
Abstract
Management of melanoma is a growing and challenging public health issue requiring novel and multidisciplinary approaches to achieve more efficient prevention and therapeutic benefits. The aim of this article is to show the critical role of APE/Ref-1 on melanomagenesis and progression. APE/Ref-1 serves as a redox-sensitive node of convergence of various signals as well as a DNA-repair enzyme, and its activation protects melanocytes and melanoma cells from chronic oxidative stress and promotes cell survival via mediation of downstream pathways. APE/Ref-1 is a strong candidate as a potential drug-treatable target for the prevention and treatment of human melanoma. Lead compounds exhibiting inhibitory effects on APE/Ref-1 are also reviewed. We anticipate potential clinical benefit in the future through inhibition of APE/Ref-1 and/or Ref-1-mediated signaling. Antioxid. Redox Signal. 11, 639–650.
Introduction
Abnormal Redox Status and Chronic Oxidative Stress is Important in the Pathogenesis of Human Melanoma
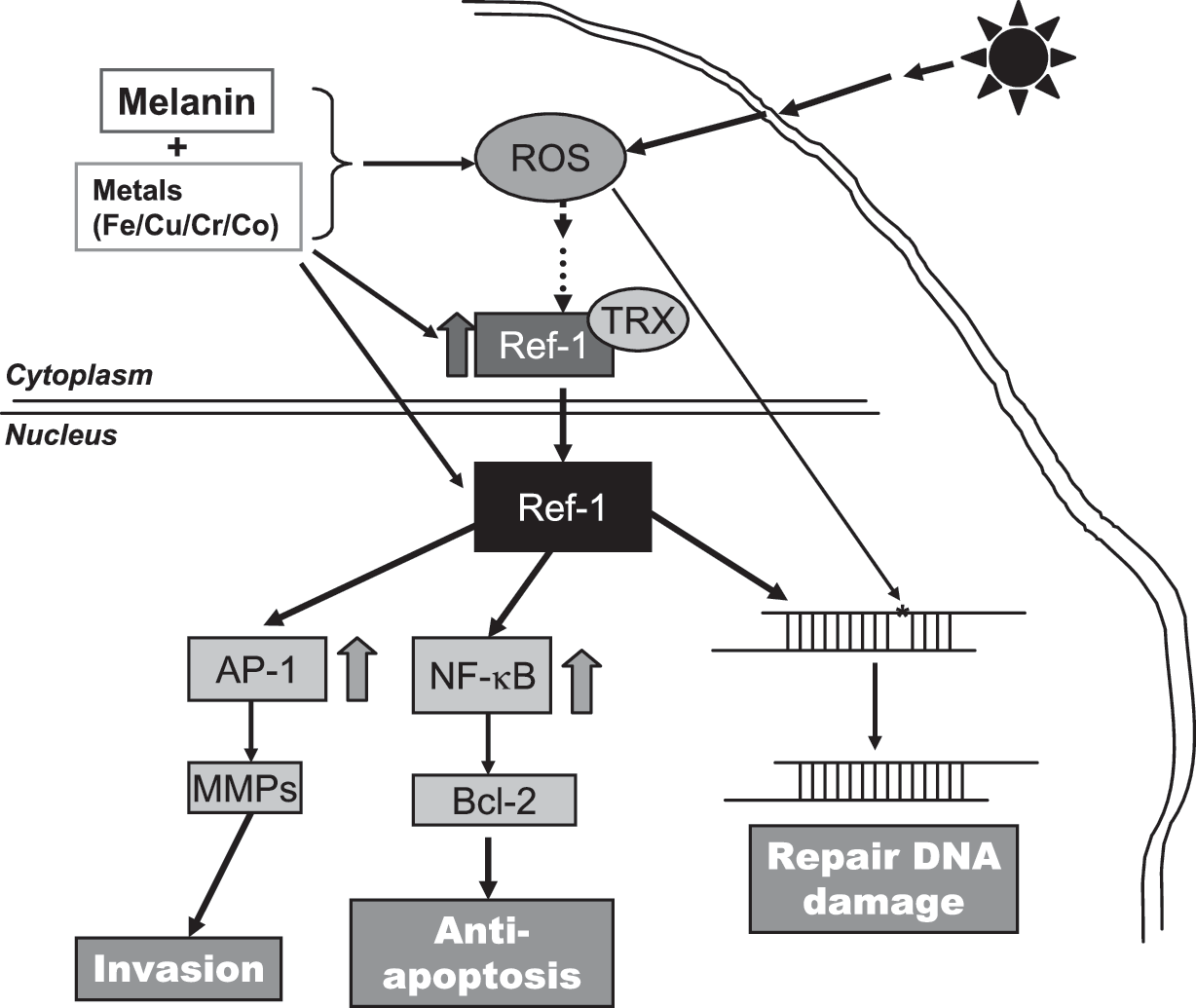
Melanin, a unique product of melanocytes, is the major pigment found in the hair and epidermis, as well as in the brain and other highly nerve-active areas such as the retina and middle ear. The predominant physiologic function of melanin, which is closely linked to its redox potential, is to protect skin from photochemical stress by serving as a disposable buffer by neutralizing reactive oxygen species (ROS) generated by UVR. Melanin synthesis represents a complex series of tightly regulated steps involving the consumption of oxygen and superoxide and the production and utilization of hydrogen peroxide, all carefully orchestrated within a well-organized organelle, the melanosome (37). Melanosomes progressively become more structurally or functionally disordered or both during melanomagenesis, evoking more ROS production and leading to an ongoing peroxide stress within the melanoma cell, especially when bound to Cu (22, 28, 31, 96). Consistently, normal melanocytes efficiently abrogate an exogenous peroxide stress, whereas human melanoma cells are seriously impaired in their ability to do so and generate higher levels of reactive oxygen species (ROS) (74). Contributions of other investigators have also shown that melanoma cells are depleted of cellular antioxidants (85), contain reduced glutathione (GSH) levels (30), and have altered levels of catalase, SOD, and other antioxidant enzymes. These considerations led us to propose a novel model for the etiology and pathogenesis of melanoma (Fig. 1), in which the initial step involved the conversion of melanin from its natural antioxidant/reduced state to that of a superoxide-generating prooxidant compound and the generation of excess (ROS), a process enhanced by the binding to melanin of certain metals found in the environment (28, 75, 76) or represented prominently in the normal melanin-synthesis pathway (i.e., Cu2+ bound to tyrosinase).

In addition to its beneficial health effects, such as vitamin D3 formation, UVR also produces many acute and chronic detrimental cutaneous changes, which are associated with the development of skin malignancies (107). Elevated ROS (i.e., H2O2) and reactive nitrogen species (RNS) levels were evident after UVR (10, 39, 103), which adds an oxidative burden to melanocytes. ROS induces various oxidative DNA damage such as 8-oxo-dihydro-2'-deoxyguanine (8-oxo-dG), which is a mutagenic lesion that is directly repaired by the DNA base excision–repair pathway (35, 70). It is also well-documented that active dietary antioxidants than can scavenge ROS exhibit promising cancer chemopreventive activities (50). For the process of melanomagenesis, extensive epidemiologic observations support the idea that the effects of UVR on the skin are complex (2, 9). A striking feature in melanoma has been the general inability to detect thymine dimers or other classic UVR-induced mutations in primary or metastatic melanomas, even in genes of interest (9), despite convoluted explanations of why this might be so (110).
Increase of APE/Ref-1 as an Adaptive Response to Oxidative Stress
APE/Ref-1 is a node of convergence for various redox-sensitive signals as well as being important in DNA repair. As the major AP endonuclease in human cells, APE/Ref-1 accounts for >95% of the total AP endonuclease activity and is essential for the protection of cells against the toxic effects of several classes of DNA-damaging agents. Recently, many studies demonstrated that many survival, proliferation, or antiapoptosis signalings are activated by APE/Ref-1–mediated transcription factors such as AP-1, NF-κB, and p53, whose regulation occurs in both a redox-dependent and a redox-independent manner (27, 118). It is well documented that elevated APE/Ref-1 is associated with chemo- and radioresistance in a number of cellular systems (11, 77, 102). Knockdown of APE/Ref-1 efficiently induced apoptosis or sensitization or both to chemical treatments in many cancer cells (57, 115, 116, 120).
APE/Ref-1 is uniquely sensitive to both intracellular and extracellular alterations of redox status. ROS not only can inhibit APE/Ref-1 activities by direct oxidation of amino acid residues (49), but also affects the expression level and subcellular localization of APE/Ref-1 (44, 88, 91, 109). Furthermore, it is well documented that both UVA and UVB cause skin inflammation with release of inflammation mediators like cytokines (IL-1 and IL-6), which further produce more ROS and increase oxidative stress (29, 107). Activation of NF-κB and AP-1 play a critical role in regulating the transcription of numerous genes involved in the immune and inflammatory response (45, 101), in which DNA-binding activity is markedly enhanced by APE/Ref-1 (27). In addition to indirect regulation of inflammation, many studies also revealed distinct induction of APE/Ref-1 in response to inflammatory stresses such as infection and asthma (3, 80, 83). For example, Helicobacter pylori–induced IL-8 gene transcription is also dependent on APE/Ref-1. As two sides of the same coin, APE/Ref-1 has beneficial effects and protects cells from ROS toxicity, but conversely, consistent induction or activation of APE/Ref-1 in response to prolonged oxidative stress switches the cellular signaling to a proliferation/antiapoptosis phenotype.
In melanoma cells, certain metals in combination with UVR, generates low-grade redox cycling with ongoing ROS generation, resulting in progressive and diffuse genetic and other cellular damage. The imbalance between pro- and antioxidants results in the activation of redox-sensitive signal transcriptions, such as AP-1 and NF-κB (15). AP-1 specifically regulates transcription of tetradecanoylphorbol 13-acetate (TPA)-responsive element (TRE)-containing genes by acting on their promoters (25), such as cyclin D1 (104) and p21 (20), which are important regulators of the cell cycle. AP-1 target genes are differentially regulated by distinct AP-1 dimers. In melanoma cells, expression of both c-Jun and JunD is evident (60, 121). Our studies suggest that JunD may be more mitogenic (121), which might be due to its cooperation with NF-κB signaling (92, 111). An activated NF-κB pathway is clearly important in melanomagenesis also (55, 67, 90), not only because this transcription factor regulates many genes (such as PTEN and FLIP) that are critically involved in apoptosis (24, 64), but also because NF-κB can be activated by antiapoptotic Bcl-2 overexpression, which suggests the existence of a forward-feedback loop (97). Our previous extensive studies have shown that in human melanoma cells, abnormal redox status is present compared with normal tissue, and activated ROS-mediated signaling was prominent (i.e., AP-1 and NF-κB) (67, 68, 73, 76, 121).
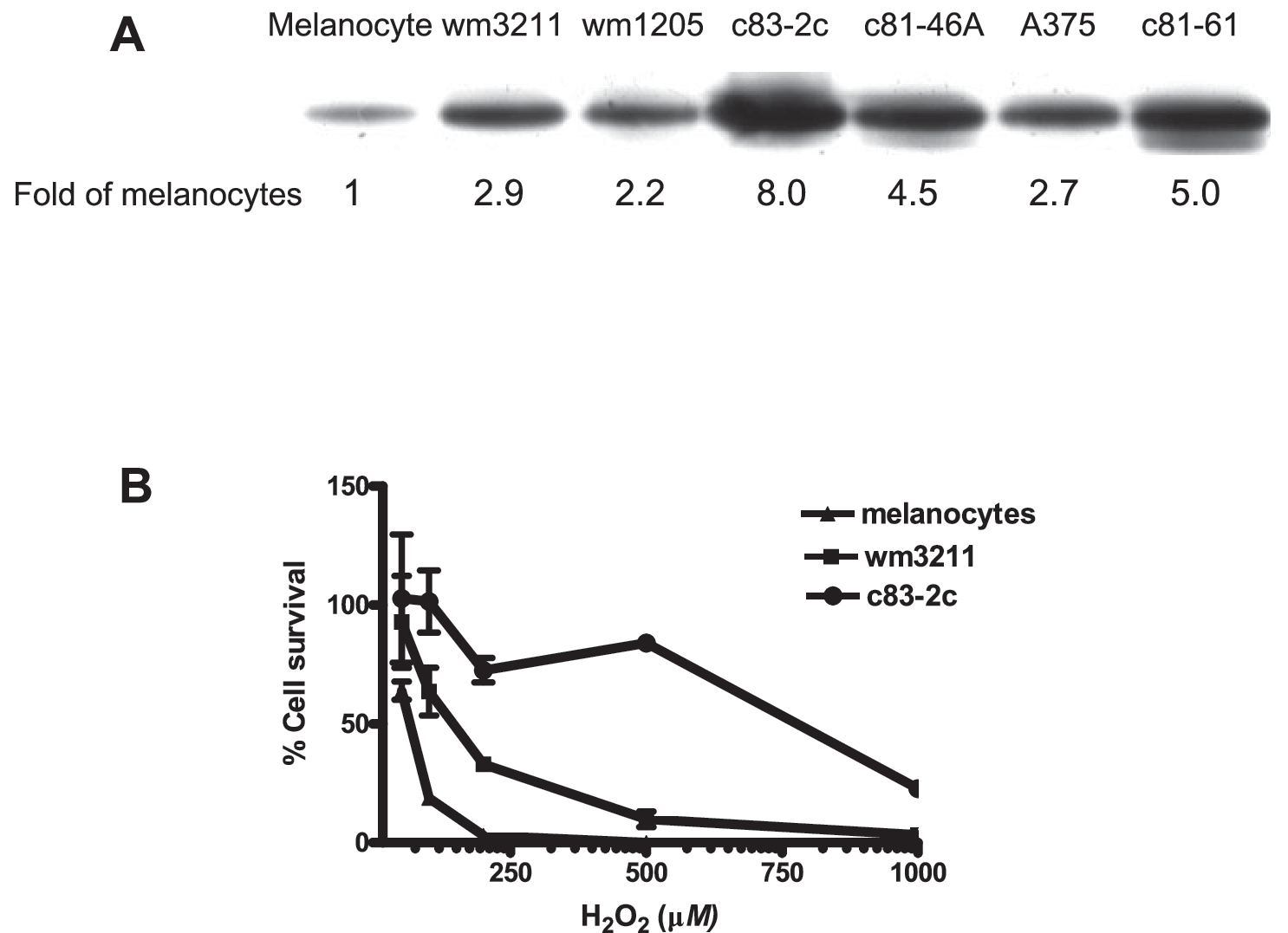
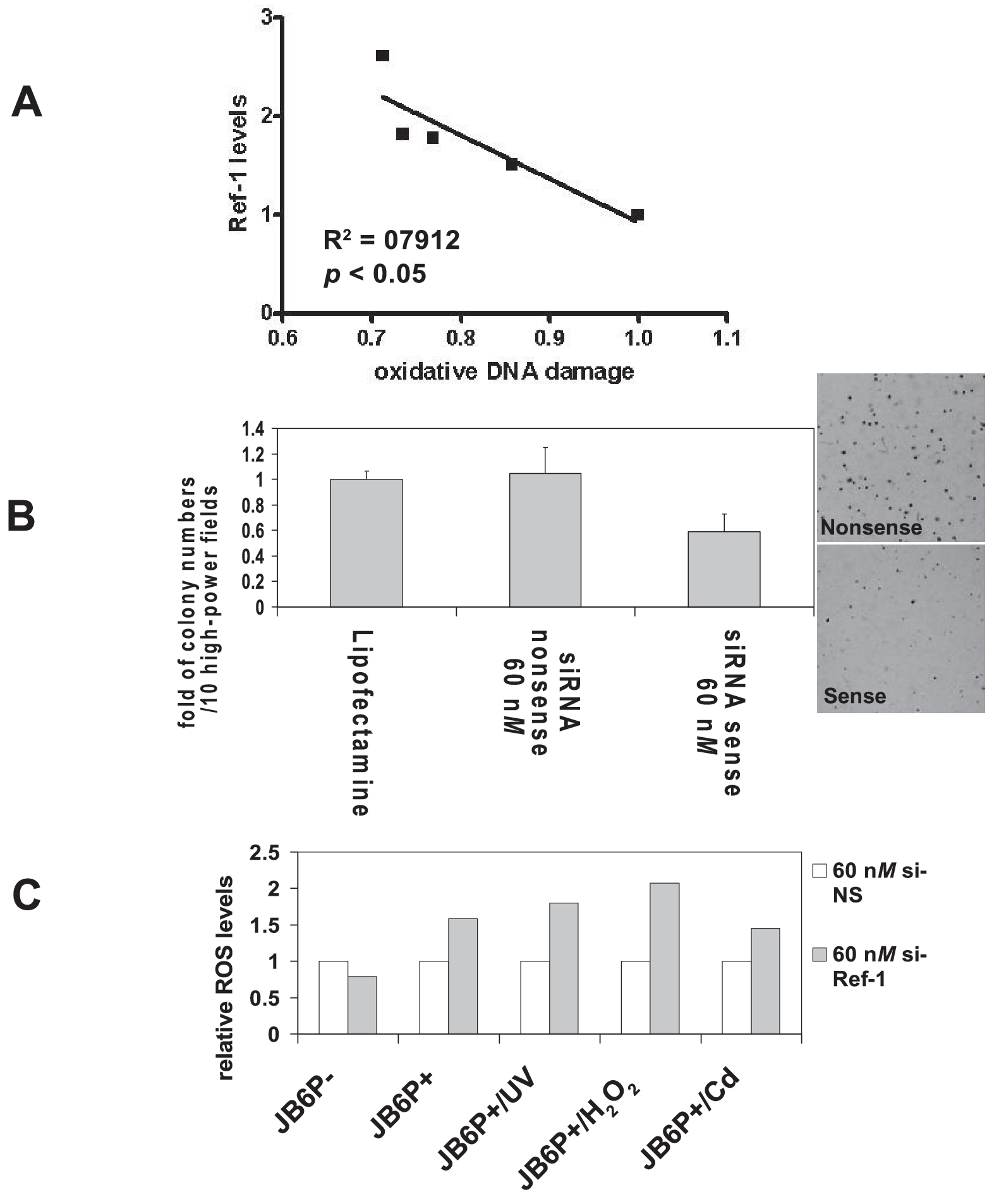
Our previous studies also demonstrated a remarkable increase of APE/Ref-1 expression levels in all tested melanoma biopsies and cell lines (Fig. 2), which was predominantly localized in the nucleus and contributed to the binding and activation of AP-1 and NF-κB (120). We characterized the APE/Ref-1 response in a series of JB6 cells (122) and found that elevated APE/Ref-1 was associated with decreased intracellular ROS levels as well as reduced oxidative DNA-damage lesions (Fig. 3). Also, depletion of APE/Ref-1 resulted in apoptosis with more ROS production and markedly reduced AP-1 transcription activities (122). Our studies also suggested that, as an adaptive response, induced APE/Ref-1 counteracts ROS stress not only by efficiently repairing oxidative DNA damage, but also through regulating redox-sensitive signaling (such as AP-1 and NF-κB).
Effects of Metals Involved in Melanomagenesis on APE/Ref-1 and APE/Ref-1–Mediated Signaling
Metals also likely play a crucial role in melanomagenesis (Fig. 4). Natural melanins are associated with a number of metal ions and have the capacity to accumulate metals. As characterized in our previous publications, we postulate that metals play an important role in converting melanin from a normal reducing status to a prooxidant state. Many metals bind to melanin with high affinity, especially certain heavy metals with redox potential, such as Cu(II) and Fe(III) (43). Additional studies also showed that Cu(II)- and Fe(III)-loaded melanin generally caused more DNA damage than Mg(II)-, Ca(II)-, or Zn(II)-loaded melanin (42). The literature indicates that supplemental copper and iron facilitate tumor growth, especially melanoma (52, 86).

Very interestingly, three large epidemiologic studies of the risk of cancer after hip replacements have been reported (81, 83, 113). Remarkably, patients with prior metal-on-metal hip replacements were identified at increased risk for cutaneous melanoma, prostate, and possibly kidney malignancies in all three studies. However, no increased risk was noted for patients with polyurethane-on-metal hip replacements. Other notable features of these studies were that the risk for melanoma increased with follow-up time in the two cohort studies. Other investigations have shown that serum and urine Co and Cr levels are markedly elevated (threefold to 10-fold) in patients with metal-on-metal hip replacements compared to controls (105). Consistent with these findings, substantial evidence shows that Co and Cr ions accumulate in melanin. Bogacz measured the affinity of various heavy metal ions for melanin in vitro, and ranked Fe3− > Cr3+ > Co2+ > Zn2+ > Mn2+ (13). Interestingly, Co and Cr affect the chemical properties of melanin and cell pigmentation (6, 84).
Depending on the system and experimental condition used, arguments both support and counter the importance of ROS in Cr genotoxicity (16, 112). In welding workers, who are exposed to high amount of vaporized Cr as well as other heavy metals, skin irritations and malignant melanoma were reported (72). Cobalt is a nonessential metal for which sufficient evidence for carcinogenicity exists in animals (IARC), with a high level of DNA-damaging capacity (38). Cobalt also induces ROS production through the Fenton reaction to generate hydroxyl radicals (·OH) in a perinuclear iron-dependent manner. The study of Leonard et al. (1998) showed that cobalt induced a significant increase in the generation of a whole spectrum of ROS (58). Although Co(II) does not react with hydrogen peroxide by the classic Fenton reaction at physiologic pH values, Co (I) mediates a Fenton-like reaction producing ·OH, which was increased remarkably in the presence of Fe2+.
Moreover, Co also mediates ROS generation in an indirect way. CoCl2 treatment is a classic model widely used to mimic the effects of hypoxia (89). Hypoxia-inducible factor-1 is a major oxygen- and metal-responsive transcription factor, which is also a key factor in mediating keratinocytes response to UVB exposure (95). Cobalt efficiently induces the activation of HIF-1 by the production of superoxide (65). Of great interest are studies that show APE/Ref-1 is a critical component of the hypoxia-inducible transcriptional complex that interacts with HIF-1 and p300 (33, 128). In addition, APE/Ref-1 was essential for the full activity of the C-terminal region of HIF-1, which contains its transactivation domain (56). However, no significant changes of APE/Ref-1 expression levels were evident with exposure to CoCl2 (50 μM) in melanoma cells (unpublished data). Whether such an effect occurs in normal melanocytes will be important to determine.
Another very interesting metal that is likely involved in melanomagenesis is iron. It is well documented that on exposure to UVR, an increase in ferric/ferrous iron occurs in the skin, and topical application of iron chelators reduces UVR-induced ROS production and skin damage, indicating a role of iron in photodamage (5, 47, 78, 94, 119). As shown in Fig. 4B, we postulate that photo-induced release of a pool of iron cations in response to blistering sunburn and the binding of Fe2− and/or Fe3+ and binding to at-risk melanin (i.e., pheomelanin and certain types of eumelanin, especially when partially oxidized), initiate low-level oxidative stress. Iron not only directly evokes ROS production by the Fenton reaction, but also potentiates Co-mediated generation of ·OH at physiologic pH values (58). Korytowski et al. (1987) studied the reactive species produced on irradiation of melanin with UVR and visible light (53). Their study showed that the production of ROS increased when melanin was complexed with iron in the presence of EDTA. Notably, melanin in the presence of low-dosage iron is associated with a low level of hydroxyl radical production; however, when melanin is saturated with Fe(III), ROS production occurs in a dose-dependent manner (87, 124). In addition, a recent study also demonstrated the pro-inflammatory activity of iron in the lung injury, at least in part, because of its induction of redox-sensitive factors (for example, TRX, APE/Ref-1, and NF-κB) (32).
The endonuclease activity of APE/Ref-1 has been shown to be affected by many factors, including different metals. The divalent metal ion soaked with the protein crystals (Sm2+ for APE-1) was found specifically to associate with the glutamate residue. Biochemical studies have shown that an APE-1 mutant with E96A change displays a significantly reduced DNA-repair activity in Mg2+ when compared with the wild-type protein (7), further supporting the notion that this amino acid contributes to metal coordination. Of considerable interest is that in in vitro studies with fibroblasts, Co does not enhance UVR-induced cyclobutane pyrimidine dimers (CPD) and 8-oxo-dG lesions formation but rather inhibits their repair (34, 48). However, dimer removal was not affected by Pb(II), Cr(VI), Fe(III), or Sn(III) (106). Controversially, another study demonstrated the inhibitory effects of Pb(II) and Fe(II) on APE/Ref-1 activity but not Co(II) (66).
Interestingly, CoCl2 (500 μM) is essential for AP endonuclease assay. Levin et al. (1991) showed that the activity of Escherichia coli endonuclease IV after exposure to EDTA, a strong nonspecific metal chelator, was restored by incubation with CoCl2 (500 μM) and to a lesser extent by MnCl2 (59). In another study, NiCl2 and CoCl2 at 1 mM concentrations stimulated both NF-κB and AP-1 activities (114), which might be mediated by APE/Ref-1 activation. These studies suggested that the effects of Co on APE/Ref-1 are concentration dependent.
Novel Strategies Targeting APE/Ref-1 to Prevent/Treat Human Melanoma
A variety of observational and experimental studies generated interest in the role of APE/Ref-1–mediated signaling in cancer, especially human melanoma. First, APE/Ref-1 is very sensitive to redox-status alterations. ROS regulates its activity and expression on both transcriptional and posttranscriptional levels. Coupled with the observation that melanoma cells exhibit abnormal redox status, induction of APE/Ref-1 as an adaptive response to prolonged oxidative stress likely plays an important role in human melanoma-genesis. Our previous studies consistently demonstrated abnormally elevated nuclear APE/Ref-1 in human melanoma cells compared with normal melanocytes, also associated with drug resistance and proliferation. Recently, by using a series of JB6 cells, we provided evidence that APE/Ref-1, in combination with ROS, plays a key role in malignant cellular transformation (Fig. 3) (122). In addition, our recent findings strongly suggested that APE/Ref-1 is involved in the regulation of metastatic potential in melanoma cells (unpublished data). Second, as discussed earlier, metals involved in melanomagenesis, especially cobalt and iron, regulate APE/Ref-1 expression and activity directly and indirectly. Third, the NF-κB pathway, regulated by APE/Ref-1, is prominent in mediating cytokine activation of leukocytes in inflammation. The involvement of APE/Ref-1 in the process of inflammation, which also occurs after sunburn, indicates interference with melanomagenesis by targeting APE/Ref-1 as a potential preventive strategy. We propose that excess endogenous ROS in human melanoma cells may also compromise the efficacy of alkylating agents–based or radiation-based therapy by inducing APE/Ref-1 in human melanoma. Accumulating studies on the role of APE/Ref-1 in promotion, progression, and drug resistance in other types of tumors have consistently confirmed its potential as an attractive target for the development of new cancer preventive and therapeutic strategies (36, 108).
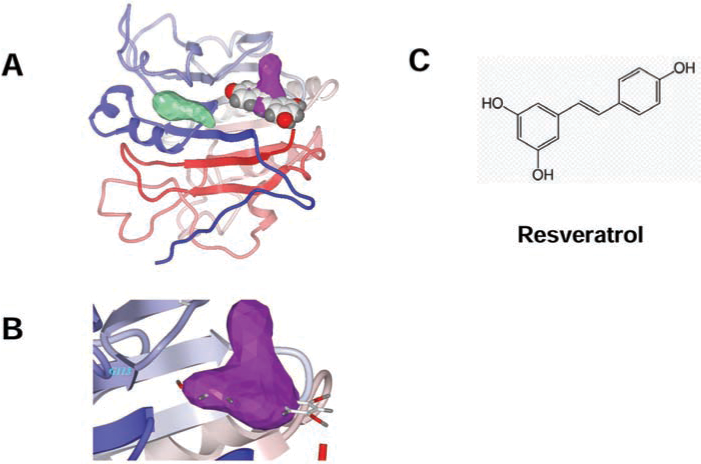
In recent years, the rapid increase in the number of high-resolution three-dimensional protein structures (1, 41), and the improvements of docking-and-scoring technology, make virtual screening (VS) an attractive and less-expensive alternative or complementary approach to the traditional methods of lead discovery and optimization (4, 51). Moreover, VS enables diverse compounds that would seldom be tested in a traditional laboratory high-throughput screen to be identified. The number of success stories from using VS in drug discovery keeps growing (69, 80). Recently we applied this docking strategy to the APE/Ref-1 receptor by screening chemical libraries (Fig. 5). Interestingly, resveratrol was found to dock into one of the two drug-treatable pockets located in the redox domain and, in preliminary screens, exhibited promising antimelanoma activities (120). The inhibitory effects of resveratrol on APE/Ref-1 occurred mostly through its redox-regulating functions and might be the major contribution to its pharmacologic activities, which are associated with significantly reduced AP-1 and NF-κB activities in many different human cancers (12, 54). Surprisingly, all other resveratrol analogues that we tested exhibited a lower docking score and lesser toxicity to human melanoma cells. In addition to resveratrol, some active lead compounds specifically targeting these two unique drug-treatable pockets were discovered by our screening. Further confirmation of these findings and chemical modifications are under way, and our early findings are discussed later. It is notable that APE/Ref-1 contains three distinct functional domains: nuclear-localization signal, redox-regulation, and DNA-repair domain. Limited experiments have been reported testing the distinct role of different domains in APE/Ref-1–mediated melanoma malignancies. These data would be critical for developing specific small-molecular inhibitors that interfere with distinct functions.
As APE/Ref-1 acts hierarchically to regulate many transcription factors (i.e., AP-1 and NF-κB), direct interference with APE/Ref-1 would be expected to result in a more-comprehensive effect than inhibition of just one downstream pathway. A small-molecule PNRI-299 (Fig. 6C) was identified as selective APE/Ref-1 inhibitor and has shown selective inhibition on AP-1 transcription (79). With a generous gift from Dr. Kahn, we tested the effects of PNRI-299 on human melanoma cells; however, at up to 100 μM concentration, no toxicity was evident in our test cell lines. One reason might be its specific inhibition of AP-1, which is markedly reduced in metastatic melanoma cells compared with normal melanocytes (121). However, exposure to intense bursts of sunlight, especially in childhood, is strongly associated with an increased risk for melanoma, whereas thick tumors (4 mm or greater), which have more metastatic potential, have significantly increased in men aged 60 years or older (23, 100). The long latent phase and slow progression of melanomagenesis provide a good opportunity for early intervention or chemoprevention. Specific APE/Ref-1 inhibitors (i.e., resveratrol analogues and PNRI-299), even without any direct melanoma cell–killing potential, might exert antiinflammation activities and be applied as preventive or photoprotective agents and protect skin from UV radiation–induced inflammation, which is mediated by AP-1 (14, 19). More recently, Luo et al. (61) reported that a small-molecule, 3-[5-(2,3-dimethoxy-6-methyl-1,4-ben-zoquinoyl)]-2-nonyl-2-propionic acid (E3330), exhibited remarkable inhibition of the redox function of APE/Ref-1 in vitro, and the IC50 is 6.5 μM, whereas no effects on repair activity of APE/Ref-1 were evident.
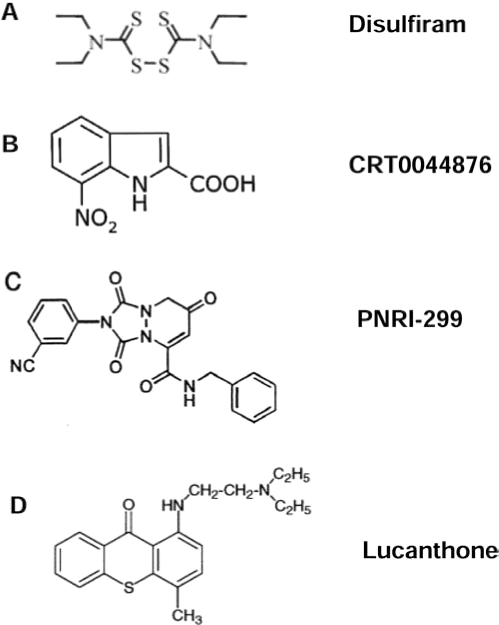
Lucanthone (Nilodin, Miracil D) was found to be an inhibitor of postradiation repair and is used as an adjuvant in radiation therapy (Fig. 1). A recent study showed that lucanthone increased the frequency of abasic sites in HeLa cell DNA, reflecting inhibition of APE repair activity, and enhanced the cell-killing effect of alkylating agents (62, 71). Notably, the APE/Ref-1 redox function or exonuclease activity on mismatched nucleotides was not affected by this compound. Additionally, by using fluorescence-based, high-throughput screening, a European group has isolated a small-molecule inhibitor, CRT0044876 (Fig. 6B), that binds to the active site of APE/Ref-1 and effectively inhibits its AP endonuclease, 3'-phosphodiesterase and 3'-phosphatase activities at low micromolar concentrations (63). Studies of in vivo efficacy and further chemical modification will be of great interest.
In addition to these direct APE/Ref-1 inhibitors, possible indirect ways may be found to inhibit APE/Ref-1–mediated signaling. Thioredoxin (TRX), a small cysteine-rich redox-active protein, directly associates with APE/Ref-1 in the nucleus and is essential for APE/Ref-1–mediated potentiation of AP-1 activity (40, 117). Combined with other observations that oxidized APE/Ref-1 lacks endonuclease activity (49), we propose that agents that block TRX/Ref-1 dimerization would decrease APE/Ref-1 nuclear translocation and induce APE/Ref-1 oxidation, resulting in subsequent APE/Ref-1 dysfunction.
Chelation therapy has been of great interest to medicine for several hundred years, although beneficial effects have been elusive in most cases. Adding to the potential mechanistic role of transition metals in melanomagenesis, metal particles are known to induce a vigorous macrophage–cytokine response associated with local osteolysis, which could serve as a promotional event for either melanoma or prostate cancer (105). Theoretically, based on our hypothesis, specific metal chelators could deplete metals and prevent the extra loading to melanin with less ROS production, which alternatively eliminates the stimulation of APE/Ref-1. In previous studies, we demonstrated that the S-based chelator pyrollidine dithiocarbamate (pDTC) strongly induced apoptosis in melanoma but was not toxic to melanocytes up to 10 μg/ml (28). In addition, the metal chelators o-phenanthroline (OP) and deferoxamine (DEF) were selectively toxic to human melanoma cells, whereas normal cultured melanocytes were resistant to OP- and DEF-induced changes (unpublished data). Notably, the alcohol-adversion drug disulfiram (DSF) (Fig. 6A) is a Cu-chelator and induces significant apoptosis in human melanoma at very low concentration (IC50 ∼20–50 ng/ml or ∼50–125 nM) through a distinct mechanism. It is Cu dependent, and addition of CuCl2 significantly enhances the DSF-induced cell death with a marked increase in intracellular Cu level and rapid ROS production. The cell-killing effect of DSF might be due to the extensive oxidation of protein initiated by Cu (17, 18). Recently, caged-iron chelators that are activated by UV have gained interests as an approach to protect against UV damage with fewer side effects and higher local protection (123).
Conclusions
Over the past decade, we have conducted a series of histologic, biochemical, chemical, and molecular experiments with melanin and human melanoma to explore the molecular mechanisms involved in melanomagenesis and melanoma progression. As summarized in Fig. 7, we proposed that prolonged oxidative stress initiated by UVR and melanin oxidation, a process enhanced by certain metals (cobalt, copper) found in the environment, is an early or primary event. The consequences of this phenomenon include many molecular changes, but as a key regulator, APE/Ref-1 is markedly induced and efficiently protects melanocytes from oxidative damage by inducing the antiapoptotic machinery and stimulating cell survival. Combined with other alterations, such as the depletion of cellular antioxidants and widespread oxidation of macro-molecules, APE/Ref-1 exhibits a critical role in melanoma-genesis and melanoma progression. A number of lead compounds show promising inhibitory effects on APE/Ref-1 and APE/Ref-1–mediated signaling, potentially with a wide range of indications from asthma to cancer therapy. The targeting of APE/Ref-1 may be a useful preventive and therapeutic strategy for the management of human melanoma and perhaps other cancers.
Footnotes
Acknowledgments
This study was supported in part by Chao Family Comprehensive Cancer Center, National Cancer Institute grant P30-CA62203, the Sun Fellowship Award, and the Oxnard and Waltmar Foundations.
Abbreviations
8-oxo-dG, 8-oxo-dihydro-2'-deoxyguanine; AP-1, activating protein-1; apurinic/apyrimidinic endonuclease-1/redox factor-1 (APE/Ref-1); DSF, disulfiram; EDTA, ethylenediaminetetraacetic acid; FLIP, FLICE-like inhibitory protein; HIF-1, hypoxia-inducible factor; NF-κB, nuclear factor kappa B; OP, o-phenanthroline; pDTC, pyrollidine dithiocarbamate; PTEN, phosphatase and tensin homologue deleted on chromosome 10; ROS, reactive oxygen species; SOD, superoxide dismutase; TRX, thioredoxin; UVR, ultraviolet radiation; VS, virtual screening.