
Abstract
Background and Purpose:
Cyclophosphamide (CP) causes damage to cancer cell DNA and oxidative stress, which occurs with decreased glutathione (GSH) and catalase and increased lipid peroxidation. However, methotrexate (MTX) may cause severe damage to the bone marrow and also cause chromosomal abnormalities in rat bone marrow after several treatments. This study aimed to investigate the protective effect of resveratrol (RES) on cytotoxicity and oxidative stress caused by CP and MTX in the stem cell (SC) line of the rat and evaluate genetic toxicity in rat blood lymphocytes.
Methods:
The rate of cell growth, production of reactive oxygen radicals, malondialdehyde, and GSH stores, mitochondria membrane potential in SCs, and rate of micronucleus in rat blood lymphocytes were investigated. SCs were exposed to a single dose of CP and MTX and different concentrations of RES for 48 hours.
Results:
CP and MTX have caused a disruption in mitochondrial function, which leads to the release of reactive oxygen radicals and increased mitochondrial membrane potential. Consequently, releasing reactive oxygen radicals caused damage to the lipid membrane and DNA while also depleting GSH reserves. However, a significant improvement was observed when SCs were exposed to RES. RES effectively scavenged reactive oxygen species and enhanced GSH reserves, improving mitochondrial function and reducing negative potential within mitochondria.
Conclusions:
RES showed protective effects against CP and MTX-induced cytotoxicity and genotoxicity, which may be attributed to its antioxidant activity. So, it can be considered effective against cytotoxicity and genotoxicity induced by CP and MTX in SCs and rat blood lymphocytes.
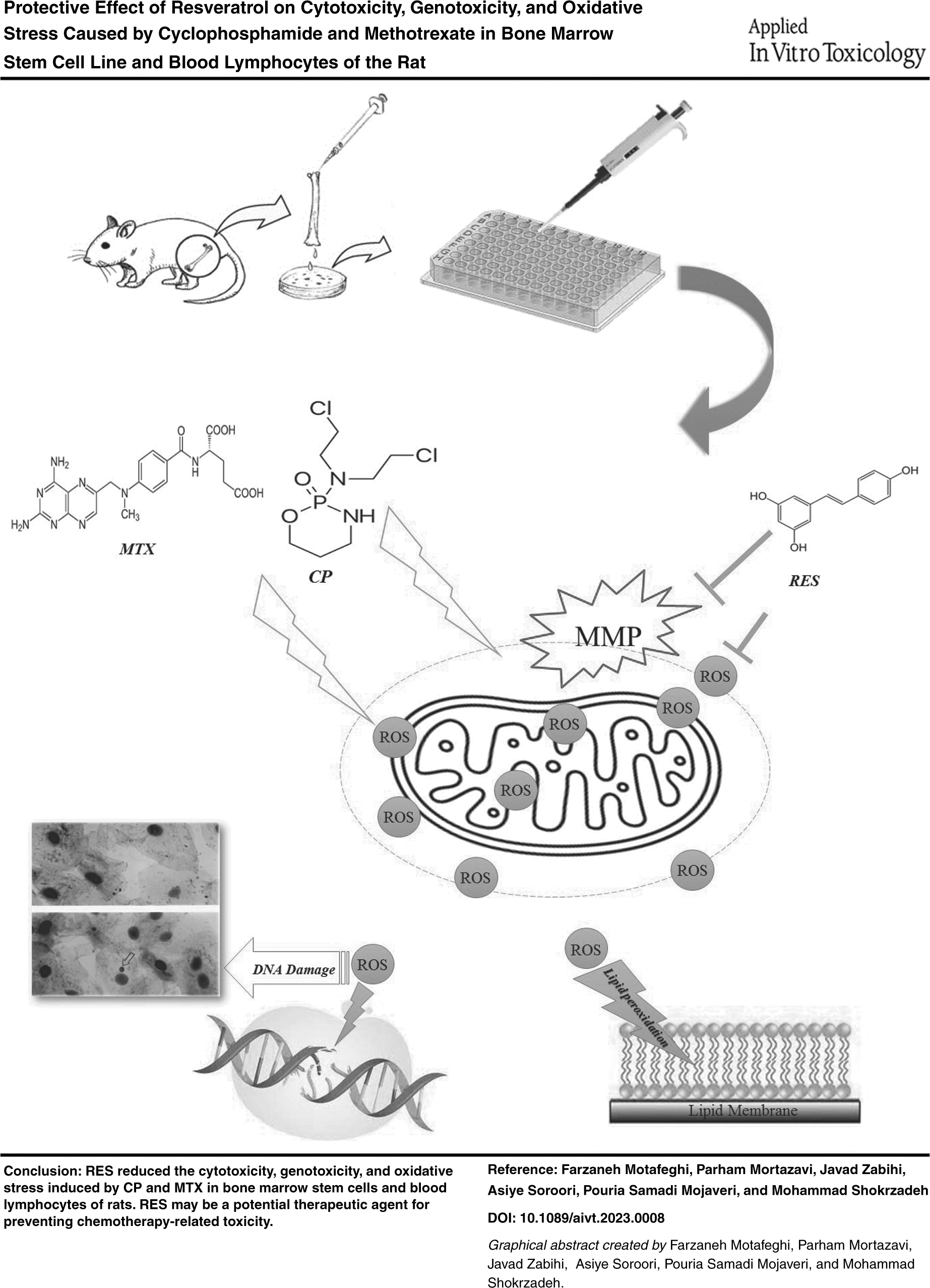
Introduction
For >40
One study found that CP causes oxidative stress, which occurs with decreased glutathione (GSH) and catalase and increased lipid peroxidation (LPO). Interestingly, previous studies have reported that CP does not cause multiple organic toxicities. However, its toxic metabolites, especially at high doses of CP, are associated with the release of large amounts of reactive oxygen species (ROS), including H2O2, HOCl, and O2, and cause oxidative stress in various organs.
Although the mechanisms behind CP-induced multiple organ toxicity are still under investigation, some reports suggest that CP toxicity may lead to endogenous oxidative stress and inflammatory response through the production of free radicals. 4 CP also produces genetic toxicity by producing oxygen/nitrogen free radicals during chemotherapy, causing DNA damage. Therefore, antioxidants are recommended to counteract chemical agent side effects. 5
As an antineoplastic agent, methotrexate (MTX) is widely used to treat acute lymphoblastic leukemia, non-Hodgkin's lymphoma, sarcomas, cancers, breast and testicular tumors, etc. In addition, it is commonly used to treat inflammatory diseases such as rheumatoid arthritis, sarcoidosis, psoriasis, and dermatomyositis. 6 However, MTX may cause severe damage to the bone marrow, gastrointestinal tract, and hematopoietic system. 7
MTX's carcinogenicity, mutagenicity, and teratogenicity have been tested in different systems. MTX also caused chromosomal abnormalities in rat bone marrow after several treatments. It has also been reported to be clastogenic in the human bone marrow. Thus, MTX is highly clastogenic in the bone marrow of rats, even after a single low-dose treatment. 8
Resveratrol (RES) with the chemical formula 3,5,4′-trihydroxy-trans-stilbene is a plant-derived polyphenolic phytoalexin. RES is produced by the enzyme stilbene synthase in response to environmental stresses such as climate change, exposure to ozone, sunlight, heavy metals, and infectious agents such as pathogenic microorganism. 9
Various positive effects of RES have been reported. It has been shown to have anti-inflammatory effects by directly inhibiting COX-2 in humans and inhibiting COX-1, COX-2, and NF-Kappa beta (NF-κB). 10 Other studies have shown that this compound has antioxidant activity, antiobesity, antidiabetes, and beneficial cardiovascular and antiaging effects. 11
There are also various proposed mechanisms, including reduction of cell viability, inhibition of angiogenesis and cell proliferation, induction of programmed cell death (apoptosis), cell cycle arrest, antioxidant effect in normal cells, and pro-oxidant effect in cancer cells. RES also inhibits the NF-κB factor. This factor is a heterodimer protein complex and is inactivated in binding to inhibitor of kappa Bs in the cytoplasm. 12 RES reduces ROS production, inflammation, and cell apoptosis. 13 RES increases NOS gene expression, which increases NO production, and mitochondrial biogenesis and inhibits NF-κB. RES also decreases the expression of tumor necrosis factor-alpha (TNF-α), which leads to the inhibition of NADPH oxidases and prevents the formation of free radicals. 14
This study aimed to investigate the protective effect of RES on cytotoxicity and oxidative stress caused by CP and MTX in the stem cell (SC) line of the rat and evaluate genetic toxicity in rat blood lymphocytes.
Materials and Methods
Material and reagents
The chemicals and reagents utilized in this research were procured from Sigma Chemical Company (Germany), and were of the utmost quality attainable in the commercial market.
Extraction and culture of bone marrow SC
Male Wistar rats weighing 200–20 g and 50 days old were utilized in this experiment. The animals used in this study were purchased from Mazandaran University of Medical Sciences and were kept at a comfortable 27°C with easy access to food and water. Ketamine and xylazine are commonly used as anesthetic agents in rodent research, but they can also be used for euthanasia when administered at high doses. Ketamine is a dissociative anesthetic that blocks the N-methyl-
The combination of ketamine and xylazine produces a synergistic effect that enhances the depth and duration of anesthesia. For euthanasia, we used a dose of 300 mg/kg ketamine and 40 mg/kg xylazine intraperitoneally (IP) for rats, which are higher than the recommended doses for anesthesia. We verified the death of the animals by checking for the absence of heart beat, respiration, and reflexes. We followed the AVMA Guidelines for the Euthanasia of Animals: 2020 Edition and the institutional animal care and use committee (IACUC) protocols for the humane and ethical use of ketamine and xylazine for euthanasia.15–17
Their femurs and calves were removed, and the connective tissues surrounding the bones were removed entirely. The bones were placed in a DMEM containing 15% fetal bovine serum (FBS) and 1% penicillin–streptomycin at 4°C and transferred to a laminar hood. Using sterile scissors, the bone's two ends were split, and the bone marrow was then flushed out, transferred to a falcon tube containing a complete culture medium, and then centrifuged at 699 g for 5 minutes.
After removing the supernatant, the cell sediment was suspended in 5 mL of fresh medium, cultured in a T25 flask, and incubated in a CO2 incubator (37°C, 5% CO2). The nonadherent cell supernatant is removed after 24 hours, and the cell environment is changed every 3 days for 14 days. When the cell density at the bottom of the flask reached a certain level, trypsin-EDTA was used to separate the cells from the bottom and transfer them to new flasks. It took three passage steps to get these cells to be highly pure. 18
The grouping of study
In cytotoxicity test
MTT assay
In total, 90 μl (5 × 104 cells per well) of bone marrow cells was seeded in a 96 microwell plate and incubated for 24 hours. After 24 hours of incubation, positive control was exposed to CP (2 μg/mL), 19 or MTX (0.3 μM), 20 and treatment group was exposed to CP/MTX + RES (25, 50, and 100 μg/mL) and incubated for 48 hours. After incubation, 10 μL of MTT solution was added to each well, with a final concentration of 0.5 mg/mL, and was incubated for 4 hours. All contents of each well were removed and washed with PBS, then 100 μL of DMSO was added to dissolve the formazan dye, and absorption was determined at max = 490 and 630 nm by the Biotek ELx800 Microplate Reader.21,22
Reactive oxygen species
Intracellular oxygen free radicals were measured by the 2′,7′-dichlorofluorescein diacetate (DCFH-DA) reagent. Intracellular esterases convert DCFH-DA to DCFH when DCFH-DA reaches living cells. It transforms into DCF when it interacts with oxygen radicals, which can be detected and read by a spectrophotometer. In total, 20 μl of DCFH solution was added to each well and incubated in the dark for 15 minutes. A fluorescence microplate reader was then used to read the absorbance at 485 and 530 nm.23,24
Measurement of LPO
LPO was determined by measuring the generation rate of thiobarbituric acid reactive substances and represented as malondialdehyde (MDA) equivalents. For measuring LPO, 100 μL of TBA reagent, 100 μL of cell suspension, and 100 μL of phosphoric acid were mixed well and incubated in a warm water bath for 30 minutes after cooling. Then 0.2 mL of n-butanol was added and shaken well, then centrifuged at 3500 rpm for 10 minutes. The n-butanol layer is separated for measurement at 532 nm by the Biotek ELx800 Microplate Reader.25,26
Measurement of intracellular GSH
In total, 1.5 mL of trichloroacetic acid, and EDTA (10%) were added to the falcons' tube containing the examined cells, previously treated with various substance concentrations, to precipitate the proteins in the following step. The samples were centrifuged at 3500 g for 15 minutes. Then 1 mL of supernatant was removed, 2.5 mL of Tris buffer was added with pH = 8.9, and 0.5 mL of 5,5′-dithiobis(2-nitrobenzoic acid) (40%) was added and incubated for 15 minutes. To complete the reaction, the tube was vigorously shaken to produce a uniform yellow color. Finally, the absorbance of the solution was measured at 412 nm using a spectrophotometer.23,27
Determination of the mitochondrial membrane potential
To test the potential of the mitochondrial membrane, rhodamine 123 is utilized as a reagent. Ten microliters of rhodamine 123 is added to the samples after dissolving mitochondrial sediment in MMP buffer. The absorbance of the sample was measured using the fluorescence spectrophotometer Schimadzou RF-5000U with the excitation and emission wavelengths of 490 and 535 nm, respectively. 28
Micronucleus assay
The grouping of study
Four healthy rat blood samples were obtained. Each sample was placed in a 37°C water bath. Then, 0.5 mL of blood and 4.5 mL of DMEM culture were added to each well. Phytohaemagglutinin (2% of the total volume) was added and incubated for 24 hours to stimulate the proliferation of blood cells. Positive control was exposed to CP (2 μg/mL), or MTX (0.3 μM); treatment 1 group was exposed to CP or MTX + RES (25, 50, and 100 μg/mL); and treatment 2 group was exposed to RES (100 μg/mL) and incubated for 48 hours. For inhibiting cellular cytokines, 3.6 μL of cytochalasin B (Cyt-B) was administered to each well 48 hours after adding PHA.
At the finish of the incubation period, the contents of each well were transferred to a centrifuge tube and centrifuged for 6 minutes. The lymphocytes were then centrifuged with KCl solution and methanol–acetic acid fixation solution, respectively. Slides were covered with two to three drops of the remaining suspension and placed in Giemsa paint solution for 20 minutes. The number of cells with two nuclei and micronuclei was counted using 40 and 100 magnification light microscopes.25,29
Statistical analysis
The software Prism, version 8, was used to conduct statistical tests. To determine the differences between groups, an analysis of variance followed by Tukey's test was employed (p < 0.05).
Results
CP and MTX inhibit the growth of SCs by inducing oxidative stress and DNA damage
Exposure of SCs to a single dose of CP and MTX inhibits the growth of SCs. RES has also inhibited the cytotoxicity of these two substances at different concentrations. Compared with the MTX and CP group, RES at the concentration of 50 and 100 μg/mL had significant differences (p < 0.01) and (p < 0.05), respectively (Fig. 1).

Cytotoxicity of CP and MTX and protective effect of RES on bone marrow stem cell line. ***p < 0.001 ****p < 0.0001 compared with control group. #p < 0.05 compared with CP group. $p < 0.05 $$p < 0.01 compared with MTX group. CP, cyclophosphamide; MTX, methotrexate.
Oxidative stress factors have a substantial impact on inhibiting and decreasing SC development. As shown in Figure 2, CP and MTX induced ROS in SCs by ∼80% and 60%, respectively. Exposure of SCs to different concentrations of RES attenuates ROS production. The concentration of 50 and 100 μg/mL of RES showed a significant difference compared with the CP and MTX groups (p < 0.05, p < 0.01) (Fig. 2).

Measurement of ROS values after cell treatment with CP and MTX and evaluation of the protective effect of RES in different concentrations. ***p < 0.001, ****p < 0.0001 compared with control group. ##p < 0.01 compared with CP group. $p < 0.05, $$p < 0.01 compared with MTX group. RES, resveratrol; ROS, reactive oxygen species.
MDA is the end product of LPO resulting from the free radical reaction with a lipid membrane. CP and MTX induced LPO up to ∼60% and 50% (p < 0.001), respectively, whereas exposure of SCs to RES reduced the MDA level up to 50% at high concentrations (p < 0.05) (p < 0.001) (Fig. 3).
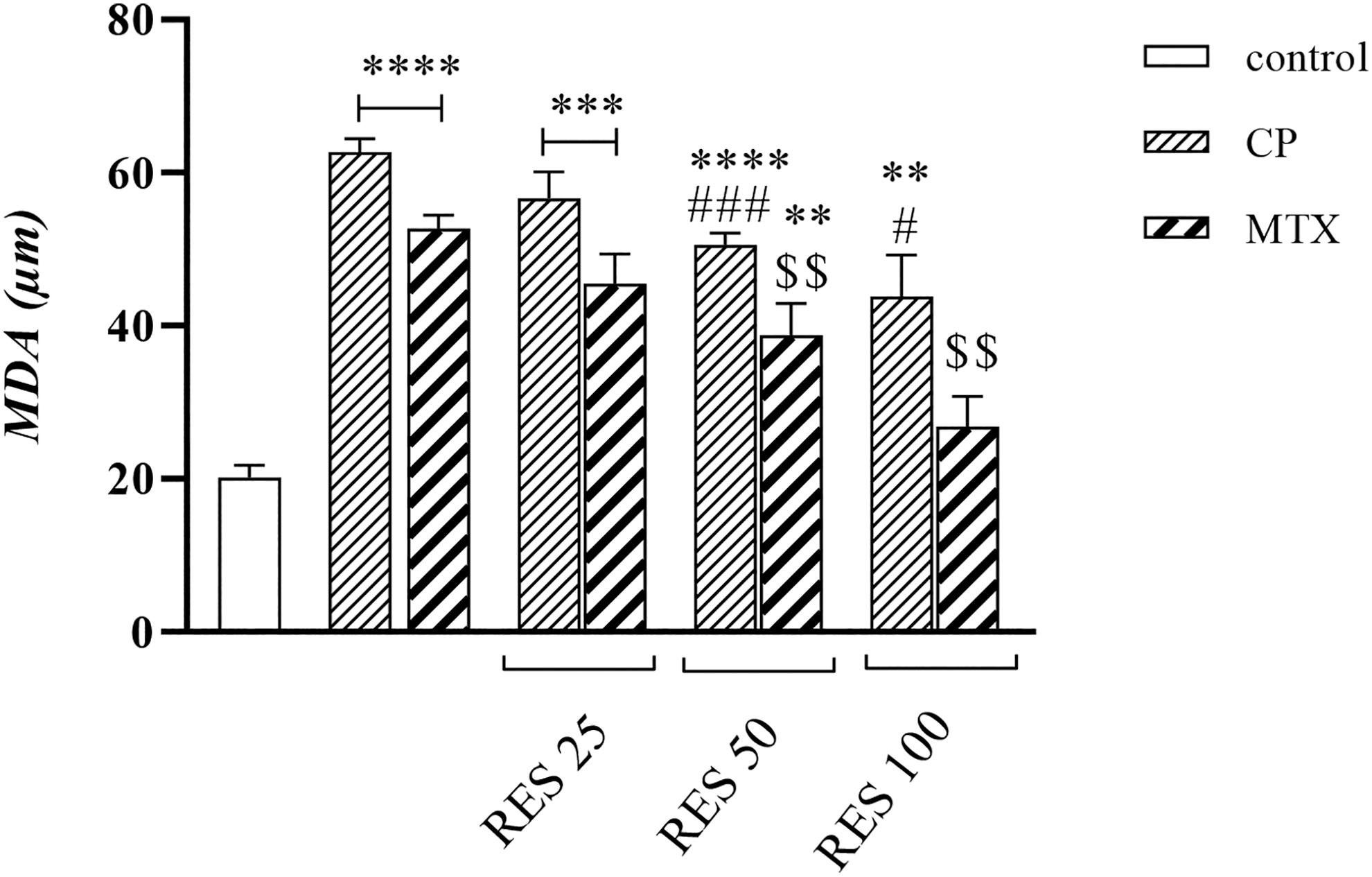
Measurement of MDA values after cell treatment with CP and MTX and evaluation of the protective effect of RES in different concentrations. **p < 0.01, ***p < 0.001, ****p < 0.0001 compared with control group. #p < 0.05, ###p < 0.001 compared with CP group. $$p < 0.01 compared with MTX group. MDA, malondialdehyde.
The intracellular antioxidant capacity declines as oxidative factor generation increases. Both CP and MTX reduce GSH by up to 60% and 50%, respectively, through inducing oxidative stress. So, SCs exposed to RES improved GSH stores up to 80% at high concentrations (Fig. 4).
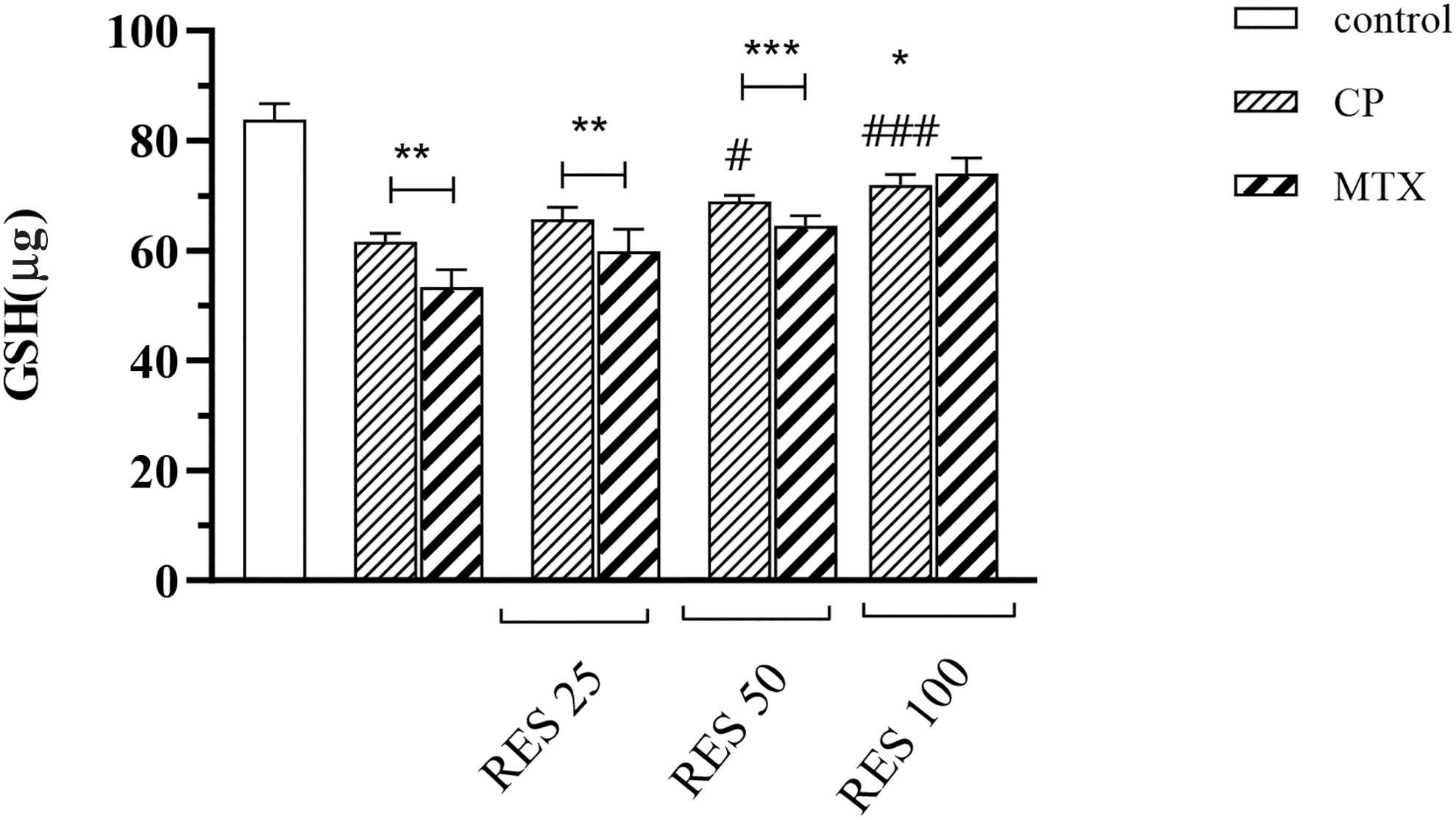
Measurement of GSH values after cell treatment with CP and MTX and evaluation of the protective effect of RES in different concentrations. *p < 0.05, **p < 0.01, ***p < 0.001 compared with control group. #p < 0.05, ###p < 0.001 compared with CP group. GSH, glutathione.
Mitochondrial dysfunction results in the release and accumulation of electrons inside the mitochondria, leading to the loss of potential of the mitochondrial membrane and a negative potential within the mitochondria. The fluorescence intensity increases as the mitochondrial membrane potential increases. The mitochondrial membrane potential degradation rate was most significant in groups administered CP and MTX. Different doses of RES improve mitochondrial membrane potential and mitochondrial function (Fig. 5).

Collapse of MMP in bone marrow stem cell after incubation with CP and MTX and the protective effect of RES at different concentrations. **p < 0.01, ***p < 0.001 ****p < 0.0001, compared with control group P group. #p < 0.05, ##p < 0.01, ###p < 0.001 compared with CP group. $p < 0.05, $$p < 0.01, $$$p < 0.001 compared with MTX group.
CP and MTX inhibit the growth of SCs by inducing DNA damage
CP and MTX were used as clastogenic agents in the micronucleus test. The number of micronuclei produced by these two substances is ∼20%. When lymphocytes were exposed to different concentrations of RES, the results showed that RES reduced the amount of CP and MTX-induced genotoxicity due to its antioxidant properties. At a concentration of 25 μg/mL, the number of micronuclei decreased to 17%, and with increasing the concentration to 100 μg/mL, it decreased to 10%. The RES group of 100 μg/mL also indicates that this extract did not cause genetic toxicity at this concentration (Fig. 6).

Percentage of micronuclei produced in lymphocytes of blood samples in vitro after exposure to CP and MTX and the protective effect of RES at different concentrations ***p < 0.001, ****p < 0.0001 compared with control group. ##p < 0.01, ###p < 0.001, ####p < 0.0001 compared with CP group. $p < 0.05, $$p < 0.01 compared with MTX group.
Discussion
Results of this study showed that CP and MTX have clastogenic and cytotoxic potential on the rat's bone marrow and peripheral blood cells. Also, RES attenuates CP and MTX cytotoxicity and genotoxicity. Bone marrow cell line exposure to CP and MTX instigated chromosomal damage, and an increase in the frequency of micro nuclei also increased oxidative stress factors. A significant reduction in the MN ratio was also observed in RES-treated groups.
A healthy diet rich in fruits and vegetables that contain antioxidants has grown crucial as people's understanding of diet's role in human health and lifespan has increased. Molecules found in foods with antioxidant activities can reduce the damage that ROS can do to cells. RES has been studied for its numerous biological activities, positive impacts on human health, and defense against several illnesses, including cancer neuronal damage, skin disorders, and aging-related consequences. 30
RES (3,40,5-trihydroxystilbene) is a phytoalexin. It is available in both cis and trans isomers. Trans-RES is the more common and stable form of the compound that transforms into cis-RES when exposed to sunshine or UV rays. Cis-RES is a highly unstable compound. In humans, RES is rapidly absorbed, with plasma concentrations peaking ∼30 minutes after oral administration, and 70% to 75% of uptake is by trans-epithelial diffusion. The bioavailability of RES is <1%. The main metabolic pathways of RES are glucuronidation and sulfation in the intestine and liver, which help limit the bioavailability of RES. 31
It has been shown that RES is a highly effective scavenger of several oxidants, including superoxide anion, hydroxyl radical, hydrogen peroxide, nitrous oxide, oxygen radical, and peroxynitrite. The antioxidant effects of RES are attributable to the existence of phenolic rings with three hydroxyl groups at positions 3, 4, and 5, as well as a conjugated double bond and the potential for electron displacement within the structural molecule. The hydroxyl group is necessary for the radical-scavenging activity of RES, but it works in synergy with the hydroxyl groups at positions 3 and 5. 32
The main ways that RES can neutralize free radicals are by transferring hydrogen atoms, transferring electrons, or losing protons in a sequence (SPLET).32–34 Studies have demonstrated that RES exhibits chelating properties through its ability to bind to metal ions, inhibiting the excessive generation of hydroxyl radicals and subsequent oxidative reactions. 35 In addition to its ROS/reactive nitrogen species (RNS) inhibiting effect, intracellular ROS/RNS generation inhibition also contributes to RES 's antioxidant properties.
In various experimental models, the activation of NADPH oxidase raises oxidative stress. Treatment of TNF-α increases NADPH oxidase activity, ROS/RNS production, and oxidative damage. RES blocks the formation of ROS from the NADPH oxidase system through the downregulation of NADPH oxidase 1. 36 Treatment with trans-RES eliminates vascular NADPH oxidase activity and subsequently reduces cellular superoxide anion production. 37
Reduced mitochondrial metabolism is the cause of elevated ROS levels in the mitochondria. Therefore, mitochondrial ROS production is compromised by the stimulation of mitochondrial biogenesis, which increases individual mitochondrial mass and decreases the flow of electrons in the mitochondrial unit. 38 RES has been demonstrated to increase mitochondrial biogenesis, inhibiting ROS production. In the presence of RES, cultured human coronary artery endothelial cells showed high levels of mitochondrial mass and mitochondrial DNA content, protein expression of electron transport chain components, and mitochondrial biogenesis. 39
DNA is sensitive to oxidative/nitrous damage due to the presence of ROS/RNS. Oxidative attacks on DNA can cause basal changes, deoxyribose damage, single- or double-strand breaks, and cross-linking. 40 Oxidative DNA damage is a prominent cause of mutations in living organisms and is linked to various disorders, such as neurovascular disease, cancer, and aging. 41 In addition, RNS interaction with DNA can cause nitrate DNA damage by forming 8-nitroguani. 40 RES exhibits promising properties as a chemical compound that may possess antimutagenic and anticancer effects.
Protecting DNA from oxidative damage and inducing DNA repair are crucial mechanisms that underlie the chemical characteristics of RES. The administration of RES has been demonstrated to induce oxidative DNA damage in both male and female mice. Lowering blood pressure and preventing strokes are associated with reduced levels of 8-OHdG, a marker of DNA damage caused by oxidative stress. 42 RES reduces DNA damage in sepsis-induced liver cells and kidneys in rats, possibly by preventing the breakdown of oxidative DNA strands and activating DNA repair processes. 43 RES reduced sepsis-induced DNA damage in Wistar rat lymphocytes, suggesting that RES may prevent septic oxidative DNA damage. 44
Numerous studies have shown that RES has scavenging and antineoplasm properties, reduces oxidative stress factors, and protects DNA from toxic compounds. Also, this study showed that RES showed antioxidant properties against cytotoxicity and genotoxicity caused by CP and MTX in rats' SCs and blood lymphocytes.
Our study's findings align with the research conducted by Gore et al., which demonstrated that the administration of CP to mice resulted in a considerable increase in DNA damage in bladder cells. The impact of CP on bladder cells may be attributable to the accumulation of free radicals. 45 The study done by Mohamed and Aly also showed that CP induced a genotoxic effect in Swiss albino mice. 46 The reduction of enzyme activity and nonenzymatic antioxidants by CP may be due to its active metabolites' production of free radicals. 47
Numerous studies have shown that MTX-induced cytotoxicity is associated with cellular and mitochondrial GSH depletion and ROS production, ultimately leading to oxidative stress and cell death.48,49 Both in vitro and in vivo animal studies have demonstrated that the administration of MTX alters the functioning of antioxidant enzymes, including GSH peroxidase, GSH reductase, catalase, and superoxide dismutase in the spinal cord, intestinal mucosa, and liver tissues. In rat hepatocytes, MTX caused mitochondrial damage and the release of cytochrome c.49,50 Another study showed that MTX significantly induced MN in polychromatic erythrocytes and significantly increased total chromosomal aberrations. 51
However, our study identified CP and MTX-induced DNA damage and oxidative stress on rats' bone marrow cells and blood lymphocytes.
Conclusion
Based on the empirical evidence presented in this study and the many investigations conducted over the preceding decades, mitochondria play a crucial role in disease. The usage of antioxidants can improve the harmful effects of drugs, both by enhancing mitochondrial function and accelerating the recovery from sickness. This study demonstrated that MTX and CP impede cell growth and produce genotoxicity.
The effects of RES on the cytotoxicity and genotoxicity generated by MTX and CP were accompanied by an improvement in oxidative stress markers and antioxidant defense. Owing to its antioxidant properties and its ability to eliminate free radicals, it has likely been able to decrease the harmful effects of medications on cells.
Authors' Contribution
M.S. contributed to conception and study design and management; F.M. contributed to conception and study design and management, in vitro study analysis data, writing the article, and drafting the article; P.M. and P.S.M. contributed to editing and revising the article; and J.Z. and A.S. contributed to in vitro study. All authors read and approved the final version of the article.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Ethical Approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed.
Footnotes
Author Disclosure Statement
All authors declare that they have no conflicts of interest.
Funding Information
This study was supported by a grant from the Research Council of Mazandaran University of Medical Sciences, IR.MAZUMS.REC.1400.311/IR.MAZUMS.REC.1400.407