
Abstract
The mammalian AP-endonuclease (APE1/Ref-1) plays a central role in the repair of oxidized and alkylated bases in mammalian genomes via the base excision repair (BER) pathway. However, APE1, unlike its E. coli prototype Xth, has two unique and apparently distinct transcriptional regulatory activities. APE1 functions as a redox effector factor (Ref-1) for several transcription factors including AP-1, HIF1-α, and p53. APE1 was also identified as a direct trans-acting factor for repressing human parathyroid hormone (PTH) and renin genes by binding to the negative calcium-response element (nCaRE) in their promoters. We have characterized APE1's post-translational modification, namely, acetylation which modulates its transcriptional regulatory function. Furthermore, stable interaction of APE1 with several other trans-acting factors including HIF-1α, STAT3, YB-1, HDAC1, and CBP/p300 and formation of distinct trans-acting complexes support APE1's direct regulatory function for diverse genes. Multiple functions of mammalian APE1, both in DNA repair and gene regulation, warrant extensive analysis of its own regulation and dissection of the mechanisms. In this review, we have discussed APE1's own regulation and its role as a transcriptional coactivator or corepressor by both re-dox-dependent and redox-independent (acetylation-mediated) mechanisms, and explore the potential utility of targeting these functions for enhancing drug sensitivity of cancer cells. Antioxid. Redox Signal. 11, 621–637.
Introduction
In view of its multiple functions, it may not be surprising that APE1-nullizygous mice have early embryonic lethality, and that no APE1-null mouse embryo fibroblast (MEF) line could be established (152). It was, however, still unclear which function(s) of APE1 are absolutely required during early embryogenesis. Because APE1 is the major enzyme responsible for repairing endogenous AP sites, it is not surprising that its DNA repair function is necessary for embryonic development. However, APE1's redox function has recently been shown to be involved in embryonic hematopoiesis and in CD40-mediated B cell activation and thus in regulation of immune responses, indicating that APE1's transcriptional regulatory functions may be also important for early embryonic development (96, 97, 162). Recently, we and others have shown that APE1 inactivation induces apoptosis in MEF conditionally nullizygous for endogenous APE1 (MEFnull) and in many tumor cell lines (47, 70). Using a complementation assay, we also showed that both the active site for repair and acetylation sites (presumably for regulatory function) of APE1 are required to prevent apoptosis of APE1-null MEF (70). Unexpected essentiality of APE1's acetylation-dependent regulatory activity suggests that APE1 is a co-regulator of essential gene(s). Although Cys65 of APE1 was proposed to be the redox active site, lack of embryonic lethality of Cys65Ala knock in mutation in transgenic mice or APE1 conditional null mutation in MEF suggested that the redox function of APE1 is not essential for survival (70, 110, 142). This could alternatively indicate that Cys65 is not involved in APE1's redox function. On the other hand, our novel observation of modulation of APE1's transcriptional function by acetylation and essentiality of acetyl acceptor Lys have provided an opportunity for elucidating the in vivo role of this modification in both negative and positive regulation of diverse genes. Moreover, it is important to identify essential vs. nonessential regulatory functions of APE1, and whether redox activation is linked to acetylation-dependent regulation. In this review, we will summarize APE1's own regulation and its redox-dependent and acetylation-mediated transcriptional function and explore the potential of this protein as a therapeutic target in drug sensitization of cancer cells.
DNA Repair Function of APE1
Multiple oxidative DNA damage such as strand breaks, base loss, and base modifications are caused by reactive oxygen species (ROS) that are generated endogenously or due to environmental stress (2, 15). Nearly all oxidized forms of DNA bases (as well as methylated or inappropriate bases) are repaired via the BER pathway which is initiated with excision of the damaged base by a DNA glycosylase to generate AP site (65, 87, 98). APE1, the second enzyme in the BER pathway, then hydrolyzes the phosphodiester backbone immediately 5′ to an AP site to produce 3′ OH group and 5′ deoxyribose-5-phosphate (25, 31). Following removal of this blocking group via dRP lyase activity of DNA polymerase β repair DNA synthesis, followed by DNA ligase action restores genome integrity (126). Oxidized base-specific DNA glycosylases have intrinsic AP lyase activity and cleaves the DNA strand 3′ to the AP site (65, 87). The resulting 3′ blocking group is removed by APE1 (or in some cases polynucleotide kinases) in the next step of repair (20, 147). APE1's 3′ phophodiesterase activity is also involved in repairing DNA single-strand breaks with 3′ blocking group directly generated by ROS (71). Unrepaired AP sites also lead to DNA strand breaks, apoptosis, and increases cytotoxicity (92). Thus, the DNA repair function of APE1 protects the cell from both endogenous and exogenous DNA damage. All APEs have dual activities as an endonuclease and a 3′phosphodiesterase (25, 31). However, mammalian APE1's endonuclease activity is quite strong relative to its 3′ exonuclease/phosphodiesterase activity (20, 25, 148). Furthermore, unlike Escherichia coli Xth with potent 3′ phosphatase/phosphodiesterase activities, the mammalian APE1 has extremely weak 3′ phosphatase activity that would be required to remove 3′ phosphate directly generated by ROS or due to AP lyase activity of mammalian glycosylases, NEIL1 and NEIL2 (66, 148). We have shown that NEIL-dependent BER utilizes polynucleotide kinase rather than APE1 (148). APE1 also coordinates BER as an assembly factor by interacting with downstream BER protein such as DNA polymerase β, X-ray cross-complementing-1 (XRCC1), proliferating nuclear antigen (PCNA), and flap endonucelase (FEN1) (27, 38, 73). A recent study shows that Bcl2, an anti-apoptotic protein directly interacts with APE1 and inhibits AP site repair by downregulating AP-endonuclease activity of APE1 (160). Exposure of lung cancer cells to the DNA damaging agent promotes Bcl2 accumulation and association with APE1 in the nucleus (160).
Regulation of APE1 Expression
Although APE1 is ubiquitously expressed in cells and tissues, its expression and subcellular localization level appear to be cell-type specific (79, 132). APE1 is regulated at both transcriptional and post-transcriptional levels. Expression of APE1 in mouse NIH3T3 cells was found to be cell cycle dependent with the highest level of APE1 in early or middle S-phase, pointing to a particular function of APE1 in this phase of cell cycle (46). The effects of ROS on APE1 induction have been extensively studied. We and others have shown that hydrogen peroxide (H2O2) and hypochlorous acid (HOCl) acts as inducers of the APE1 gene (56, 120). Subsequently, several in vivo and in vitro studies confirmed APE1 gene activation by oxidative stress (57, 111). This observation is of particular interest, because H2O2 and HOCl are endogenously formed during inflammatory response of macrophages and lymphocytes. Endogenous ROS may elevate the level of DNA damage which then signals an increase in APE1 level, thus enhancing the BER capacity. Indeed, induction of APE1 was found to be accompanied by an adaptive response of cells to the cytotoxic and clastogenic activity of oxidative agents, indicating its physiological relevance of the phenomenon (42, 56, 120). Consistent with this, Helicobacter pylori infection which induces oxidative stress activates APE1 expression in gastric epithelial cells (29, 30). Similarly, induction of oxidative stress was shown to be involved in the enhanced nuclear translocation of thioredoxin (TRX) and APE1 and augmentation of the APE1/NF-κB complex formation in the parenchyma cells of injured lung (52). In many cell types, ROS-mediated activation of APE1 involves two steps. In the first step, APE1 translocates from the cytoplasm to the nucleus. In B-lymphocytes and thyroid cells, such translocation is fairly rapid, within an hour, whereas in HeLa and other cells the process takes many hours (120, 134, 136). The second step involves de novo protein synthesis via transcriptional activation of the APE1 promoter, because various agents that block transcription or protein synthesis, also abolish induction of APE1 (120). Additionally, APE1 induction is associated with an increase in AP-endonuclease activity and cell's resistance to cytotoxic effect of H2O2, methyl methane sulphonate (MMS), bleomycin, and γ-radiation (42, 56, 120). Transiently overexpressed APE1 protects cells against genotoxicity and cell killing provoked by ROS (42). However, whether protection against ROS-induced cell killing by APE1 is due to of its repair or transcriptional regulatory functions or both is still unknown.
ROS activates APE1 promoter-dependent reporter expression in Chinese hamster (CHO) cells. Mutation of a CREB-binding site (CRE) abolished the H2O2-mediated activation of the APE1 promoter (57). Interestingly, a recent study identified an AP-1/CREB binding site in the mouse promoter which is essential for arsenite-induced transcriptional activation of APE1 in mouse 10T1/2 cells (48). This study showed that arsenite induced APE1 mRNA and protein levels in mouse fibroblasts, and that ATF4/c-Jun heterodimer was the responsible transcription factor for this activation. Moreover, suppression of APE1 or of ATF4 enhanced arsenite cytotoxicity in both mouse 10T1/2 and human lymphoblastoid TK6 cells (48). High conservation of this CREB-binding site in the human (hAPE1) and mouse (mAPE1) promoters highlights its importance in oxidative stress-induced transcription activation (63). In this context, the contribution of APE1 to cellular arsenite resistance is consistent with its proposed DNA repair function in BER under oxidative stress and suggests that APE1 activity may become limiting in DNA repair under certain conditions. Another transcription factor, Egr-1, a member of the immediate-early transcription factors that are rapidly and transiently induced by ROS, binds to the APE1 promoter and activates luciferase expression (102, 111). Pines et al. (111) identified a consensus Egr-1 binding site in the proximal APE-1 promoter. Moreover, they showed that APE1 increases Egr-1's cis element-binding activity after H2O2 treatment (111). Thus, activation of both APE1 and Egr-1 after oxidative stress seems to represent a positive autoregulatory loop between APE1 and Egr-1 and that could explain the early transcriptional activation of APE1 expression.
Other external stimuli such as hormones and cytokines modulate APE1 expression. Thyrotropin (TSH) induces APE1 expression in thyroid cells (5, 131, 134). Similarly, human chorionic gonadotropin has been demonstrated to enhance APE1 mRNA synthesis in murine Leydig cells (128). IL-2-dependent APE1 upregulation has also been demonstrated in a murine Pro-B cell line (156). Interestingly, Helicobacter pylori-induced IL-8 activation in gastric epithelial cells was found to be dependent on APE1 (103). Another recent study demonstrated that ATP-mediated purinergic receptor activation upregulates APE1 expression in human tumor thyroid cell line (112).
Although the cloning of the APE1 gene and characterization of its promoter were performed several years ago, the molecular mechanism(s) for APE1's regulation has still not been completely explored (63, 64). The human APE1 gene appears to have multiple positive and negative regulatory elements. Activation of APE1 expression by signal transducer and activator of transcription-3 (STAT3) in the liver and both redox-dependent and independent function of APE1 has been shown for protection against Fas-mediated liver injury (59). Earlier, our laboratory identified three negative calcium-responsive elements (nCaREs: nCaRE-A, nCaRE-B1, and nCaRE-B2) in the distal APE1 promoter (72). Two such elements (nCaRE-A and nCaRE-B) were originally identified in the promoter of PTH gene which is negatively regulated by extracellular calcium and APE1 was identified as one of the trans-acting factors (107). These observations strongly suggest that APE1 may negatively regulate its own expression by binding to these nCaREs, in particular to the nCaRE-B2.
We have recently shown that wild-type (WT) p53 but not mutant p53 negatively regulates APE1 expression in colon carcinoma HCT116 cells (159). Ectopic overexpression of WT p53 but not mutant p53 (V143A, L22G, and T23S) significantly decreased APE1-promoter-dependent luciferase expression in unstressed p53-null HCT116 cells (159). Furthermore, we have shown decrease in the levels of both APE1 mRNA and protein after camptothecin treatment in HCT116 p53(+/+) cells, but not in the isogeneic p53 null cells. Interestingly, the candidate p53-responsive cis element located in the −184 to −143 base pairs (bp) segment of the APE1 promoter lacks a consensus p53-binding site but has one Sp1-binding site (159). However, we showed using chromatin immunoprecipitation (ChIP) assay that p53 was indeed bound to the APE1 promoter, indicating that the APE1 repression by p53 is not mediated via direct cis element binding, but rather involves p53's indirect recruitment to the promoter by other transcription factor (possibly Sp1) (159). Our findings are intriguing because p53 acts as a pro-apoptotic factor in the cellular response to stress, whereas APE1 is a pro-survival protein whose DNA repair function is essential for protecting cells from oxidative DNA damage. It appears that the cell uses p53 to downregulate APE1 expression in response to DNA damage, and promotes apoptosis. Consistent with this idea, several earlier studies showed reduction of APE1 expression during apoptosis in neurons after transient global cerebral ischemia in rats (34, 50, 90, 143). Additional studies are needed to unravel the physiological significance of such downregulation.
Another factor that modulates APE1 expression is hypoxia, which mimics oxygen tension that is encountered by cells in tissues in vivo. Hypoxia induces APE1 mRNA and protein levels in HT29 cells (158). Elevation of APE1 steady-state mRNA levels is an early event following hypoxia, and persists after restoration of cells to normoxia (158). Nuclear run-on analysis demonstrated that induction of transcription is responsible for elevation of APE1 mRNA (158). Changes in APE1 expression in response to hypoxia was correlated with its requirement for enhanced AP-1 binding following hypoxia via redox activation (158). However, another possible role for prolonged expression of APE1 following hypoxia relates to DNA repair function that remains to be elucidated. Although it is not known whether hypoxia-inducible factors (HIFs) bind-specifically to the APE1 promoter or enhancer, APE1 regulates HIF-1α functions in vivo (36, 69). APE1 up-regulation significantly potentiates hypoxia-induced expression of a reporter construct containing the HIF-1α-binding site (36). Moreover, Ema et al. (36) and Carrero et al. (17) showed that APE1 is critical to linking coactivator proteins, CBP/p300 and SRC-1 to HIF-1α. In contrast, Hall et al. showed that hypoxia downregulates APE1 protein level in both calf pulmonary artery endothelial (CPAEC) and human umbilical vein endothelial (HUVEC) cells (60). Such hypoxia-induced decrease of APE1 was associated with significant induction of apoptosis in CPAEC and HUVEC cells (60). Thus, APE1 downregulation may be permissive in promoting apoptosis in endothelial cells in response to hypoxia. Indeed, APE 1 overexpression was shown to protect CPAEC cells from hypoxia-induced apoptosis (60).
Recently, it has been shown that soy isoflavones down-regulate expression of APE1 in PC3 prostate cancer cells (119). Moreover, pretreatment with soy isoflavones inhibits radiation-induced APE1 expression and activation of NF-κB. Although the mechanism by which soy isoflavones down-regulates APE1 expression is not known, downregulation of APE1 and inhibition of NF-κB activation by soy isoflavones was shown to inhibit tumor growth in vivo (119).
Redox Activation Function of APE1
The regulation of protein functions via redox reaction is becoming increasingly evident not only for transcriptional regulation by trans-acting factors but also for other activities such as that of protein phosphatases (23, 113). APE1 enhances DNA-binding of AP-1 via reduction of the conserved Cys272 residue in the DNA-binding domain of c-Jun (149, 150). Oxidation of APE1 significantly diminishes its ability to stimulate AP-1 DNA-binding activity (149). The reduced Ref-1 activity of oxidized APE1 can be restored by treatment with thioredoxin (TRX), suggesting that a redox–sensitive Cys in APE1 is involved in the reductive activation of the AP-1 complex (67). Ionizing radiation induced nuclear translocation of thioredoxin and its stable interaction with APE1, leading to subsequent activation of AP-1 indicated a thiol exchange cycle APE1's Cys reduces oxidized TFs and is subsequently reduced by TRX (67, 138, 146). A large number of stress-inducible transcription factors including NF-κB (101), ATF/CREB (150), p53 (75), Pax5 (135, 136), Pax8 (16, 133), c-Myb (150), HIF1-α (69), and others have been reported to be activated by APE1-mediated redox activation, presumably redox mechanism involving the same Cys residue. Analysis of several deletion mutants of APE1 revealed that N-terminal 1-127 amino acids (aa) residues is sufficient for its redox activation of c-Jun binding in vitro and repair and redox activities of APE1 are encoded by two distinct regions of APE1 (151) (Fig. 1B). Interestingly, N-terminal 61 amino acids residues which are not conserved in E. coli and yeast APN1 are essential for APE1's activation of c-Jun in vitro (142). Site-directed mutagenesis analysis identified Cys65 as the redox active site in APE1 (142). In addition, it was proposed that Cys93 interacts with the Cys65, presumably via intramolecular S–S bridge formation, and is also involved in redox regulation of AP-1 (142). X-ray structure of hAPE1 lacking 40 aa residues, solved by Mol et al. (99) with our collaboration, show Cys65 itself is buried and not easily accessible by other proteins. Moreover, the distance between Cys65 and Cys93 is too far to form a S–S bond unless there is a conformational change occurring upon oxidation (Fig. 2). The human APE1 has seven conserved Cys residues at position 65, 93, 99, 138, 208, 296, and 310 (Fig. 2). The x-ray crystallographic structure suggests that Cys138 located on the surface is easily accessible for oxidation/reduction (Fig. 2). Furthermore, whether the oxidation product of APE1 is intramolecular S–S or sulfenic acid (SOH) has not been established. Reversible oxidation of Cys to intra or intermolecular S–S from two Cys residues is more widely proposed than the single Cys-SH oxidation to Cys-SOH (23, 26, 114). Both SOH and S–S are reduced by reductases, but the key difference is that the single residue SOH does not require a second, appropriately positioned Cys for S–S bridge formation. H2O2 generates Cys-SOH when the pKa of Cys-SH is lowered for ionization in the proximity of basic residues (137). Interestingly, the His–Cys sequence in protein tyrosine phosphatases, which strongly enhances Cys reactivity (91), is also present in APE1 (H309C310).
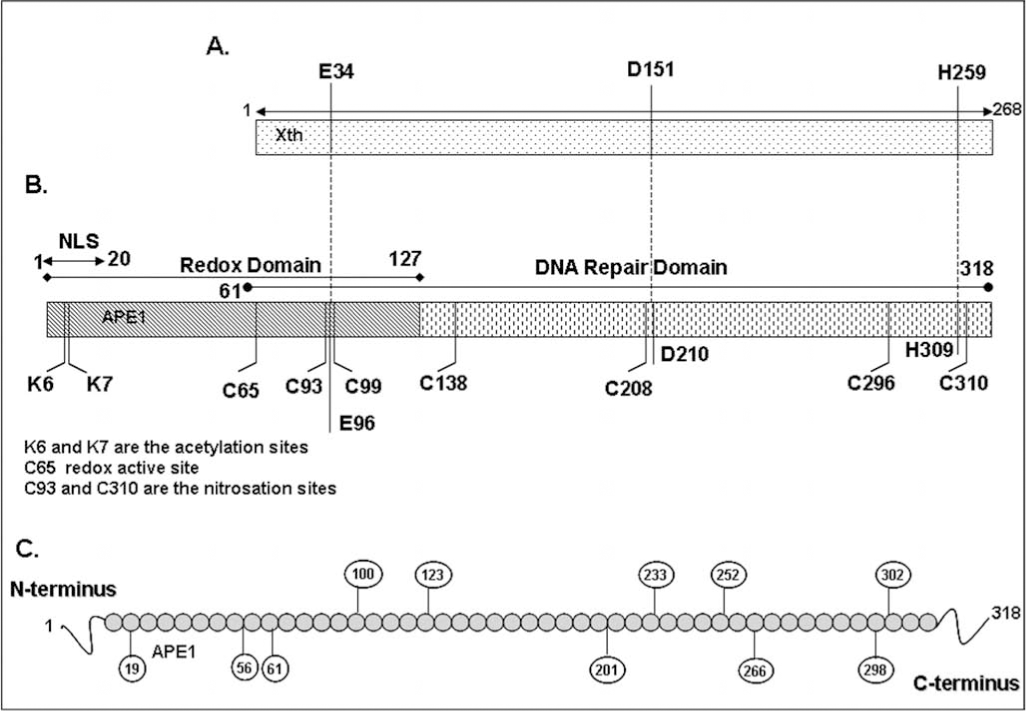

The original suggestion of Cys65 as the active site was supported by the NMR evidence for TRX interaction with Cys65-containing polypeptide (116). Seo et al. (124) showed that the Cys65 mutant acts as a dominant negative inhibitor of the redox function of selenomethionine-containing APE1. Furthermore, apoptosis of neurons after oxidative stress is prevented by ectopic WT APE1 but to a smaller extent by the Cys65Ala mutant (139). Overexpression of either repair defective His309 or Cys65 protected cells to a similar extent upon H2O2 treatment, indicating that both repair function and redox active Cys65 are involved in protecting the cells from oxidative damage (42). Moreover, several studies using transcription factor-dependent reporter expression assays showed that the Cys65 mutant does not behave like the WT APE1 (76, 111, 162). However, this Cys65-mediated proposed redox function of APE1 is recently been challenged by Curran's group (110) who showed that homozygous APE1 Cys64Ala (in human Cys65) knock-in mouse mutants are viable and retain normal levels of AP-1 DNA binding activity. Moreover, the recombinant Cys64Ala APE1 mutant showed normal c-Jun reducing activity like WT APE1 in vitro (110). Taken together, these studies confirm that the APE1 Cys64/65 is not essential for APE1's redox regulatory function. It is thus possible that a second Cys residue could also serve as the redox active residue. Alternatively, either Cys65/64 or Cys93 (both included in the redox active domain of APE1) could serve as a redox active site in the absence of the other. However, given the location of these two Cys in the proposed structure of the APE1 (Fig. 2), we must also consider that neither of the residues is involved in its redox function. However, a recent study showed that small molecule inhibitor 3-[5-(2, 3-dimethoxy-6-methyl-1,4-benzo-quinoyl)]-2 nonyl-2-propionic acid (E3330) specifically inhibits APE1's redox function in ovarian cancer cells (93). Moreover, E3330 also inhibits mouse retinal vascular endothelial cells proliferation and angiogenesis in vitro. Very recently, Ando et al. (3) showed that APE1 stimulates p50 or c-Jun DNA-binding by facilitating their reduction by reducing agents such as glutathione or thioredoxin, and none of the APE1's Cys residues is essential for this activity.
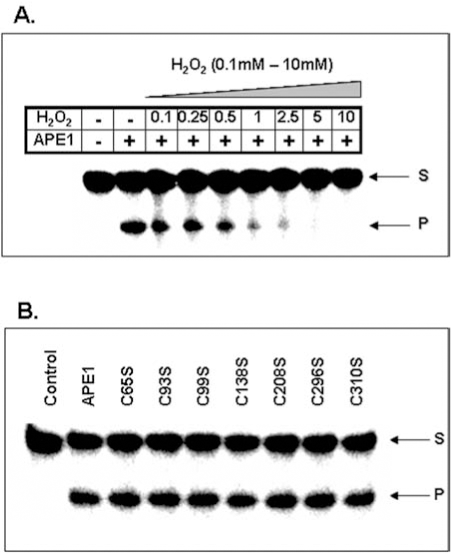
Despite the formation of dimeric APE1 in vitro, there is no strong evidence for favoring S–S formation over S-OH during oxidation of APE1, because the second Cys-SH in the S–S bond has not been identified (53). However, our data show that oxidation of recombinant APE1 with H2O2 inhibits it AP-endonuclease activity in a dose-dependent manner (Fig. 3A). Interestingly, Kelley's group suggested that oxidation of Cys310 residues also affects APE1 DNA repair activity (81). Treatment with 10 mM H2O2 or 3 mM diamide completely abolished DNA repair activity, while high dithiotheritol (DTT) level restored the activity (81). We have shown that none of the Cys→Ser mutation affected APE1's endonuclease activity in low (< 2 mM) Mg2+ (Fig. 3B). However, the Ser99 mutant behaved uniquely because of strong inhibition of its enzymatic activity in higher (10 mM) Mg2+ that is commonly used is activity measurements (94). Furthermore, unlike WT and other Cys→Ser mutants, the Ser99 mutant did not bind to DNA as judged by electrophoretic mobility shift analysis (94). The unexpected involvement of Cys99 (∼16A° from the active site) in APE1's substrate binding and catalysis provides an example of involvement of a residue far from the active site.
Covalent Modifications of APE1
Post-translational modifications of proteins have emerged as the main mechanisms for the control of many biological functions, including signal transduction, gene expression, protein degradation, nucleoplasmic shuttling, and protein–protein interaction (51). It is evident that the activity of a single protein with pleiotropic functions could be “finely tuned” via post-translational modifications, including phosphorylation, acetylation, nitrosation, ubiquitination, and methylation, etc., in order to coordinate specific biological activities. Given the multiple functions, it is not surprising that APE1 is post-translationally modified in vivo. An earlier study by Yacoub et al. (154) showed that APE1 can be phosphorylated in vitro by casein kinase I and II (CKI and CKII). APE1 contains a number of potential phosphorylation sites including consensus sequences for CKI, CKII, and protein kinase C (PKC) (Fig. 1C). CKII-mediated phosphorylation of APE1 abolished DNA repair activity in vitro, while phosphorylation by CKI or PKC has no effect (154). In contrast, a subsequent study by Fritz and Kaina (43) showed that APE1 phosphorylation by CKII enhances redox activation of the AP-1 transcription factor and has no effect on its DNA repair activity. At this time, the reason for these discrepancies is unclear, whereas unequivocal evidence for of APE1's in vivo phosphorylation and identification of phosphorylation sites are clearly warranted. The first in vivo evidence for PKC-mediated phosphorylation of APE1 was shown by Hsieh et al. (68). Although immunoprecipitated APE1 from phorbol 12–myristate 13-acetate (PMA) or hypochlorite treated cell extracts showed enhanced DNA-binding of AP-1 presumably by PKC-mediated phosphorylation of APE1 (68), this study did not identify the specific PKC phosphorylation site nor did address whether APE1's endonuclease activity is also affected by PKC phosphorylation. Thus, an attractive hypothesis is that exposure of cells to genotoixc stress leads to phosphorylation-dependent stimulation of another post-translational modification (which we are currently investigating); this in turn modulates APE1's redox activity for activation of stress-inducible transcription factors such as AP-1, NF-kB, and p53. Since different phosphorylation sites of APE1 have not been functionally characterized in vivo by site-directed mutagenesis, the biological significance of APE1's phosphorylation is still unclear.
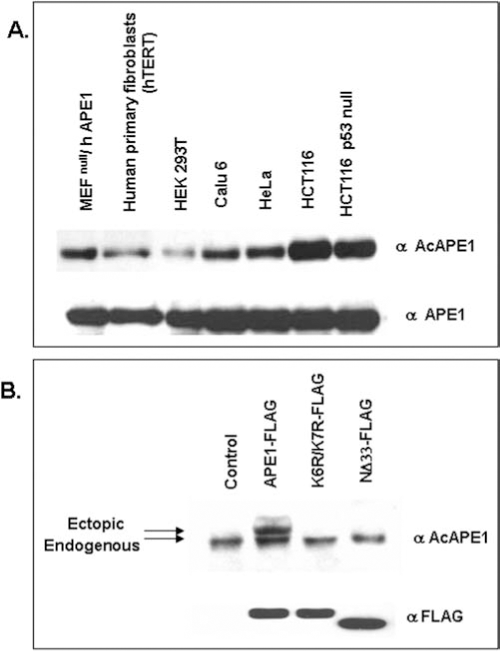
We have earlier established APE1's acetylation at Lys6 and Lys7 by the histone acetyltransferase p300 both in vivo and in vitro (10, 12). Our in vitro studies with recombinant WT APE1 or K6R/K7R or K6L/K7L with p300 identified the acetyl-acceptor residues as Lys6 and Lys7 (10). However mass spectroscopic analysis of in vitro acetylated APE1 (AcAPE1) could not detect diacetylated APE1 that could be explained by the possible steric effects of acetyl groups attached to ɛ-amino groups during second acetylation (10). Our data thus indicate that either Lys6 or Lys7 but not both can be acetylated in the same molecule. It is interesting to note that Lys6 and Lys7 are conserved in most mammalian APE1 including human, mouse, and bovine, however Lys7 is not conserved in rat and Chinese hamster, suggesting that acetylation of Lys6 has specific biological function in mammals (Fig. 4). We prepared affinity-purified AcAPE1-specific antibody using a human APE1 peptide with acetylated Lys6 and showed that the AcAPE1 antibodies are highly specific for AcAPE1, and do not cross react with at least 25-fold excess unmodified APE1 as judged by immunoblotting (38, unpublished data). Moreover, this antibody recognizes ectopic FLAG-tagged WT APE1, but not nonacetylable K6R/K7R APE1 in cell extracts confirming its specificity for AcAPE1 (Fig. 5B). Using this antibody we have provided unequivocal evidence for the presence of endogenous AcAPE1 under normal conditions (Fig. 5A).

Although the first 20 amino acid residues contain a consensus nuclear localization signal (NLS) sequence (MPKRGKK) that includes the acetylation sites (Fig. 1B), we have shown that acetylation of K6 and K7 of APE1 is not involved in its nuclear localization (74, 129). Although lysine residues are targets for both acetylation and ubiquitination, and acetylation may protect a protein from ubiquitination and degradation (55, 77), both WT and nonacetylable K6R/K7R APE1 mutant showed similar in vivo stability (74). We have shown earlier that acetylation of other DNA repair proteins involved in BER modulates their activity. For example, acetylation of K338 and K341 of 8-oxoguanine-DNA-glycosylase (OGG1) enhances its catalytic activity while acetylation of NEIL2 at the conserved K52 abolished its DNA glycosylase activity (9, 11). In contrast, acetylation of APE1 does not change its AP-endonuclease (DNA repair) activity, but rather modulates its transcriptional regulatory functions (10). We showed earlier that APE1 acetylation stimulates the formation of nCaRE-B complex which also contains hnRNP-L and HDAC1 (10). It appears likely that acetylation-mediated conformational change in the disordered N-terminal segment (encompassing 40 aa residues) which is dispensable for APE1's endonuclease activity, modulates protein–protein interaction. Our recent data indicate that APE1 stably interacts with YB-1 and acetylation enhances its binding with YB-1 both in vivo and in vitro (unpublished data).
Recently, S-nitrosation of APE1, another post-translational modification, has been shown to occur in vivo. Treatment of S-nitroglutathione, an S-nitrosating agent, stimulated nuclear export of APE1 through S-nitrosation of Cys93 and Cys310 in a CRM1-independent manner (117). Immunofluorescence studies using HA-tagged APE1 showed that this translocation process is dependent on Cys93 and Cys310 residues and can be reversed by reductive reagents or antioxidant, however could not be mimicked by oxidative stress (117). Although the precise mechanism by which S-nitrosation controls nuclear export of APE1 is not clear, aa residues 64–80 (close to Cy 93) were found to be critical for the nitrosation-mediated cytosolic translocation of APE1 (117). Moreover, inhibition of this translocation after overexpression of histone deacetylase HDAC2 raised the possibility that acetylation of APE1 (because we have shown that APE1 stably interacts with HDAC2) may cross talk with nitrosation in this process (12, 117). Recently Hara et al. (62) reported S-nitrosation-mediated nuclear translocation of glyceraldehyde 3 phosphate dehydrogenase (GAPDH). Although our earlier studies suggested that APE1 acetylation is not relevant to its nuclear localization, the possible role of acetylation in nuclear export can not be ruled out. Although there is no experimental evidence for APE1's ubiquitylation or sumoylation, its interaction with the ubiquitin conjugating enzyme ubc9 raises the possibility that APE1 may undergo turnover through these modifications (157).
APE1 as a Transcription Factor
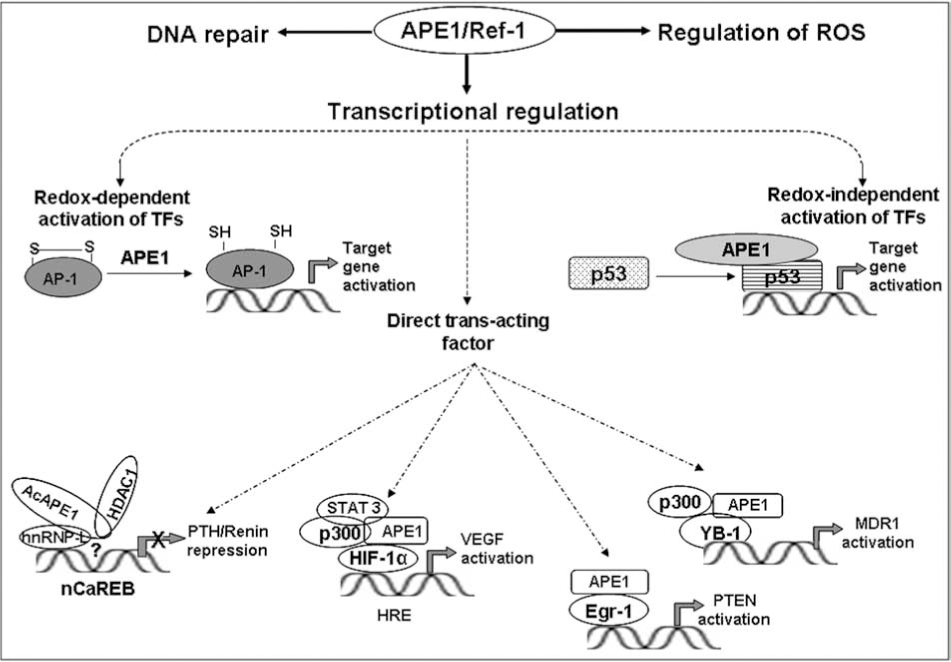
APE1 as a trans-acting factor was discovered by Okazaki's group when they identified APE1/Ref-1 as one of the proteins that binds to negative calcium response elements (nCaRE) complex in the human PTH gene promoter (107). In most cell types, rise in intracellular [Ca2+]i triggers activation of many genes' expression and secretion, with the notable exception of parathyroid cells and renin-producing juxtaglomerular cells, where a rise in [Ca2+]i suppresses both renin and PTH gene expression (58, 155). This is mediated by the binding of cognate trans-acting factors to nCaRE in the PTH and renin promoter (107). Two such elements (nCaRE-A and nCaRE-B) in the PTH gene promoter were identified and APE1 was found to be associated with these elements (107). Subsequently, Macfee et al. identified putative nCaREs in some 100 genes, including several genes including calmodulin and β-myosin, that are subject to Ca2+-mediated regulation (95). An nCaRE-B sequence, identical to that in the PTH promoter, at 1.8 kb upstream of the transcription start site in the human renin gene has been identified (44). Involvement of this element in Ca2+-mediated repression of renin promoter-driven reporter expression has been shown in chorio-decidual cells (44). Again, APE1 was identified as a component in the nCaRE-B bound-complex, analogous to the situation in the PTH gene, and calcium-induced nuclear translocation of APE1 was also observed (44). However, APE1 does not directly bind to the nCaRE-B, indicating a requirement for additional factors in the trans-acting complex assembly. The Ku70 (Ku86) proteins were identified in the complex bound to nCaRE-A (21). Subsequently, heterogeneous ribonucleoprotein L (hnRNP-L) and APE1 were identified as major components of the complex bound to nCaRE-B (88). We showed earlier that AcAPE1 does not bind alone to nCaRE-B, although acetylation of APE1 enhances the binding of nuclear extracts to nCaRE-B (10). An increase in extracellular Ca2+ increases the level of acetylated APE1 due to Ca2+-induced activation of p300 HAT activity, presumably by inducing phosphorylation of p300 catalyzed by protein kinase C or Ca2+-dependent calmodulin (CAM) kinase IV (1). Moreover, using ChIP assay, we showed that acetylation of APE1 enhances recruitment of APE1-HDACs complexes to the PTH promoter in a Ca2+-dependent manner and AcAPE1 acts as corepressor. However, the detailed molecular mechanism by which APE1 represses renin gene expression by binding to nCaRE-B is still unclear.
An earlier study showed that oxidative stress-induced activation of APE1 increases Egr-1's DNA-binding activity in osteoblastic HOBIT cells (111). Egr-1, a transcription factor with tumor suppressor function, regulates expression of many genes, including p53 and PTEN, that control cell growth arrest and apoptosis (7, 140). Coimmunoprecipitation studies showed that APE1 stably interacts with Egr-1 and treatment with H2O2 strongly stimulates their association (111). Recently, Tell's group in collaboration with us has shown that Egr-1-mediated activation of PTEN expression is dependent on APE1 acetylation (39). Treatment of HeLa cells with HDAC inhibitors was further shown to increase APE1 acetylation and induced PTEN expression (39). The absence of such induction in APE1 downregulated HeLa cells confirmed APE1's role in regulating inducible PTEN expression. Moreover, activation of Egr-1-dependent PTEN promoter due to overexpression of WT APE1, but not of the nonacetylable K6R/K7R mutant further confirms that acetylation-enhanced direct transcriptional activity of APE1 in Egr-1-dependent PTEN expression (39).
Stable interaction of APE1 with HlF1-α, STAT3, and CBP/p300 in hypoxia-induced expression of vascular endothelial growth factor (VEGF) further suggests a role for APE's direct regulatory role in VEGF expression (54, 161). APE1's interaction with HIF-1α and p300 (but not with ATF/CREB) and its presence was found to be critical for assembly of the hypoxia-inducible transcriptional complex on the hypoxic response element (HRE) in the VEGF gene promoter in the rat pulmonary artery endothelial cells (PAEC) (161). Moreover, immunodepletion of APE1 prevented association of HIF-1 with p300, ATF, and CREB, indicating the critical role of APE1 in complex formation (161). Supporting this, a recent study Gray et al. (54) showed that APE1 stably interacts with STAT3 and HIF-1α, and STAT3, CBP/p300, and APE1 are components of a transcriptional complex that regulates Src-dependent hypoxia-induced expression of VEGF in pancreatic and prostate carcinomas (54). STAT3 and HIF-1α bind simultaneously to the VEGF promoter, where they form a complex with the transcription coactivators CBP/p300 and APE1. In addition to enhancing DNA affinity of transcription factors, APE1 may target recruitment of p300 to the transcription complexes (54) which suggests that APE1 acts as a direct transcriptional coactivator in hypoxia-induced VEGF expression. Why would APE1, a DNA repair protein, with redox function, also be present as a stable component of both constitutive and hypoxia-inducible transcription? In this regard it is worth noting that p300, best known as a scaffold protein that facilitates transcriptional complex assembly, and is also involved in chromatin remodeling via its intrinsic HAT activity (105), also acetylates Lys6 (or Lys7) in APE1. Thus, p300-catalyzed acetylation of APE1, perhaps in concert with other proteins, may be the key to incorporation of APE1 in the constitutive and hypoxia-inducible transcriptional complex. Additional studies will be required to settle this issue.
Recently, using affinity screening, we identified other interacting partners of APE1 including Y-box-binding protein YB-1 that regulates many genes positively or negatively (84, 106). APE1 stably interacts with YB-1 and its acetylation enhances its binding to YB-1, leading to activation of the Y-box-dependent MDR1 promoter (unpublished data). Mutation of Lys6 and Lys7 significantly decreased the binding. The exact mechanism by which acetylation of APE1 activates MDR1 expression is not clear. The likely redox-inactive Cys65Ser and Cys138Ser APE1 mutants behaved like WT APE1 in modulating YB-1-mediated MDR1 promoter activity, suggesting that APE1's redox activity is not involved in MDR1 activation. Using the ChIP assay, we have provided direct evidence that APE1 is present with the Y-box-bound complex in the MDR1 promoter in vivo, and that tricostatin A (TSA, a specific HDAC inhibitor) treatment enhanced (presumably via increased APE1 acetylation) MDR1's promoter occupancy (unpublished data).
Redox-independent transcriptional function of APE1 in modulating HIF-1α, Egr-1, and YB-1 activities is not without precedent. Both redox-dependent and redox-independent transcriptional function of APE1 in modulating p53-DNA-binding was first described by Jayaraman et al. (75). Recombinant APE1 was shown to stimulate DNA-binding of full length p53, which was further strongly stimulated in the presence of DTT, indicating that APE1-mediated activation of P53 might be independent of its function as a redox activator. This was further supported by the observation that while APE1 stimulated binding of full length p53 in the presence of DTT, it was incapable of stimulating binding of p53 deletion mutant (p53Δ30), which lacked the carboxy terminal 30 amino acid residues, suggesting that redox-independent activation of p53 is due to interaction of APE1 with its C-terminal region. This was confirmed in vivo by the observation that APE1 overexpression stimulated transactivation function of full length p53 in a reporter-based assay but not the p53Δ30 mutant (75). Although stable interaction between p53 and APE1 could not be shown, Far Western and coimmunoprecipitation Western assay suggested weak interaction between these proteins (49). It is thus tempting to speculate that APE1 has dual functions with respect to p53 binding: binding to the C-terminus abolishes negative regulation, thereby making the protein accessible to redox regulation. Recently, Hanson et al. (61) showed that APE1 promotes association of p53 dimers into tetramers, and destacking of higher oligomeric forms into tetrameric form in vitro, thereby enhancing p53 binding to target DNA. This novel assembly function of APE1 in promoting tetramer formation of p53 was proposed to be the underlying mechanism for it redox-independent effects on p53.
Although APE1 was identified in diverse trans-acting complexes, APE1 by itself has no affinity for any specific cis element; rather it affects promoter activity by binding to trans-acting factors specific for distinct cis elements. Thus APE's presence in diverse trans-acting complexes requires interaction with diverse partners (Fig. 6). In the case of nCaRE-B, we identified hnRNP-L and HDAC1. Similarly for HRE-binding, APE1 interacts with HIF-1α, STAT3, and p300, and with YB-1 and p300 for Y-box response element (YRE) dependent regulation (Fig. 6). However, how APE1 acts as a coactivator or corepressor in such diverse complexes is not clear.
AcAPE1 as a Cancer Therapeutic Target
APE1 overexpression, often observed in tumor cells, is associated with resistance to various anticancer drugs (13, 78, 80, 86, 115, 121, 153); its downregulation sensitizes tumor cells to such agents (14, 122, 144). Several studies have shown that targeted reduction of APE1 protein by specific anti-sense oligo or siRNA sensitized to MMS, H2O2, bleomycin, temozolomide (TMZ), and gemcitabine (14, 89, 109, 141). Whether such enhanced sensitivity is solely due to the loss of APE1's DNA repair activity or also due to the loss of its transcriptional regulatory function or both is still unknown. Because APE1 plays a central role in the repair of endogenous DNA damage and small base adducts induced by alkylating agents via the BER pathway, it is not surprising that APE1 overexpression is associated with tumor cells' resistance to alkylating drugs. However, downregulation of APE1 was shown to sensitize tumor cells to etoposide or cisplatin (86, 153), which cannot be easily explained by the repair function of APE1 as these drugs induce DNA double-strand breaks or DNA intra-strand cross-links that are repaired via the APE1-independent nonhomologous end joining or nucleotide excision repair pathways. As we mentioned earlier, APE1 acetylation enhances its YB-1 binding and activates MDR1 gene expression (unpublished data). Tumor cells frequently develop drug resistance after initial chemotherapy due to elevated level of MDR1 which is often upregulated due to promoter activation after drug treatment (19, 85). Immunostaining analyses of various human cancers including breast cancer and osteosarcoma showed that the elevated nuclear level of YB-1 is closely associated with the acquisition of MDR1-mediated multidrug resistance (6, 45, 104). We have demonstrated that downregulation of APE1 sensitizes MDR1-overexpressing tumor cells to anticancer drugs such as doxorubicin and cisplatin, which suggests a molecular basis by which APE1's transcriptional function and YB-1 could be linked to induce drug resistance of tumor cells. This was further supported by our observation showing a significant correlation between APE1 and MDR1 levels in many lung cancer tissues (unpublished data). Thus, identification of small molecules that blocks p300-mediated APE1 acetylation or inhibition of interaction of AcAPE1 with YB-1 or other transcription factor could be used as potential therapeutic targets for cancer drug sensitization.
APE1 in Neurodegenerative and Cardiovascular Diseases
Ischemia and oxidative stress induced by spinal cord injury (SCI) are known to be associated with death of neuronal cells. Moreover, chronic oxidative stress and DNA damage have been implicated in several neurodegenerative diseases, including Parkinson's (PD), Alzheimer's (AD), Huntington's (HD), and amyotrophic lateral sclerosis (ALS) (8, 22). Isobaric hyperoxia (100% oxygen) stimulated APE1 expression in the hippocampus and basal forebrain of young rats, while aged rats showed no significant changes in APE1 protein levels in all brain areas after hyperoxia (35, 118). In contrast, several studies showed that global cerebral ischemia or traumatic brain injury or cold injury-induced brain trauma (100) induced oxidative stress decreases APE1 expression in the hippocampus and is associated with neuronal apoptosis in rats (34, 50, 143). This specific inhibition of APE1 expression may affect the extent of apoptosis after ischemia. Consistent with this, overexpression of WT APE1 in hippocampal and sensory cells reduced neuronal death after H2O2-mediated oxidative stress (139). C65A repair competent/redox incompetent APE1 conferred only partial cell survival, suggesting redox and repair functions of APE1 are essential for neuronal survival. In spinal cord injury, APE1 expression was noted to be decreased making neurons susceptible to oxidative DNA damage. APE1 is differentially expressed in cells of the nervous system (32, 33). In AD brain, APE1 has been visualized in a subpopulation of senile plaques by immunolocalization, indicating a functional relationship between APE1 and amyloid beta (Aβ) protein, possibly a pivotal role for APE1 in Aβ accumulation (130). APE1 expression was observed to be increased in nuclear extracts of AD patients compared to controls (24). Increased APE1 immunoreactivity was further supported from the hippocampus of AD brains in areas of neuronal injury and plaque-like structures (130). ALS is characterized by degeneration of motor neurons. APE1 expression and activity was observed to be lower in patients with sporadic ALS (83). In contrast, an increase in APE1 was also reported in the brain and spinal cord samples of patients with ALS (125), and reduced DNA repair activity of APE1 protein in familial ALS patients (108) has been shown. It was proposed that action of inducible transcriptional factors such as c-Jun in mammalian neurons is likely to be regulated by constitutively expressed APE1 and protecting neuronal cells after brain injury and oxidative DNA damage.
Although APE1 null mice are embryonic lethal, partial loss of APE1 in the heterozygous mice (APE1+/−) causes cardiovascular diseases such as hypertension (76). Jeon et al. (76) made a remarkable observation that APE1+/− mice have markedly elevated blood pressure, clearly indicating APE1's important role in maintaining blood pressure. This phenotype is accompanied by decreased basal nitric oxide (NO) production in arteries from APE1+/− mice (76). However, expression of endothelial nitric oxide synthase (eNOS) cannot be a major determinant of the hypertensive phenotype of APE1+/− mice, because eNOS levels are actually increased in the vessels of these mice, despite their impaired ability to generate nitric oxide. The redox activator function of APE1 was shown to be involved in moderate upregulation of H-Ras expression and activation of AKT, which potently activates eNOS (76). Therefore, it is critical to emphasize that APE1 may coordinate multiple nuclear events that impact vascular reactivity and blood pressure control through multiple mechanisms in addition to its effects on NO generation. It is too soon to know exactly how APE1 contributes to regulation of blood pressure, in addition to the effects on nitric oxide synthase activity that have been elegantly defined by Irani and colleagues (76). However, it is tempting to speculate that APE1 may play an integrative role in maintaining vascular tone through regulation of master control genes such as nitric oxide synthase, renin, and perhaps others that tune vascular responses.
Vascular endothelial cells can undergo apoptosis in vitro in response to variety of pathophysiological conditions such as hypoxia and pro-inflammatory cytokine (28, 145). Hall et al. (60) showed that decreased APE expression was associated with hypoxia-induced apoptosis in HUVEC and CPACE cells (60), which were significantly inhibited by overexpression of APE1. Deletion of the redox-sensitive domain of APE1 (aa 1–127 residues) abolished the anti-apoptotic function of APE1 in both hypoxia and tumor necrosis factor α (TNF-α) induced endothelial cell death (60). This study suggests that up-regulation of APE1 promotes endothelial cell survival in response to hypoxia and TNF-α through NF-κB-independent and NF-κB-dependent signaling cascades. Interestingly, a recent study shows that APE1-mediated NF-κB nuclear translocation is responsible for lipopolysaccharide-stimulated inducible NO synthase (iNOS) expression in a murine macrophage cell line (127). In contrast, Angkeow et al. (4) showed that cytoplasmic APE1 translocates to the nucleus upon oxidative stress induced by hypoxia/reoxygenation (H/R) in HUVEC cells. APE1 overexpression suppressed H/R-induced oxidative stress, NF-κB activation, and apoptosis (4). It was proposed that cytoplasmic APE1 inhibits oxidative stress (H2O2) generation by inhibiting the rac GTpase-activity and thus prevents H2O2 generation and NF-κB activation (4, 107). This novel cytoplasmic role of APE1 in preventing oxidative stress and apoptosis indicates that both nuclear and cytoplasmic function of APE1 may be important for preventing apoptosis in endothelial cells.
Expression of adhesion molecules accelerates the adhesion and migration of monocytes towards sites of inflammation in response to a variety of stimuli and has been implicated in atherosclerosis (123). A recent study by Kim et al. (82) showed that APE1 overexpression inhibits TNF-α induced VCAM expression in HUVEC cells. Moreover, overexpression of APE1 suppressed monocytes cells (U937) adhesion to HUVEC cells after TNF-α stimulation (82). Moreover, this anti-adhesive property of APE1 is mediated by a NOS-dependent mechanism. It was shown that APE1 may inhibit VCAM by inhibiting superoxide production and p38 MAPK activation (82).
Concluding Remarks and Future Perspective
Our and others studies have confirmed that APE1's DNA repair function is essential for somatic cell survival in mammals. However, the essentiality of APE1's acetylation dependent (redox-independent) transcriptional activity was quite unexpected. Moreover, the observation that the Cys65 residue of APE1 is not essential for redox regulation of AP-1 DNA binding indicated that the exact molecular mechanism underlying the redox activity of APE1 remains unclear. Therefore, definitive identification of redox active Cys residue of APE1 in vivo is essential to settle this issue. Moreover, whether post-translational modification such as phosphorylation, acetylation, or nitrosation is needed for maintaining APE1's redox activation function in vivo is not known. APE1 has been found to stimulate transcriptional activity of numerous factors involved in diverse physiological functions such as cell cycle control, apoptosis, drug resistance, angiogenesis, cellular growth, hematopoesis, and cardiovascular functions (40, 41, 47, 70, 76, 82, 121, 122, 144, 162). It is not clear whether APE1 acts as a direct or indirect sensor in all of these cases for modulating cellular functions. Furthermore, stable interaction of APE1 with several transcription factors and its presence in several trans-acting complexes have raised the possibility of its redox-independent transcriptional functions. We have provided evidence that suggest acetylation of APE1 modulates its coactivator or corespressor functions. However, how APE1 acts as a coactivator or corepressor in such diverse complexes is not clear. One of the major challenges is to elucidate the molecular basis for redox-independent trans-acting activity of APE1 in diverse complexes responsive to repression, activation of many genes. Moreover, there are many still unanswered questions regarding how different post-translational modifications of APE1 control its subcellular localization or trans-acting functions. Given that frequent overexpression of APE1 is associated with tumor cells resistance to various anticancer drugs, elucidating the signaling mechanisms involved in controlling its expression is extremely important from both basic and clinical perspectives. Moreover, attenuation of the APE1's repair or transcriptional functions (both redox and acetylation-dependent) by small molecule inhibitor(s) is a potential approach for reversing drug resistance. Further studies involving APE1 mutants deficient in DNA repair or redox or acetylation dependent function will settle the issues whether repair or transcriptional function or both are involved in tumor cells resistance to drugs.
Footnotes
Acknowledgments
Research supported by United States Public Health Science grants R01 ES08457, R01 CA53791, and P01 ES06676 (SM), American Heart Association grant 0565008Y (KKB). We thank Drs. T. Izumi (Louisiana State University Health Science Center, New Orleans), N. Oezguen, S. Sengupta (University of Texas Medical Branch, Galveston), R. Chattopadhyay (University of Virginia, Charlottesville) for their help with the figures. We also thank other members of Dr. Mitra's laboratory for their helpful discussions.
Abbreviations
aa, amino acids; AcAPE1, acetylated APE1; AD, Alzheimers disease; ALS, amyotrophic lateral sclerosis; AP, apurinic/apyrimidinic; AP-1, activator protein-1; APE1, AP-endonuclease 1; Aβ, amyloid beta; BER, base excision repair; bp, base pair; CAPEC, calf aortic pulmonary endothelial cell; ChIP, chromatin immunoprecipitation; CHO, Chinese hamster; CKI, casein kinase I; CKII, casein kinase II; DTT, dithiotheritol; Egr-1, early growth response protein-1; E3330, 3-[5-(2, 3-dimethoxy-6-methyl-1,4-benzoquinoyl)]-2nonyl-2-propionic acid; eNOS, endothelial nitric oxide synthase; FEN1, flap endonuclease 1; H2O2, hydrogen peroxide; hAPE1, human AP-endonuclease 1; HAT, histone acetyl-transferase; HDAC1, histone deacetylase 1; HIF-1α, hypoxia-inducible factor-1α; ?hnRNP-L, heterogeneous nuclear ribonucleoprotein L; HOCl, hypochlorous acid; HRE, hypoxia response element; HD, Huntington's disease; HUVEC, human umbilical vein endothelial cells; IL, interleukin; IR, ionizing radiation; mAPE1, mouse AP-endonuclease 1; MD, molecular dynamics; MDR, multidrug resistance; MEF, mouse embryonic fibroblast; MMS, methyl methane sulphonate; nCaRE, negative calcium response element; NEIL, nei-like; NF-kB, nuclear factor-kB; NLS, nuclear localization sequence; NO, nitric oxide; OGG1, 8-oxoguanine DNA glycosylase; PAEC, pulmonary artery endothelial cell; PAX, paired beox containing genes; PCNA, proliferating cell nuclear antigen; PD, Parkinson's disease; PKC, protein kinase C; PMA, phorbol 12-myristate 13 acetate; PTH, parathyroid hormone; Ref-1, redox effector factor-1; ROS, reactive oxygen species; SCI, spinal cord injury; SH, sulfhydral; SOH, sulfenic acid; S–S, disulfide; STAT3, signal transducer and activator protein 3; TFs, transcription factors; TNF-α, tumor necrosis factor alpha; TRX, thioredoxin; TSA, trichostatin A; TSH, thyrotropin; VEGF, vascular endothelial growth factor; WT, wild type; XRCC1, x-ray cross-species complementing 1; YB-1, Y-box-binding protein-1; YRE, Y-box response element.