
Abstract
Oxidative stress, resulting from mitochondrial dysfunction, excitotoxicity, or neuroinflammation, is implicated in numerous neurodegenerative conditions. Damage due to superoxide, hydroxyl radical, and peroxynitrite has been observed in diseases such as Alzheimer's disease, Parkinson's disease, and amyotrophic lateral sclerosis, as well as in acute conditions that lead to neuronal death, such as stroke and epilepsy. Antioxidant therapies to remove these toxic compounds have been of great interest in treating these disorders. Catalytic antioxidants mimic the activities of superoxide dismutase or catalase or both, detoxifying superoxide and hydrogen peroxide, and in some cases, peroxynitrite and other toxic species as well. Several compounds have demonstrated efficacy in in vitro and in animal models of neurodegeneration, leading to optimism that catalytic antioxidants may prove to be useful therapies in human disease. Antioxid. Redox Signal. 11, 555–569.
Reactive Oxygen Species, Reactive Nitrogen Species, and Oxidative Stress
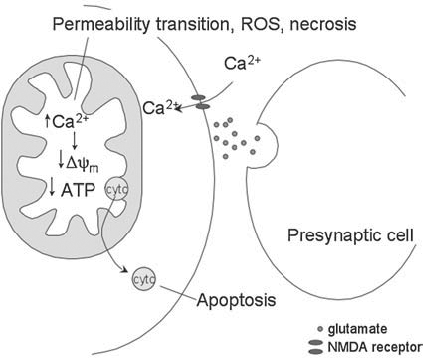
Reactive oxygen species (ROS) are generated in various cellular compartments as a consequence of normal metabolism, and under homeostatic conditions are kept in check by endogenous antioxidant defenses that include detoxifying enzymes such as superoxide dismutase (SOD), catalase, and glutathione peroxidase. Oxidative stress describes the pathologic condition in which the balance of oxidant generation and detoxification is tipped toward a prooxidant state, antioxidant defenses are overwhelmed, reactive species accumulate, and damage to nucleic acids, proteins, and membrane lipids ensues. Three phenomena leading to oxidative stress in the brain, and implicated in neurodegenerative disease, include the inhibition of mitochondrial metabolism (Fig. 1), neuronal excitotoxicity (Fig. 2), and neuroinflammation (Fig. 3).


In the mitochondrion, superoxide is formed as a result of electron “leak,” which allows the single-electron reduction of molecular oxygen. The level of superoxide formed under physiologic conditions, when mitochondria are respiring primarily on NAD-linked substrates, has been estimated to account for 0.1% to 0.4% of electron flow (23, 65). This superoxide is normally removed by the mitochondrial superoxide dismutase, MnSOD, which dismutates superoxide to hydrogen peroxide and oxygen. The significance of detoxifying endogenous mitochondrial superoxide is made clear by the severe pathologies present in the MnSOD-null mouse (86). Lacking MnSOD is either embryonic or neonatal lethal, depending on the genetic background (69). On the CD1 background, mutants survive ∼8 days, and develop fatty liver, metabolic acidosis, and cardiomyopathy (44, 86, 99). If treated with the catalytic antioxidant MnTBAP, the life span of the mice can be extended, and the liver and heart pathology can be prevented, but a neurodegenerative phenotype is uncovered, indicating the importance of detoxifying endogenous mitochondrial superoxide in the brain (57, 98). Other catalytic oxidants that are better able to cross the blood–brain barrier are able to rescue the spongiform encephalopathy (57, 100).
It is possible to increase the rate of mitochondrial superoxide production by increasing the oxidation state of the electron-transport chain (ETC), either by severely inhibiting ETC complexes or by otherwise increasing the mitochondrial membrane potential. Superoxide production increases dramatically when electron flow is inhibited at either complex I or complex III. This is of particular interest with regard to Parkinson disease (PD), as reduced activity of complex I has been observed in patients with PD (130, 131).
In addition to being a by-product of mitochondrial metabolism, superoxide is produced at the plasma membrane by NADPH oxidase, an enzyme found in many cell types and especially important to the function of immune cells. NADPH oxidase assembly is induced by exposure to growth factors and cytokines, and the resulting superoxide is dismutated to hydrogen peroxide by cytosolic or extracellular SOD or both. Mice null for the cytosolic and extracellular forms of SOD do not display the severe phenotypes observed in MnSOD-null mice, but do demonstrate reduced life span, increased oxidative damage to DNA, increased cancer rates, and increased sensitivity to some stresses (18, 19, 42, 69, 120).
Superoxide readily inactivates aconitase, releasing iron from the enzyme's labile Fe-S cluster (54, 85). However, superoxide has a short half-life and does not readily react with DNA or polyunsaturated lipids (51). Despite this, MnSOD-null mice have a severe neonatal lethal phenotype, which indicates that mitochondrially generated superoxide is highly toxic. The conversion of mitochondrially generated superoxide into more-reactive species, such as hydroxyl radical or peroxynitrite, probably accounts for much of its toxicity.
Superoxide spontaneously dismutes to hydrogen peroxide, a reaction catalyzed by SOD. Hydrogen peroxide is more stable than superoxide, with a longer half-life and the ability to diffuse through membranes, allowing it to function as a signaling molecule (for review, see 149). In the presence of free iron, hydrogen peroxide can react via the Fenton reaction to form hydroxyl radical, a very potent oxidant. Hydroxyl radical readily reacts with DNA, membrane lipids, and protein.
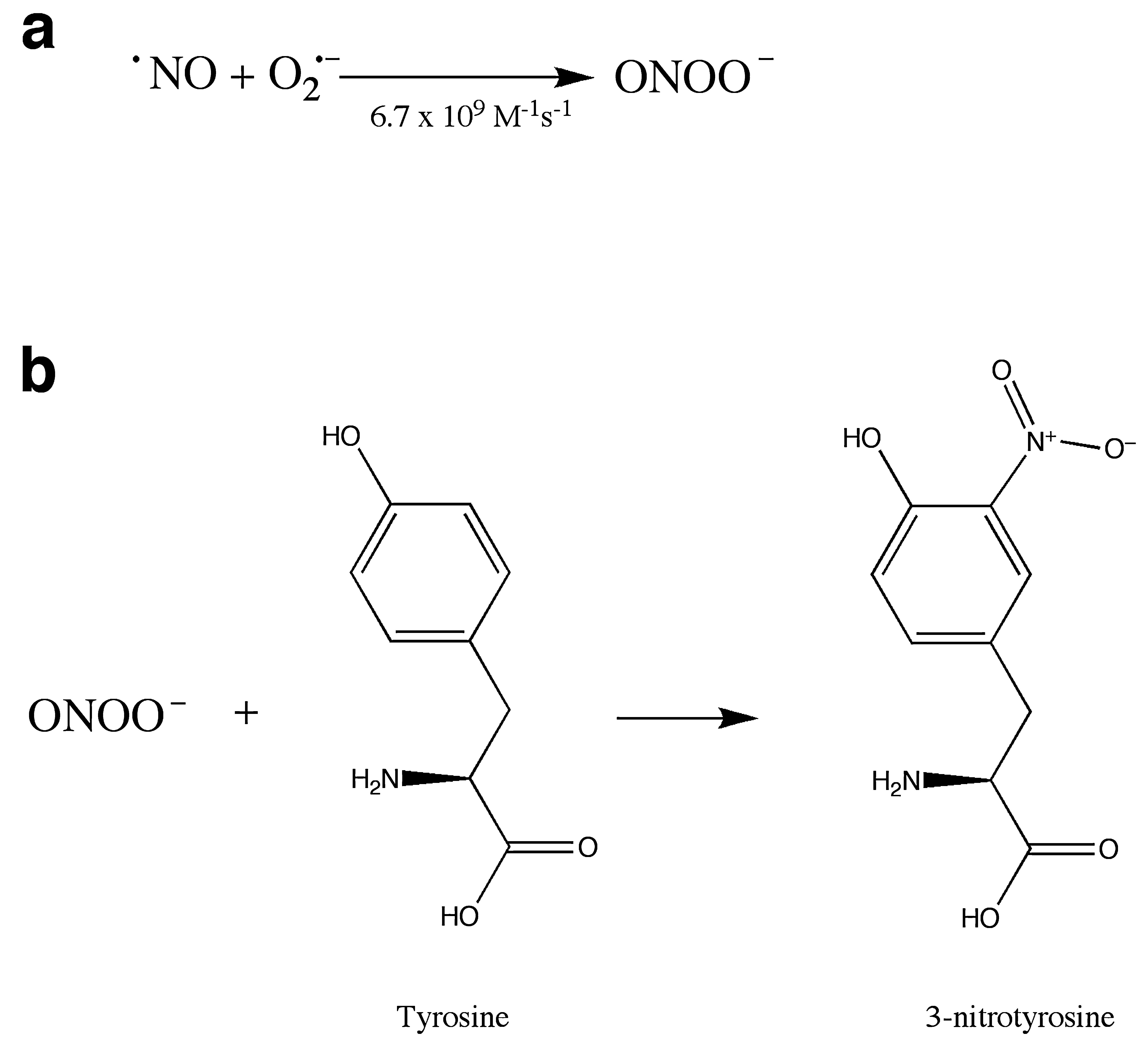
Reactive nitrogen species (RNS) also play a role in oxidative stress. Nitric oxide (NO) acts as a signal-transduction molecule in vasodilation (71, 107) and neuronal signaling (8), and nitrosylation is proposed to be a redox-sensitive protein modification involved in signal transduction (reviewed in 119, 141). NO also reacts with superoxide in a reaction that is diffusion limited, forming peroxynitrite (ONOO—) (Fig. 4a), a powerful oxidant. This reaction can serve as a mechanism to control levels of superoxide or NO or both, affecting cell signaling by NO. Additionally, peroxynitrite and its breakdown products can react with proteins, resulting in nitration of tyrosine residues (Fig. 4b) (73). Both superoxide and nitric oxide are produced in the course of the inflammatory response, leading to the formation of nitrotyrosine, which can be used as a molecular footprint of nitrosative stress in inflammation (80). Nitrotyrosine has been considered a hallmark of oxidative stress and is increased in many disease states, including Parkinson's disease (59) and Alzheimer's disease (67).
Catalytic Antioxidants
Currently, three major classes of metal-containing catalytic antioxidants are known: metalloporphyrins, macrocyclics, and salen compounds (Fig. 5). Each of these contains a manganese moiety that catalyzes the dismutation of superoxide in a mechanism analogous to that found in native SOD.
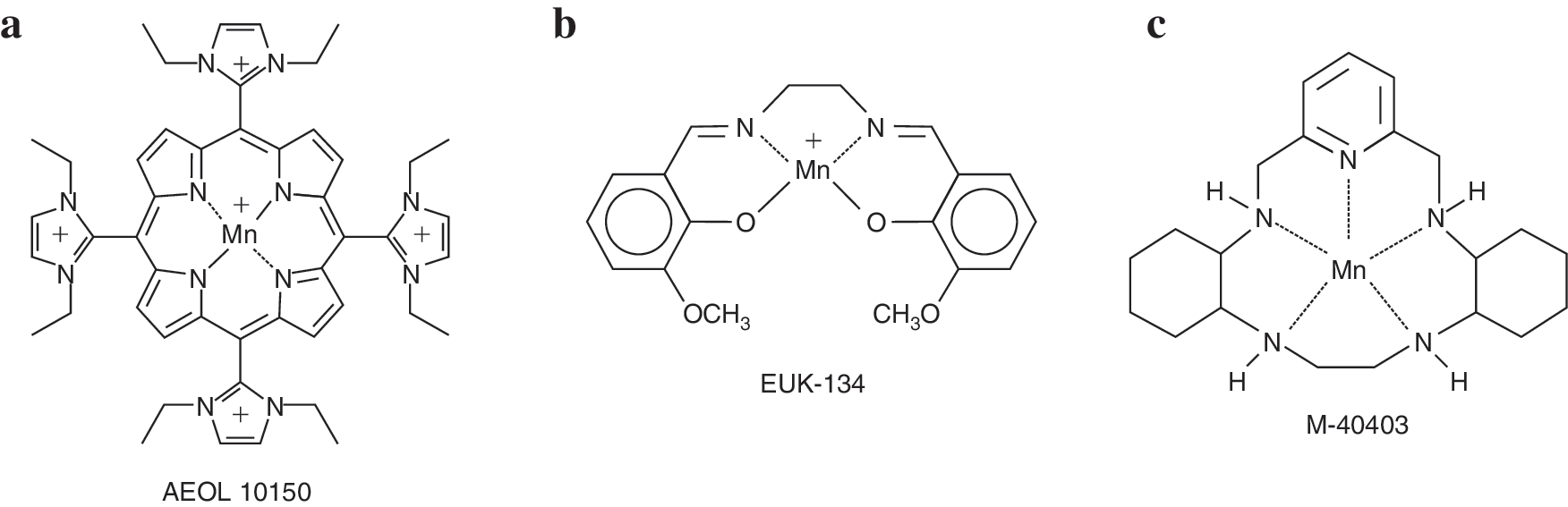
Metalloporphyrins [AEOL series, developed by Aeolus Pharmaceuticals, Inc. (Laguna Niguel, CA) a subsidiary of Incuara Pharmaceuticals] contain manganese coordinated by four axial nitrogen ligands. These compounds have been demonstrated to scavenge superoxide, hydrogen peroxide, lipid peroxyl radicals, and peroxynitrite (reviewed in 112). In the dismutation of superoxide, the manganese changes its valence state between Mn(III) and Mn(II) in a series of alternate reduction and oxidation steps, similar to the alternating reduction and oxidation that occurs in native SODs. In the dismutation of hydrogen peroxide, the conjugated ring system may undergo reversible one-electron oxidation, analogous to the heme prosthetic groups of catalases (38). In general, SOD and catalase activities have been correlated in the metalloporphyrins, but recently developed members of the family have relatively high catalase and lipid peroxidation inhibitory activity and low SOD activity (146), allowing investigation of new hypotheses regarding in vivo activity. The older generation of metalloporphyrins (i.e., pyridine and carboxylic acid derivatives) have only <1% of the catalase activity of native catalases, yet they are able to protect against hydrogen peroxide (33). The scavenging of lipid peroxyl radicals and of peroxynitrite may involve formation of an oxo-Mn(IV) complex, which is reduced by endogenous antioxidants such as ascorbate and glutathione (83).
Salen compounds (EUK series, developed by Eukarion, Inc., Bedford, MA, acquired by Tyrian Diagnostics, North Ryde, Sydney, Australia, in 2005) are similar to the metalloporphyrins in that they contain a manganese coordinated by four axial ligands, but the four axial ligands of salen compounds consist of two oxygen and two nitrogen atoms rather than four nitrogen atoms. EUK compounds have been demonstrated to scavenge superoxide and hydrogen peroxide, to react with ONOO− (133), and may react with lipid peroxides as well. Their mechanisms of action are thought to be similar to those of the metalloporphyrins.
Macrocyclics (M series developed by MetaPhore Pharmaceuticals, Inc., Fort Lee, NJ and St. Louis, MO) contain a manganese atom coordinated to five nitrogen ligands. The pentavalent coordination allows the manganese to participate only in single-electron transfers, and so makes these compounds specific for the scavenging of superoxide, relative to hydrogen peroxide or peroxynitrite, which require two-electron transfers. Other compounds, such as flavins and ubiquinones, can participate in one-electron transfers, so the macrocyclics may react with compounds besides superoxide in vivo.
The carboxyfullerenes are an additional class of compounds that can function as radical traps, but one member of which (C3; Fig. 6) has been described as having SOD activity with a rate-constant ∼100-fold slower than endogenous SOD (5). These compounds do not contain metal centers, but are thought to attract a superoxide molecule to an electron-deficient area of the C3 molecule, holding the superoxide there until a second superoxide molecule approaches and the dismutation reaction can occur with help from carboxyl groups on C3 or from water molecules (5). The compounds can also scavenge hydroxyl radical and other free radicals, probably because of their highly conjugated double-bond systems. C3 is concentrated in the mitochondria.

The antioxidant properties of each of these families of compounds have been studied in vitro, and, as is discussed later, several compounds have been demonstrated to have biologic activity in vivo. A question remains, however, regarding which antioxidant activity is most important to protection in a given system. Compound activities in vivo do not always correlate with the activities measured in vitro, making it difficult to predict activity in a disease model. A compound may participate in additional reactions or demonstrate novel activities in a biologic context. In vivo experiments comparing different classes of compounds in the same model have not been done, but would be informative in determining mechanisms.
Additional modification of compound activities is achieved by localization, both within the cell and within the organism. Compounds are being engineered to improve their ability to cross the blood–brain barrier, to increase their concentration in hydrophobic membranes, or to target the mitochondria. Regardless of its activities as measured in vitro, the concentration of a compound in the correct compartment or tissue may dramatically affect its ability to demonstrate efficacy in a disease model.
Catalytic Antioxidants and Neurodegeneration
Parkinson's disease
Parkinson's disease (PD) is a progressive disorder characterized by the selective death of dopaminergic neurons in the substantia nigra. Approximately 15% of cases are familial, with five causal genes identified to date (reviewed in 108). The functions of the proteins encoded by these genes implicate protein degradation, mitochondrial function, and other processes in the disease. How these inherited forms of the disease relate to sporadic PD is unclear. PD patients have reduced complex I activity in the substantia nigra relative to age-matched controls (75, 130). Focus on complex I is further supported by the discovery that MPTP, a compound that induces a Parkinson-like syndrome in humans and other organisms, including mice, is an inhibitor of complex I (27, 68, 91, 138).
Inhibition of complex I could result in reduced ATP production, leading to an energy-starved state that would interfere with the maintenance of membrane ion gradients and neuronal function. Additionally, inhibition of complex I might lead to an increase in ROS, which could damage the dopaminergic neurons and lead to cell death. Evidence of oxidative stress is seen in PD brain, including increased oxidative modification of DNA (4), proteins (3) and membrane lipids (35, 36). The hypothesis that inhibition of complex I may contribute to the development of PD has led to a search for environmental toxins that possess this function. Epidemiologic studies implicate exposure to paraquat, an herbicide, as a risk factor in acquiring PD (68, 91. For review, see 16).
Another hallmark of PD is increased inflammation. Microglia, the resident immune cells of the brain, are activated and proliferate in the substantia nigra in PD. The cells are likely responding to the death of dopamine neurons, and their proliferation and activation contribute to additional death, essentially resulting in an amplification of the pathology. Antiinflammatory drugs have been found to be protective in an MPTP mouse model of PD (39, 156). Additionally, population studies suggested that nonsteroidal antiinflammatory drug use may reduce the chance of developing PD (24, 151), although other studies found no protective effect (13, 46).
Catalytic antioxidants have been found to be neuroprotective in several animal models of PD. Two EUK compounds, EUK-134 and EUK-189, were demonstrated to protect against apoptotic cell death in dopaminergic cells treated with paraquat and to protect against neuronal death in primary neuronal cultures (113). In the same study, pretreatment with EUK-189 prevented the loss of nigral dopaminergic neurons in mice exposed to paraquat (113), and it was proposed that the EUK compounds exerted their effect by inhibiting the activation of JNK-mediated apoptosis. Pretreatment of primary dopaminergic cultures with EUK-134 prevented the accumulation of nitrotyrosine in tyrosine hydroxylase in response to either MPP+ or 6-hydroxydopamine (117). Carboxyfullerene (94) and MnTBAP (a metalloporphyrin) (26) were able to protect against 6-hydroxydopamine, but not against MPP + toxicity.
A major advancement in the field of catalytic antioxidants was the demonstration that AEOL 11207, a lipophilic metalloporphyrin, protected against MPTP toxicity in vivo when administered orally (89). This compound is a member of a new series of metalloporphyrins, the AEOL 112 series of glyoxylate metalloporphyrins (146), which were designed to have greater lipid solubility, oral bioavailability, and ability to cross the blood–brain barrier. In vitro assays were used to rank compounds from the AEOL 112 series with respect to ability to scavenge hydrogen peroxide generated by paraquat-treated mitochondria (21). The IC50 measured for AEOL 11207 in the in vitro assay, 104 nM, agreed well with the brain concentration reached in vivo (∼200 nM) at which antioxidant effects were observed (89). Treatment of MPTP-intoxicated mice with the compound AEOL 11207 reduced the death of dopaminergic neurons in the substantia nigra, the decrease in dopamine levels in the striatum, and markers of oxidative damage including 3-nitrotyrosine levels, 4-hydroxy-2-nonenol levels, and a decrease in the ratio of oxidized to reduced glutathione (89). The oral efficacy of AEOL 11207 in a well-established model of PD suggests the potential clinical utility of this class of compounds.
The future applicability of catalytic antioxidants to PD is promising, but substantial issues remain. First, in animal and cell-culture models, catalytic antioxidants have been administered before or concurrently with exposure to the toxin or both, which cannot mimic the situation in human beings. Individuals do not demonstrate parkinsonian symptoms until they have lost a large percentage of their dopaminergic neurons, at which time interventions to prevent cell death may be too late. Additionally, the precise identity of the species against which the compounds are acting is unknown, because the compounds have demonstrated activity against superoxide, peroxide, and peroxynitrite. The compounds may be blocking ROS resulting from an initial inflammatory response, in addition to removing ROS resulting from mitochondrial dysfunction. In the case of AEOL 11207, it likely primarily targets membrane peroxides because the compound is lipophilic and has high relative catalase activity and weak SOD activity (21).
Alzheimer's disease
Alzheimer's disease (AD) is a progressive dementing disease involving loss of cortical and hippocampal neurons and characterized by the accumulation of extracellular amyloid plaques and intracellular neurofibrillary tangles. The majority of cases are sporadic, but early-onset familial AD cases have led to the identification of three genes encoding amyloid precursor protein (APP) and presenilins 1 and 2. The amyloid plaques observed in AD brain are composed of a 4-kDa peptide, β-amyloid (Aβ), which is a cleavage product of APP. The presenilins are involved in APP processing, and the convergence of the three known genetic determinants of AD on this one peptide has led to the amyloid hypothesis of AD, which proposes that the accumulation of Aβ is responsible for AD (56; for review, see 145). The only genetic risk factor yet confirmed for late-onset or sporadic AD is the ApoE4 allele of apolipoprotein E, which may indicate a role for cholesterol metabolism or vascular function in the disease.
A great deal of evidence has been accumulated for oxidative stress in AD, with increased damage detected to lipids (129), protein (139), nuclear DNA (53), mitochondrial DNA (97), and RNA (104) in AD relative to those in age-matched controls. Whether oxidative damage is responsible for the disease or merely associated with it has been difficult to determine. Increased oxidative damage was found to precede pathology in one study (105), but the lesions associated with AD can also be a source of ROS (see later). In some studies, diets rich in antioxidants have been associated with a decreased AD risk, but in others, no effect has been seen (reviewed in 140). Treatment with dietary antioxidants has not been demonstrated to slow disease progression (55, 127), although it has been hypothesized that the intervention may have been introduced too late into disease onset (reviewed in 140).
Aβ may directly contribute to the oxidative stress observed in AD. The peptide accumulates in mitochondria and can inhibit the electron-transport chain, particularly complex IV, cytochrome oxidase (COX) (30). However, decreased COX activity reduced plaque load and ROS in another study (52), challenging the proposition that COX inhibition may be a source of ROS in AD. Aβ interacts with ABAD, resulting in ROS production and apoptosis (95). Additionally, Aβ itself may enzymatically generate hydrogen peroxide and hydroxyl radical in the presence of copper (9, 30, 70, 106). Alternatively, it has been hypothesized that APP or Aβ or both could function as antioxidants under nonpathologic conditions (7).
Substantial evidence exists for neuroinflammation in AD (reviewed in 11, 124). Microglia are found within and around amyloid plaques and can be activated in vitro by Aβ peptide (76). Epidemiology studies indicate that NSAID therapy reduces the risk of AD (15, 142; reviewed in 143, 150). However, this protection may be significant only for those individuals with at least one ApoE4 allele (144). Except for one early positive result (123), clinical trials of NSAIDs have not demonstrated protection against disease progression (1, 2, 121, 132), or against disease incidence (62).
Catalytic antioxidants have demonstrated some efficacy against Aβ-induced toxicity in vitro. EUK-8 inhibited cell death in response to Aβ in organotypic hippocampal cultures (17). Microglial proliferation induced by fibrillar Aβ was blocked by catalase, EUK-8, and EUK-134 (76), leading to the conclusion that hydrogen peroxide resulting from activity of the NADPH oxidase mediates microglial activation by Aβ. Additionally, ROS from the NADPH oxidase have been implicated in mediating the effects of Aβ on vascular dysfunction (110). The Aβ peptide was found to accumulate as supranuclear cataracts in AD (58) and a mouse model of AD (101). Cataract development in the mouse model was reduced by treatment with EUK-189 (101), which provides the first example of a catalytic antioxidant protecting against an Aβ-related pathology in vivo.
Stroke
In ischemic stroke, blood flow is interrupted to a portion of the brain, usually by a clot blocking an artery. The subsequent reperfusion of the affected brain area when blood flow is resumed is accompanied by a burst of oxidative damage. A region of acute cell death results, with a surrounding penumbra of damaged tissue that degenerates more slowly. Rescuing the cells in the penumbra from death is the goal of most stroke-treatment models.
Reactive oxygen and nitrogen species have been implicated in the death of cells in the penumbra. A decrease in ascorbate levels, indicative of increased free radical damage, was detected after cerebral ischemia (50), and glutathione levels are decreased during reperfusion after ischemia (103, 109). Overexpression of Cu/Zn SOD protects against focal ischemic injury in adult mice (81), transient focal ischemia in mice (158), global ischemia and reperfusion injury in rats (22), and transient focal ischemia in rats (32), implying that superoxide is a toxic species in stroke. However, protection provided by SOD1 overexpression was dependant on the presence of glutathione peroxidase in a mouse model of focal ischemia/reperfusion (29), meaning that it is essential to detoxify the resulting hydrogen peroxide as well. Nitric oxide levels also are increased in ischemia (155). These levels, when generated by nNOS, are protective (43). However, a further increase in NO levels produced by iNOS can be damaging if NOS is converted to peroxynitrite in the presence of superoxide (43).
More than 100 clinical trials have been carried out with compounds proposed to work as neuroprotectants in stroke (61). In 1999, a series of guidelines was published for preclinical and clinical testing of drugs for acute ischemic stroke (AIS) by the Stroke Therapy Academic Industry Roundtable (STAIR). The guidelines for preclinical testing include the demonstration of efficacy in permanent and transient ischemia, in rodent and primate models, from multiple laboratories and with long-term follow-up, before clinical trials are warranted. A spin-trap agent, NXY-059, was believed to be a promising neuroprotective agent after animal studies and generated excitement after a positive effect in initial clinical trials [SAINT I, (84)]. However, a second, larger trial failed to demonstrate any protection by NXY-059 in acute ischemic stroke (137), demonstrating the difficulty in extending animal research to human trials, and leading some to question the feasibility of neuroprotection as an approach in treating stroke (152). Some authors, however, have used this as an example of the costs of not adhering closely enough to the 1999 STAIR criteria (128), after a reexamination of the initial animal studies.
Several catalytic antioxidants have proven effective in stroke models. The macrocyclics M40403 and M40401 decreased infarct volume when administered 2 h before hypoxia/ischemia injury (135). Especially promising with regard to treatment of human disease are models in which catalytic antioxidants protect against damage when administered several hours after the stroke event, mirroring the situation in which the patients cannot receive treatment until they reach the hospital and are diagnosed as having had a stroke. The macrocyclics were not protective when administered after transient focal ischemia (136), which may indicate that detoxifying superoxide is not sufficient to provide protection in the absence of added hydrogen peroxide detoxification. A single injection of EUK-134 3 h after middle cerebral artery occlusion (permanent regional cerebral ischemia) prevented further increase in infarct volume (10). In this study, EUK-134 was found to be more effective than EUK-8, a related compound with lesser catalase activity, consistent with the genetic study indicating that hydrogen peroxide is especially important to ischemia-related injury (29). A panel of related EUK compounds was tested in the same model, as well as a cell-culture model (37). The ability to protect in the culture model was related to the catalase activity of the compound, but interestingly, the activities of the compounds in the two assays were not always correlated, pointing out the difficulty in predicting in vivo activities (37). AEOL 10113 reduced infarct size when administered up to 6 h after focal ischemia (96). AEOL 10150 also reduced infarct size in a transient ischemia model (134) and attenuated the increase in inflammatory gene expression in response to transient middle cerebral artery occlusion (14). These results have led to optimism that catalytic antioxidants might have therapeutic value in stroke.
Epilepsy
Epilepsy is not classified as a neurodegenerative disease per se, but seizure activity leads to cell loss and neurodegeneration, which contribute to disease progression. Epilepsy can be age-related, developing in response to age-related diseases such as stroke. Evidence exists of oxidative stress accumulating in epilepsy. Status epilepticus causes a depletion of glutathione (88). Prolonged seizure activity results in excitotoxicity, which leads to an influx of extracellular calcium, a dysregulation of intracellular calcium, and mitochondrial ROS production (25, 40, 111, 122). The cell death that results is dependant on superoxide production (111), and that superoxide is produced by the mitochondria (85). The cell death may activate an inflammatory response, resulting in more oxidative damage (125).
Catalytic antioxidants have reduced oxidative damage in epileptic animals, although they have not been demonstrated to reduce seizure duration or latency. Pretreatment with EUK-134 prevented neuronal damage and decreased markers of oxidative damage, including protein nitration, resulting from kainite-induced seizures, although the compound did not affect the latency or duration of seizures (125). The catalytic antioxidant MnTBAP also inhibited kainite-induced oxidative DNA damage, aconitase inactivation, and neuronal death (87). Whether these compounds would reduce disease severity or progression remains to be tested.
Amyotrophic lateral sclerosis
Amyotrophic lateral sclerosis (ALS), or Lou Gehrig disease, involves the progressive loss of motor neurons, resulting in muscle atrophy. The majority of cases are sporadic, but 15–20% of familial cases are associated with mutations in SOD1. This led to the hypothesis that ALS was caused by an increase in superoxide; however, many of the mutations associated with the disease do not change the activity of SOD1, and sod-1–null mice do not develop ALS. Therefore, a gain of toxic function is proposed for the SOD1 mutations, although that function has not yet been identified. Hypotheses include a copper-dependent ROS-generating function of mutant SOD1 (45, 63), a propensity of mutant SOD1 to aggregate (74, 77), increased apoptosis due to Bcl-2 sequestration by aggregated SOD1 (118), and an ability of mutant SOD1 to bind to rac, activating the NADPH oxidase on microglial cells and enhancing superoxide production (66).
The mutation acts non–cell-autonomously, in that astrocytes carrying the mutation can induce apoptosis in nonmutant neurons (102, 148), and nonmutant astrocytes can extend the survival of mutant neurons (28), although with high enough expression levels, expression of mutant SOD1 only in neurons is enough to cause motor neuron disease (74). Interestingly, overexpression of wild-type SOD1 results in the formation of aggregates in astrocytes (74), and oxidation of wild-type SOD1 can cause it to misfold and gain some of the toxic properties of mutant SOD1 (47), leading to the hypothesis that oxidative stress and oxidative damage of wild-type SOD1 could be behind sporadic ALS (79).
Despite the lack of support for the hypothesis that SOD1 mutations lead to an increase in superoxide because of a decrease in SOD activity, substantial evidence exists for increased oxidative stress in ALS. Mice carrying a mutant human form of SOD1 accumulate damaged DNA, protein, and lipid (64, 92, 154), and neural tissue from ALS patients also demonstrates oxidative damage (48, 49, 72). In a cell-culture model, expression of several mutant forms of SOD1 was found to cause an increase in mitochondrial superoxide and neuronal cell death, and cell survival was increased by a concomitant overexpression of SOD2 (160). Expression of mutant SOD1 resulted in mitochondrial dysfunction and increased oxidative and nitrative damage in astrocytes, which could be rescued by mitochondrially targeted antioxidants (20). Overexpression of SOD2 and phospholipid glutathione peroxidase (93) and treatment with a mitochondrially targeted antioxidant peptide (114) extended the life span in a mouse model of ALS. Copper chelators can also extend survival, consistent with a metal-catalyzed oxidative-stress component of ALS (116).
Deleting the gene for gp91, the catalytic component of the microglial NADPH oxidase, blocked neuronal cell death in a mouse ALS model (157), supporting a role for neuroinflammation. However, a clinical trial of minocycline, an antiinflammatory drug, did not slow disease progression (60), although it was protective in an animal model of the disease (82, 147). This points out a long-running issue of lack of correlation between in vitro and animal and human studies in ALS (reviewed in 12), analogous to that seen for neuroprotective drugs in stroke.
Catalytic antioxidants have shown promise in animal models of ALS. AEOL 10150 extended survival from symptom onset up to threefold, when the drug was administered to each mouse on first day of muscle weakness in the mouse G93A model of ALS (31). In this same study, AEOL 10150 was combined with dietary creatine and rofecoxib, with no additional increase in survival. In another study, AEOL 10150 was administered alone or in combination with the histone deacetylase inhibitor phenylbutyrate (PBA) at symptom onset in the G93A mouse model (115). All three paradigms were found to extend survival, but the combination therapy was the most effective, extending survival by 19%. EUK-8 and EUK-134 also extended survival when administered before disease onset (78), and C3 similarly delayed symptom onset and death (41). Human trials with a reactive oxygen and reactive nitrogen species scavenger R+ pramipexole demonstrate the drug is well tolerated and hint at positive effect on disease progression (153), providing hope that antioxidant strategies may translate into treating human disease.
Conclusions
Oxidative stress is a component of and plays a complex role in many neurodegenerative diseases. In some cases, oxidative stress is secondary to other pathology (e.g., oxidative damage occurs as the result of an immune response initiated by cell death). In other cases, oxidative damage due to mitochondrial dysfunction is a primary cause of cell death and neuronal loss. Catalytic antioxidants can intervene at each of these steps, preventing cell death in response to mitochondrial dysfunction or excitotoxicity and preventing the immune-response–driven vicious cycle of increased oxidative and nitrative damage, which can contribute to subsequent cell death and disease progression. Therefore, multiple ways exist in which catalytic antioxidants can exert the protective effects observed in the in vitro and in vivo models of neurodegeneration, and much promise is seen that these compounds may present useful therapies in human disease.
Footnotes
Abbreviations
Aβ, amyloid beta; AD, Alzheimer's disease; AIS, acute ischemic stroke; ALS, amyotrophic lateral sclerosis; ApoE, apolipoprotein E; APP, amyloid precursor protein; DNA, deoxyribonucleic acid; ETC, electron-transport chain; iNOS, inducible nitric oxide synthase; MnSOD, manganese superoxide dismutase; MnTBAP, manganese (III) tetrakis (4-benzoic acid)porphyrin; MPP+, 1-methyl-4-phenylpyridinium; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; NAD, nicotineamide adenine dinucleotide; NADPH, nicotineamide adenine dinucleotide phosphate; nNOS, neuronal nitric oxide synthase; NO, nitric oxide; NSAID, nonsteroidal antiinflammatory drug; ONOO−, peroxynitrite; PD, Parkinson's disease; RNA, ribonucleic acid; RNS, reactive nitrogen species; ROS, reactive oxygen species; SOD, superoxide dismutase; SOD1, intracellular Cu/Zn superoxide dismutase; SOD2, mitochondrial manganese superoxide dismutase; STAIR, Stroke Therapy Academic Industry Roundtable.
Acknowledgments
Dr. Patel has received grant funding from the National Institutes of Health (RONS45748, R01NS39587, R21NS53548, and Aeolus Pharmaceuticals, a company that develops metallopophyrin antioxidants for human disease.