compared with the control group. *p < 0.05, **p < 0.01, and ***p < 0.001 compared with the SSZ group. SOD, superoxide dismutase.
The effect of selenium and zinc on caspase-3 activity exposed to SSZ in colon cancer cells (HT-29)
After mitochondrial damage and increased depolarization of the mitochondrial membrane in the presence of SSZ, caspase-3 activity also increased. In the colon cancer cell line, in the presence of selenium, zinc, and SSZ, caspase-3 activity increased and stopped the growth of cancer cells (Fig. 7).

The effect of selenium and zinc on caspase-3 exposed to sulfasalazine in HT-29. *p < 0.05, **p < 0.01 compared with the SSZ group.
The effect of selenium and zinc on tail moment exposed to SSZ in colon cancer cells (HT-29)
Figure 8 of the Tail moment compares different concentrations of selenium and zinc with the SSZ group, indicating a significant difference with the control group (p < 0.0001). As shown in the graph, no significant difference was observed with the SSZ group at a concentration of 10 and 50 μM of Sel and Zn.
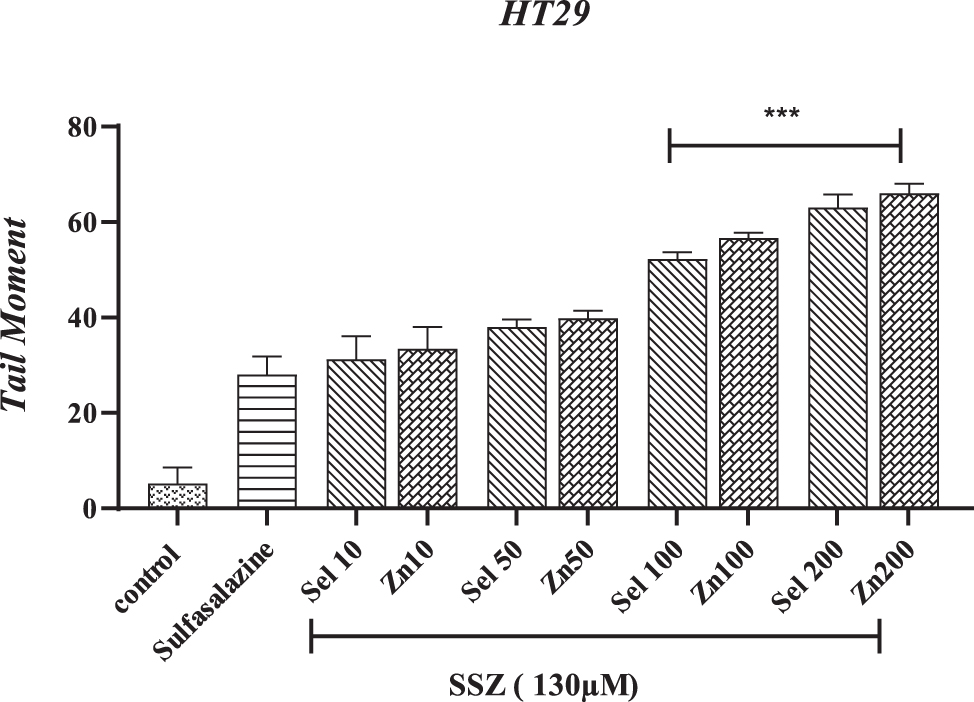
The effect of selenium and zinc on tail moment exposed to sulfasalazine in HT-29. ***p < 0.001 compared with the SSZ group.
Mathematical model
Mathematical equation determination for HT-29 cell line and its treatment with SSZ along with zinc and selenium.
To model the effect of selenium levels on the number of cell viability, a regression model was fitted on the data. The best regression model was chosen based on the result of the F test for coefficient regression and lack of fit parameters. The model expresses the linear relationship between cell viability and selenium levels (y = 47.14–0.12 × ) (Table 1). The lack of fit test shows that this model fits the data within 82% probability.
Mathematical Model of Sulfasalazine + Selenium/Zinc
Therefore, the model cited earlier can predict the effect of different levels of selenium on the percentage of living cells in the range of 0–200 μM. The F test result shows that the model's regression coefficient is significant at 1%, so it can express the effect of different levels of selenium on cell viability. Based on the model, increasing the selenium level reduced the amount of living cells linearly by 0.12% (Table 2; Fig. 9).
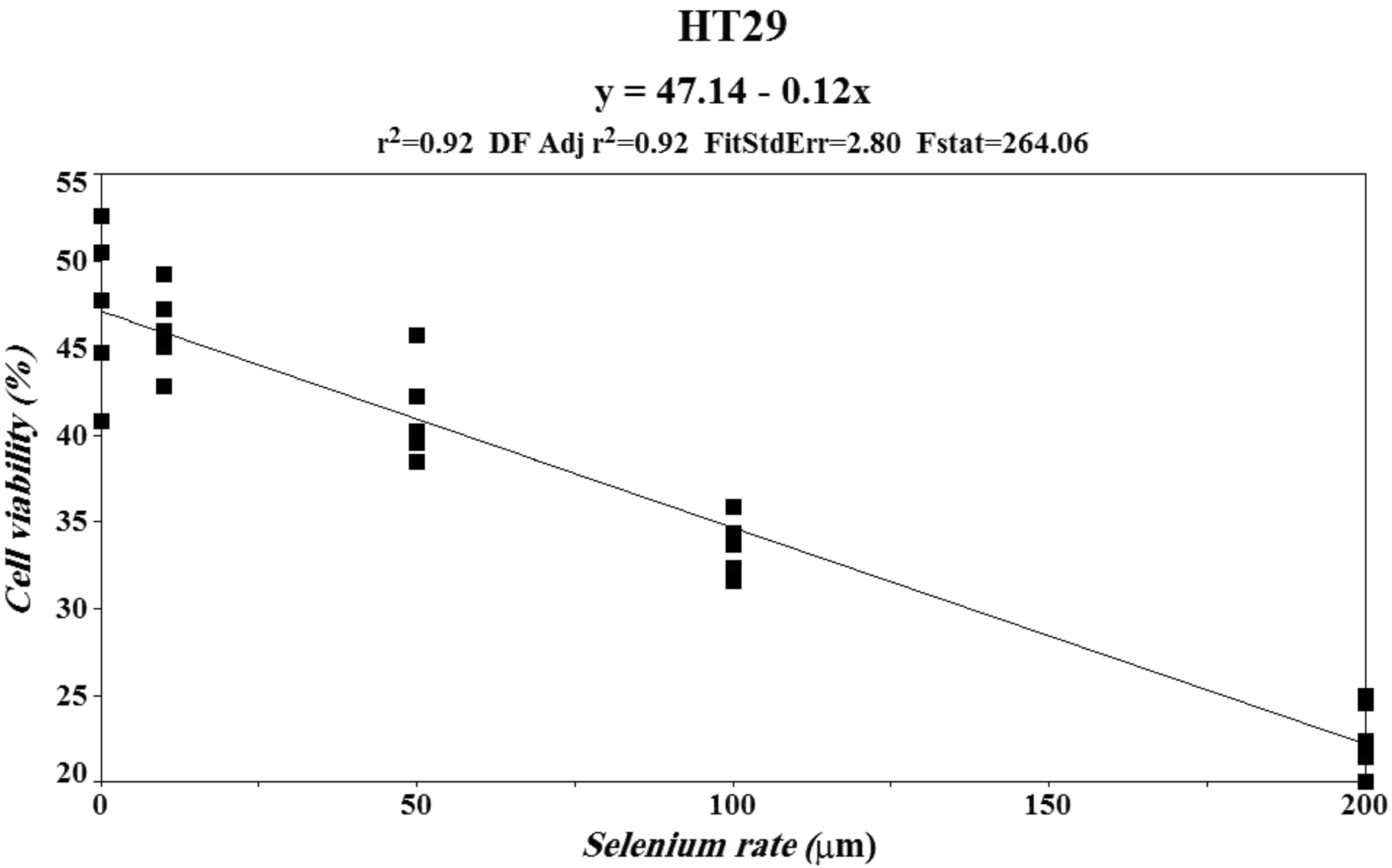
Mathematical model diagram determined for HT-29 cell line and its treatment with sulfasalazine and selenium. This equation shows that with increasing each unit of selenium concentration, the amount of y-axis decreases by 0.12%.
Statistical Parameters of F Test Related to Human Colorectal Adenocarcinoma Cell Line Treated with Sulfasalazine and Selenium
DF, degrees of freedom; HT-29, human colorectal adenocarcinoma; SEL, selenium; SZZ, sulfasalazine.
The model that well expresses the relationship between zinc levels and the number of cell viability is logarithmic (y = 47.26–0.35 × 0.5 lnx) (Table 3). The slope of the curve was higher at concentrations of 0–5 μM, indicating that the effect was more significant during this concentration range. After a concentration of 5 μM, the effect is constant with each increase of zinc, and the slope decreases logarithmically. The lack of fit test shows that this model fits the data with a 94% probability (Table 3; Fig. 10).

Mathematical model diagram determined for HT-29 cell line and its treatment with sulfasalazine and zinc. DF, degrees of freedom.
Statistical Parameters of F Test Related to Human Colorectal Adenocarcinoma Cell Line Treated with Sulfasalazine and Zinc
SSZ, sulfasalazine; ZN, zinc.
Discussion
A long-term clinical experience shows that SSZ is used to treat IBS. The potential SSZ anticancer activity in CRC has yet to be examined. Since the initial discovery of SSZ, an increasing number of anticancer studies have shown this compound in various cancers, including breast cancer, pancreatic cancer, glioma, and chronic lymphocytic leukemia.23–25
Recent studies have shown that SSZ induces apoptosis in several human cancer cells, including esophageal cancer, lung cancer, and glioblastoma cells. 26 Recently, Guo et al have reported that SSZ can inhibit cell growth in hepatocellular carcinoma through autophagy. 27 Most research on the mechanisms of action of chemotherapeutic agents has focused on apoptosis, a major pathway of cell death initiated by genotoxic stress. As a result, apoptosis-related molecules are currently the main target of cancer treatments.
However, autophagy induced by various chemotherapeutic agents can enhance the anticancer potential of drugs. The SSZ treatment activated the extracellular signal-regulated kinase (ERK) pathway and increased the Atg5–12 complex. These results suggest that SSZ may have the same molecular mechanisms as cisplatin. Considering the effects on Akt and ERK signaling pathways, these findings suggest that SSZ-induced autophagy is regulated by Akt inhibition and ERK activation. 26
The results of a study suggest that another form of cell death causes cell death caused by SSZ. On the other hand, at a higher concentration of SSZ, both the levels of caspase 3 (effector caspase) and cleaved (adenosine diphosphate-ribose) polymerase (Poly [ADP-ribose] polymerase [PARP]) (a caspase 3 substrate) were slightly increased. Further, increased expression of microtubule-associated protein 1A/1B-light chain 3 preceded caspase 3 and PARP cleavage. 26
It was previously described by Marino et al that anticancer agents could rapidly initiate autophagy and subsequently activate apoptosis. Another study showed that in human squamous carcinoma cells, SSZ first initiates autophagy, which induces apoptosis. In conclusion, because SSZ induces non-apoptotic cell death at low concentrations, autophagy is the primary type of cell death induced by SSZ.26,28
As it has been determined from the results of this study, SSZ increases caspase 3 activity in colon cancer cells as well.
Oxidative stress is considered a contributing factor in the disease's onset, progression, and severity. However, the functioning mechanisms are not yet entirely clear. Strategies to reduce oxidative stress can improve treatment outcomes. In general, antioxidants have been considered acceptable substances to reduce the content of oxidative stress factors in any disease. 29
Selenium (Se) is a fundamental and unrivaled micronutrient and, as opposed to other trace elements that bind non-covalently to proteins, is contained in polypeptides as the 21st amino acid, selenocysteine. 30 The outcome of further studies (on a smaller measure) showed that Se supplementation was effective on cancers of the prostate, lung, bladder, colon, esophagus, heart, stomach, and thyroid.31–35 Selenium can regulate local invasion and migration by modulating the expression of redox-active proteins, regulating inflammatory and immune responses, balancing the intracellular redox status, and modulating the redox status of multiple proteins and apoptosis. It can affect cancer, block angiogenesis, activates or deactivates essential proteins that regulate cell proliferation, and potentiates phase II carcinogenic detoxifying enzymes. 30
Clinical trial outcomes and epidemiology data do not fully consent to the advantage of Se supplementation. Numerous laboratory and animal studies show that Se is an encouraging prophylactic and antineoplastic chemical. For example, Se optimal concentration renders a special preventive effect, which may be finite. However, determining the most effective Se compound is consistently challenging because different Selenium-containing compounds may affect cells through various mechanisms. Lately, scholars have cited Se valuable as an anti-metastatic factor (containing anti-invasion, anti-migration, and anti-angiogenic capabilities). 30
Studies have shown that selenium has anticancer effects due to oxidation and reduction of selenite. The role of superoxide production was also confirmed by H2Se using SOD. The ROS production in in vitro models has been detected by the reaction of selenite with GSH, and other thiol compounds. 36
Moreover, it is widely recognized that the effectiveness of Seme compounds as chemopreventive and anticancer agents is related to their chemical form and dosage. Dependence of chemical structure has been demonstrated in the conflicting results of two clinical trials using 200 μg/day of selenium supplementation in cancer prevention. 37
Another study reported that metabolites of Se compounds had been shown to induce ROS, which can induce oxidative changes and DNA breakage. Studies have focused on Se-induced stress responses in various cultured cancer cells, suggesting that most of Se's role in cancer prevention is attributed to ROS-induced apoptosis or cell cycle arrest in cancer cells. Consistent with this concept, it has been shown that Se-induced apoptosis in cancer cells can be suppressed by antioxidants and is dependent on p53. 38
All previous studies have shown that selenium reduces the risk of cancer. In combination with GPx and GSH, it increases the amount of intracellular ROS and the death of cancer cells and as our results show, selenium in combination with SSZ reduces the growth of cancer cells.
Zinc plays a severe role in regulating cellular functions and is essential in cancer therapy. Also, zinc restrains thioredoxin reductase, a major mediator in the cellular response to oxidative stress that is often expressed in cancer. 39
There is growing evidence that oxidative stress is essential in developing several chronic human diseases, such as atherosclerosis, vascular disease, mutagenicity, cancer, and nerve damage. Therefore, zinc supplementation may be a beneficial prophylactic chemical for several cancers such as prostate or colon cancer.40,41 A study showed that zinc supplementation in healthy individuals reduces oxidative stress and its associated by-products produced by cells, including MDA and 8-hydroxy deoxyguanosine. 41
A study showed that zinc induces cytotoxicity in HepG2, A549, and Wi38 cells using different concentrations (IC50 = 308.11, 413.02, 463.15 μg/mL). These data showed that ZnSo4 decreased cell viability in malignant and non-malignant cells and induced cytotoxic effects. Cell cycle and apoptosis by flow cytometry showed a significant increase in HepG2 cells damaged by ZnSo4 with cell cycle arrest in the G2/M phase and increased apoptosis. In addition, p53 and casp3 messenger RNA expression levels were increased. The Comet assay method of three different concentrations of ZnSo4 showed that ZnSo4 could cause DNA damage. 42
As this study shows, along with all previous studies, zinc and selenium combined with SSZ have been shown to reduce the cancer cells' growth in colon cancer cells.
Conclusion
This study confirmed that using zinc and selenium might improve anticancer drugs' effectiveness and reduce the process of cell growth. The investigation has not yet precisely defined the role of zinc or selenium in molecular mechanisms involved in the pathophysiology of chronic diseases such as cancer. However, data show that zinc deficiency causes oxidative stress and promotes these diseases. The biochemical and molecular basis of this process needs to be clarified. Therefore, further studies may highlight this nutrient's role in preventing cancer progression.
Footnotes
Authors' Contributions
M.S.: Contributed to conception and study design and management. F.M.: Contributed to the data analysis, writing the article, and drafting the manuscript. B.K.: Contributed to the mathematical model. P.M., M.E., A.S., and R.S.: Contributed to stress oxidative assay and edited the manuscript. All authors read and approved the final manuscript.
Author Disclosure Statement
All authors declare that they have no conflict of interest.
Funding Information
This study was supported by a grant from the Research Council of Mazandaran University of Medical Sciences (Grant No. 5966).