
Abstract
Endothelial cells in vivo are constantly exposed to mechanical forces such as cyclic strain. In endothelial cells, Nox4-containing NAD(P)H oxidase complexes have been identified as major sources of superoxide anion (·O2 −) formation. In this study, we analyzed the effect of cyclic strain on endothelial ROS formation by electron paramagnetic resonance spectroscopy, cytochrome c assay, and dihydroethidium fluorescence, on NO formation by Griess reaction and on gene expression by RT-PCR and Western blot. Primary cultures of human umbilical vein endothelial cells were exposed to 2–18% cyclic strain for up to 24 h using the Flexercell system. Long-term application of 5–12% cyclic strain downregulated Nox4 expression and ROS formation in a time-dependent manner. Downregulation of Nox4 was further confirmed by promoter analysis using dual-luciferase assay. Cu/Zn SOD, MnSOD, and catalase expression was decreased after application of chronic 12% cyclic strain. In contrast, endothelial NO formation and eNOS were increased by cyclic strain. Strain-dependent Nox4 downregulation was abolished by eNOS inhibition with L-NAME. In conclusion, physiological levels of cyclic strain downregulate Nox4 expression and superoxide anion formation. This novel mechanism might contribute to a vasoprotective balance between NO and superoxide anions in response to physiological mechanical stimulation of endothelial cells. Antioxid. Redox Signal. 11, 2385–2397.
Introduction
Endothelial cells can differentiate between hemodynamic stimuli such as shear stress or strain (24). While a variety of experimental studies has analyzed the impact of shear stress on endothelial cells, little is known about the impact of cyclic mechanical strain on endothelial function. Cyclic strain is defined as a repeated mechanical deformation of the vessel wall through the transmission of heart frequency and blood pressure amplitude (23). Under physiological conditions, in vivo measurements in different species predicted 5–6% cyclic strain, which can be increased to >20% under hypertensive conditions (41, 49). The application of cyclic strain causes an adaptation of the vascular cells by increased formation of actin stress fibers and reorganization of actin filaments (6, 25, 61). Under pathophysiological conditions such as hypertension, biomechanical forces can alter endothelial and vascular smooth muscle cells remodeling (29). Cyclic mechanical strain can influence different cellular functions such as proliferation (32) and apoptosis (26) in endothelial cells.
The knowledge about the impact of cyclic mechanical strain on the formation of reactive oxygen species is limited. Cyclic strain increased the superoxide anion formation in endothelial and vascular smooth muscle cells (35). This could be blocked by a dominant-negative mutant of Rac (58). Recently, Sung et al. reported an opposite effect on reactive oxygen species (ROS) formation by cyclic strain in venous or arterial endothelial cells (52). However, the molecular source for this ROS formation in response to cyclic mechanical strain has not been characterized.
A major source of reactive oxygen species in the vessel wall are NAD(P)H oxidase complexes (17). Specific vascular NAD(P)H oxidase complexes seems to play a key role in the response to cyclic strain. Mechanical stretch of vascular smooth muscle cells resulted in a rapid ROS formation and p47phox membrane translocation that is followed by an increase in Nox-1 transcripts thereby enhances MMP-2 mRNA expression and pro-MMP-2 release (18). Recently it has been shown that physiological strain increases but pathological strain decreases endothelial cell survival and tubulogenesis, and these effects require a NAD(P)H oxidase subunit p22phox–dependent pathway (26).
In endothelial cells, a Nox4-containing-NAD(P)H oxidase complex has been identified as a major source of superoxide anion (·O2 −) and H2O2 formation (3, 33, 46). Nox4 siRNA inhibits VEGF-induced endothelial cell migration and proliferation, while overexpression of dominant negative Nox4 blocks PDGF receptor autophosphorylation (8). Recently, Natarajan et al. reported that Nox4-generated ROS regulate endothelial cell motility (39). In vitro studies based on either antisense or siRNA suppression suggested a role of Nox4 in smooth muscle cell and endothelial cell proliferation (37, 42, 51). In rat aortic vascular smooth muscle cells, Nox4 is responsible for basal H2O2 production, while ·O2 − production is dependent on Nox1 (9). Recently we observed by DNA microarray, real-time PCR, and Western blot, that the Nox4/p22phox complex is the major NAD(P)H oxidase complex in human endothelial cells (14). Therefore, we focused in this study on this NAD(P)H complex.
Whether Nox4 is involved in strain-dependent ROS formation in endothelial cells is not known. Furthermore, most published in vitro studies focused on short-term effects of cyclic strain on cellular ROS formation.
Therefore, we tested the impact of long-term cyclic strain on endothelial ROS formation and the expression of the major endothelial NAD(P)H oxidase subunit, Nox4.
Materials and Methods
Cell culture
All cell culture reagents were purchased from Invitrogen (Karlsruhe, Germany). The chemicals were from Sigma-Aldrich (Munich, Germany) unless otherwise specified. Primary cultures of human umbilical vein endothelial cells (HUVEC) were isolated using collagenase II as previously described (50). In order to minimize variations in primary cultures, at least three umbilical cords were used for each cell preparation. The isolated human endothelial cells were pooled and cultured on gelatin-coated plates in medium M199 supplemented with 10% fetal calf serum and 1% growth supplement. The ethical review board of the Medical Faculty of the Technical University, Dresden approved this study (EK124082003). The investigation was conducted according to the principles of the Helsinki Declaration. The human microvascular endothelial cell line (HMEC-1) (2) was kindly provided by Francisco J. Candal. The HMEC-1 cells were cultured in Endothelial Cell Growth Medium 2 (EGMV2) purchased from PromoCell (Heidelberg, Germany).
Application of cyclic strain
The Flexercell FX-4000 system with loading posts (Flexcell International Corporation, Hillsborough, NC) was used for the cell equibiaxial strain experiments. Human umbilical vein endothelial cells (HUVEC, passage 1) were cultured on silicone elastomer-bottomed and collagen type I-coated 6-well plates. After reaching confluence, endothelial cells were incubated in serum-free medium (Endothelial cell medium, PAA, Kolbe, Germany) overnight. Next, HUVECs were subjected to cyclic mechanical strain with an elongation of 2–18% at 1 Hz for up to 24 h. An internal control accompanied each cell culture dish from the same human endothelial cell preparation without application of cyclic strain.
Quantification of mRNA expression by RT-PCR and real-time PCR
After stimulation with cyclic strain, RNA was isolated using peqGOLD TriFast (peqlab Biotechnologie GmbH, Erlangen, Germany). In reverse transcriptase (RT) reactions, equal amounts of total RNA (500 ng) from HUVECs were incubated for 3 min at 70°C, and subsequently reverse transcribed into cDNA using random hexamer primers and SuperScript III Reverse Transcriptase (Invitrogen, Karlsruhe, Germany) for 1 h at 42°C. The Nox4 and p22phox mRNA expression was determined by PCR using specific primers as previously described (10, 44, 50). Furthermore, we used the following primer pairs for amplification of catalase (sense: 5′-ACA TGG TCT GGG ACT TCT GG-3′; antisense: 5′-GTC CAG GAG GGG TAC TTT CC-3′) and manganese superoxide dismutase (MnSOD) (sense: 5′-CGT CAC CGA GGA GAA GTA CC-3′; antisense: 5′- AGT CAC GTT TGA TGG CTT CC-3′). The mRNA expression was normalized to 18S rRNA.
For real-time PCR analyses of Nox4 and Cu/ZnSOD mRNA, TaqMan® gene expression assays (Hs00418356_m1 for Nox4, HS00916176_m1 for Cu/ZnSOD, and eukaryotic 18S rRNA endogenous control, VIC/MGB probe, primer limited, for 18S rRNA) were used according to the manufacturers protocol (Applied Biosystems, Darmstadt, Germany) in a LightCycler®System (Roche Applied Science, Mannheim, Germany).
Protein isolation and Western blot analysis
A polyclonal Nox4 antibody was generated by immunizing rabbits using human Nox4-specific peptides and subsequent purification with affinity chromatography. After static culture or application of cyclic strain, cells were washed twice with PBS, lysed in 50 mmol/L Tris-HCl (pH 7.0), 1% SDS, Protease Inhibitor Cocktail (P8340, Sigma-Aldrich, Munich, Germany), incubated for 10 min at 95°C and centrifuged for 10 min at 6000 g and 4°C. The protein concentration was determined with BCA Protein Assay Reagent (Perbio Science, Bonn, Germany). Proteins (20 μg/lane) were separated by SDS-PAGE (10%) and transferred to Protran® membranes (Schleicher & Schuell, Dassel, Germany). Membranes were incubated with primary antibodies against Nox4 (1:1,000; generated as described above), p22phox (1:250; Santa Cruz Biotechnology Inc., Heidelberg, Germany), eNOS and Heme oxygenase-1 (HO-1) (1:2,500, 1:250; BD Biosciences Transduction lab (Heidelberg, Germany), manganese superoxide dismutase (MnSOD) (1:1,000; Calbiochem, Bad Soden, Germany), Cu/Zn superoxide dismutase (Cu/ZnSOD) (1:1,000; Novocastra Laboratories, Newcastle, UK), or GAPDH (1:2,500; Hytest, Turku, Finland). The primary antibodies were subsequently detected with secondary horseradish peroxidase conjugated anti-rabbit IgG or anti-mouse IgG (Zytomed, Berlin, Germany). The protein expression was detected with Western Lighting Chemiluminescence Reagent Plus (PerkinElmer LAS, Rodgau, Germany) and quantified using AIDA Image Analyzer software (Raytest, Berlin, Germany).
Nox4 promoter deletion studies
The promoter region of human Nox4 gene containing 1251 bp was cloned into pCR-II-TOPO vector (Invitrogen) using PCR primers corresponding to the genomic sequence. 5′-terminally deleted Nox4 promoter fragments of 725 bp and 428 bp were generated and cloned into luciferase reporter vector pGL3-basic (Promega, Mannheim, Germany). All constructs were confirmed by DNA sequencing. HMEC-1 cells were transfected with pRL-CMV as the control plasmid and pGL3-1251, pGL3-724 or pGL3-428 Nox4 promoter constructs, or the pGL3-basic vector control using TransFectin Lipid Reagent (Bio-Rad Laboratories, Munich, Germany) and subsequently exposed to cyclic strain of 12%. The Nox4 promoter activity was determined 24 h after transfection by dual-luciferase assay (Promega) as described by the manufacturer, corrected to transfection efficiency of pRL-CMV and normalized to control vector pGL3-basic.
The computer-based analysis of potential transcription factor binding sites was performed with the Transcription Element Search System (
Detection of reactive oxygen species
Confluent HUVECs were subjected to cyclic strain for 24 h in medium M199 (1% FBS, 0.1 ECGS) without phenol red. After static culture or application of cyclic strain, ROS formation was measured using electron paramagnetic resonance (EPR) spectroscopy. After the experiment, HUVECs were detached with trypsin/EDTA and collected by centrifugation (300 g for 10 min at room temperature). The cell pellets were resuspended in modified HEPES-Tyrode buffer (pH 7.4). For the incubation, we used identical cell numbers. Superoxide production was measured by 30 min incubation of cell-permeable spin probe 1-hydroxy-3-methoxycarbonyl -2,2,5,5-tetramethylpyrrolidine (CMH, Noxygen Science Transfer & Diagnostics, Enzlingen, Germany) at a final concentration of 1 mmol/L. Oxidation of the spin probe CMH by ·O2 − forms stable 3-carboxymethoxyl nitroxide radicals (CM·). CMH is more sensitive than CPH to detect superoxide anions in cell homogenates (9). Production of H2O2 was measured by co-oxidation of cell impermeable 1-hydroxy-3-carboxy-2,2,5,5-tetramethylpyrrolidine (CPH, Noxygen Science Transfer & Diagnostics) in horseradish peroxidase (HRP)-acetamindophenol (AAP) reaction (9). Cell homogenates were incubated with 1 U/ml HRP, 1 mM AAP, 100 μM NADPH, 0.2 mM DTPA, 50 U/ml superoxide dismutase (SOD), and 500 μM CPH in HEPES-Tyrode buffer containing 25 μm deferroxamine and 5 μM DETC. The rate of H2O2 formation was determined by measuring catalase (Cat, 50 μg/ml) inhibited nitroxide generation.
The cell suspension was immediately transferred into a 50 μl EPR capillary, which was placed into the EPR cavity of a Bruker 300E EPR spectrometer. EPR settings were as follows: center field, 1.9901 g; microwave power, 1 mM; modulation amplitude, 10 G; receiver gain, 1 × 103; sweep time, 10 s.
Upon reaction with ROS, CPH is transformed into a stable CP· radical (3-carboxy-proxyl). Therefore known concentrations of 3-carboxy-proxyl were used to quantify the levels of radical forms of spin probes. Measurements were performed by Noxygen Science Transfer & Diagnostics GmbH.
In addition, ROS formation was assessed by fluorescence using dihydroethidium (DHE, Molecular Probes, Invitrogen). Endothelial cells were exposed to cyclic strain. After 24 h, 5 μmol/L DHE was added and cells were incubated at 37°C for 30 min in medium M199 without phenol red in the dark. Cells were washed in phosphate-buffered saline, detached with trypsin/EDTA and collected by centrifugation (300 g for 10 min at room temperature). DHE fluorescence was quantified using a FluoStar Optima (BMG, Offenburg, Germany; λex: 535 nm, λem: 610 nm).
In order to determine ·O2 − formation during exposure of cyclic strain to endothelial cells, further measurements were performed by cytochrome c assay as previously described (10). In order to determine the superoxide anion-dependent part of ROS formation, superoxide dismutase (SOD: 40 μg/mL, Sigma-Aldrich) were added 1.5 h before the measurement using cytochrome c (40 μmol/L). While HUVECs were continuously exposed to cyclic strain, reduction of cytochrome c was determined spectrophotometrically at 550 nm every additional 30 min in all dishes.
Amplex red assay
H2O2 was measured using Amplex red/horseradish peroxides fluorescence assay (Invitrogen). Amplex Red (50 μM) and horseradish peroxidase (0.1 U/ml) were added to colorless medium (M199). After 60 min, the H2O2-dependent oxidation of Amplex Red was measured in cell culture supernatant using a FluoStar Optima (BMG, Offenburg, Germany; λex: 544 nm, λem: 590 nm).
Nitric oxide determination
Nitrite, the major metabolite of nitric oxide (NO) in aqueous solution was measured as previously described (15). In brief, HUVEC supernatant was removed and Griess reagent containing sulfanilamide and N-(1-naphthyl) ethylenediamine was added to each tube for diazotization of sulfanilic acid by NO. After 10 min incubation at room temperature for full color development, the nitrite release was measured as the increase in absorbance at 540 nm and related to a standard curve generated by known concentrations of nitrite.
Statistics
Data are shown as mean ± SEM. Each experiment was performed at least three times. Statistical analysis was performed using Student's t test or ANOVA followed by Holm–Sidak test. p Values <0.05 were considered statistically significant.
Results
Time-dependent regulation of endothelial NAD(P)H oxidase and antioxidative enzymes by cyclic strain
In HUVECs exposed to cyclic strain, the mRNA expression of the major endothelial catalytic NAD(P)H oxidase subunit Nox4 and the membrane-bound subunit p22phox was downregulated in a time-dependent manner (Fig. 1A). A significant downregulation of both genes was observed after 24 h of 12% cyclic strain. Next we analyzed the time-dependent (1, 3, 7, 24 h) regulation of the antioxidative enzymes manganese superoxide dismutase (MnSOD), copper-zinc superoxide dismutase (Cu/ZnSOD), and catalase (Cat). MnSOD mRNA expression was induced after 3 h, but decreased after 7 h of 12% cyclic strain. Long-term cyclic strain (24 h) of 12% significantly inhibited the mRNA expression of all three enzymes significantly (Fig. 1B).

Impact of cyclic strain on ROS formation and Nox4 expression in human endothelial cells
Next, we determined the contribution of cyclic strain to ROS production. HUVECs were exposed to cyclic strain of 2, 5, 8, 12, and 16% for 24 h. We observed a strain-dependent downregulation of intracellular ROS formation with a maximum effect at 12% using DHE fluorescence (Fig. 2A). ROS formation returned to baseline after exposure of 16% cyclic strain. In line with these findings, Nox4 mRNA expression started to decrease at 2% and was strongly inhibited at 12% cyclic strain, while at 16% and 18% Nox4 expression returned to baseline (Fig. 2A).
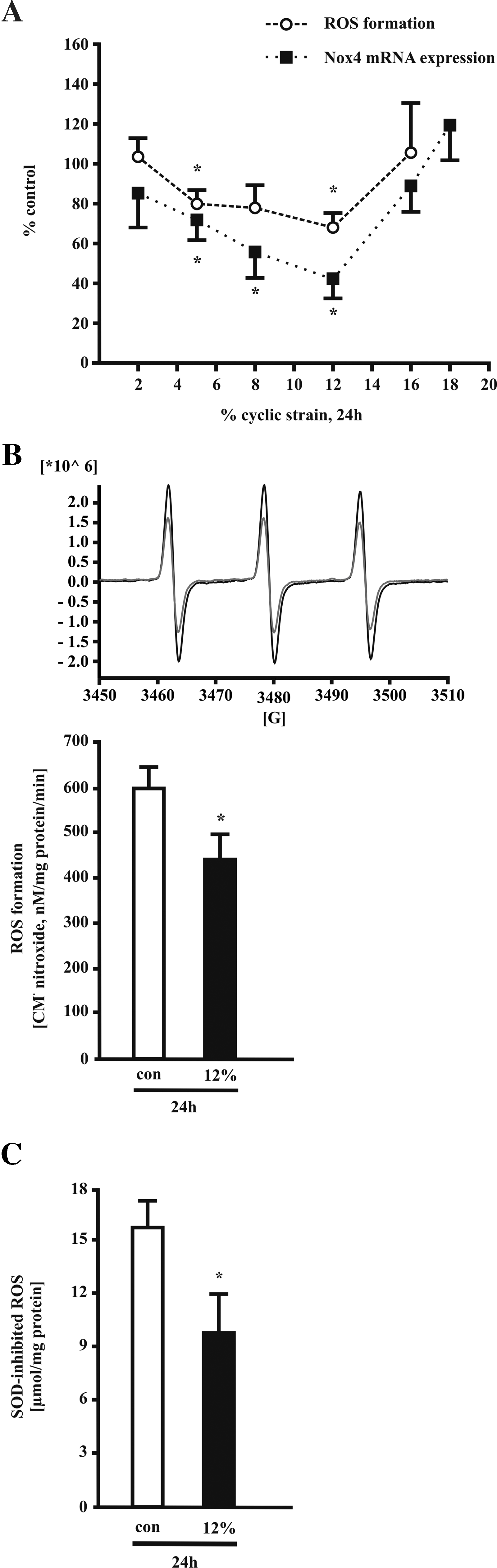
The downregulation of ROS formation by 12% long-term cyclic strain was confirmed using further methods. First, electron paramagnetic resonance (EPR) spectroscopy was used to detect endothelial superoxide anion production with the cell-permeable spin probe CMH. Using this method, the downregulation of ROS formation after 24 h of cyclic strain (12%) was corroborated (Fig. 2B). Second, measurement of ROS by SOD/CAT-inhibited cytochrome c assay showed a similar reduction of ROS formation in response to 12% cyclic strain for 24 h (Fig. 2C).
Because a vascular H2O2 release via Nox4 is also reported, we measured the H2O2 production in response to cyclic strain in endothelial cells. Application of 12% cyclic strain for 24 h results in a trend to increased H2O2 formation as measured by EPR in intact cells with the spin probe CPH (Fig. 3A). In parallel extracellular ·O2 − production tends to decrease (Fig. 3B). Use of rotenone (5 μM) and apocynin (500 μM) had no effect on strain-dependent ROS and H2O2 formation measured by EPR (Fig. 3C) and Amplex Red fluorescence (Fig. 3D).
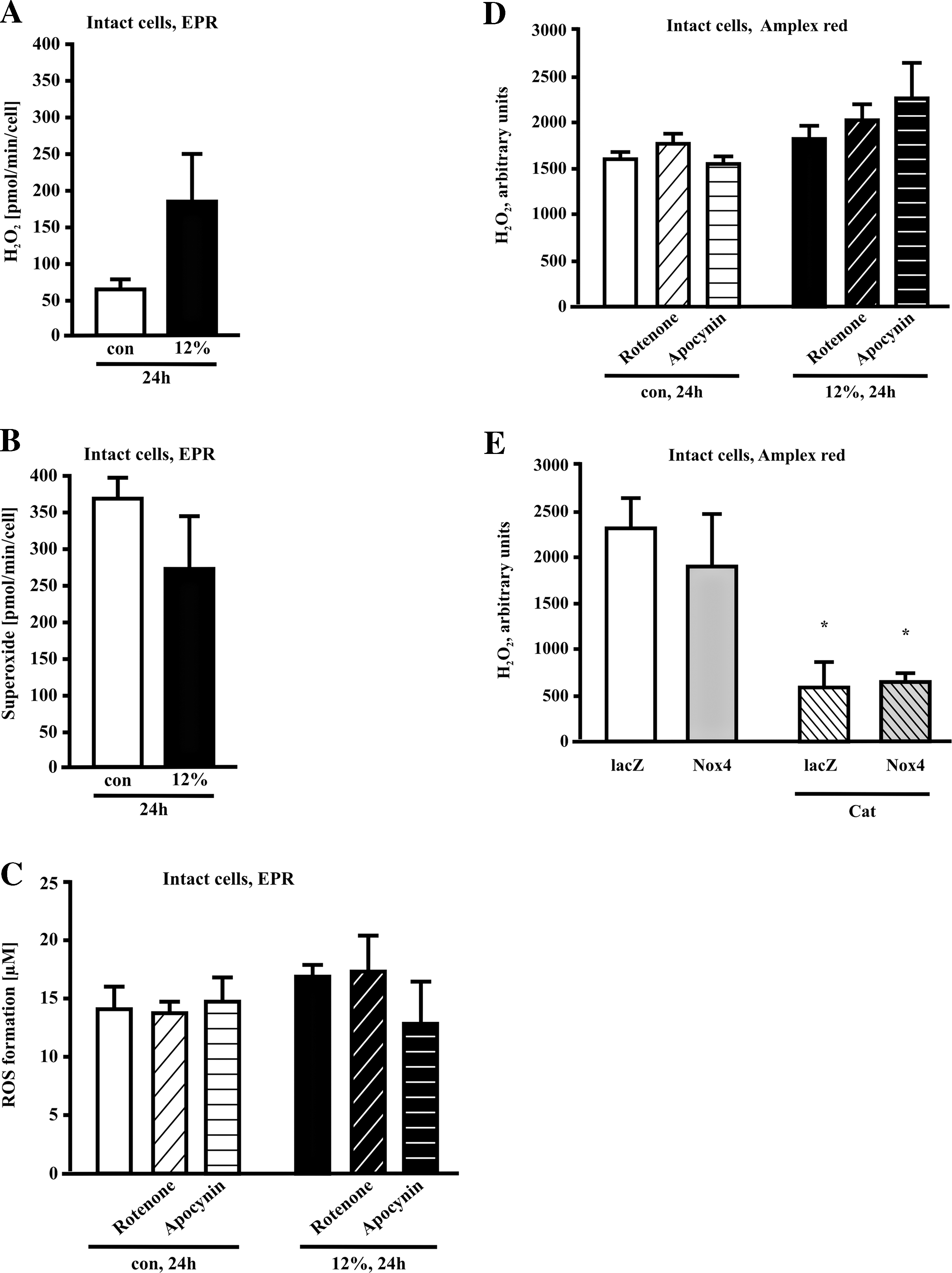
Next, we analyzed the impact of Nox4 overexpression on H2O2 generation in human endothelial cells. Using the Amplex Red assay, we were not able to detect a significant difference in H2O2 production of endothelial cells overexpressing human Nox4 compared to endothelial cells expressing a lacZ-control vector (n = 3). The Amplex Red fluorescence signal of human endothelial cells under basal conditions and in cells overexpressing Nox4 could be blocked in a similar way with catalase (50 μg/ml), (Fig. 3E).
Strain-dependent regulation of endothelial NAD(P)H oxidase and antioxidative enzymes by long-term cyclic strain
In HUVECs exposed to cyclic strain Nox4 mRNA was strain-dependently downregulated (Figs. 2A and 4A). In contrast p22phox mRNA was downregulated after 5% and 12% cyclic strain to the same degree (Fig. 4A). Next we analyzed Nox4 protein expression after 5% and 12% long-term cyclic strain (24 h) using a novel Nox4 antibody. While 5% cyclic strain had no effect, 12% cyclic strain significantly downregulated Nox4 protein expression (Fig. 4B). In contrast to Nox4, p22phox protein expression was not significantly regulated after application of 12% cyclic strain for 24 h (Fig. 4B). Of note, all three antioxidative enzymes MnSOD, Cu/ZnSOD, and Cat were significantly downregulated by 5% and 12% of long-term cyclic strain at mRNA level (Fig. 4C).

The downregulation of MnSOD by cyclic strain (12%, 24 h) could be confirmed on the protein level (Fig. 4D). Due to the lack of a specific Cat antibody, we were not able to confirm this downregulation at the protein level. However, significant downregulation of Cu/ZnSOD protein expression was shown after application of 12% cyclic strain for 24 h (Fig. 4D). In addition, the protein expression of HO-1 and the lectin-like oxLDL receptor LOX-1 is not affected by cyclic strain (data not shown).
Regulation of Nox4 promoter by cyclic strain
Next, we generated Nox4 promoter deletion constructs to study the effect of cyclic strain on Nox4 promoter activity. HMEC-1 cells were transfected with different Nox4 promoter constructs (Fig. 5A) and exposed to cyclic strain of 12% for 24 h. Nox4 promoter activity was quantified as luciferase reporter gene activity. Cyclic strain reduced Nox4 promoter activity of constructs pGL3-N4P + 239/-725 and pGL3-N4P + 239/-1251 in comparison to static controls transfected with the same constructs (Fig. 5B). This provides further evidence that the downregulation of Nox4 by cyclic strain is mediated on the transcriptional level.
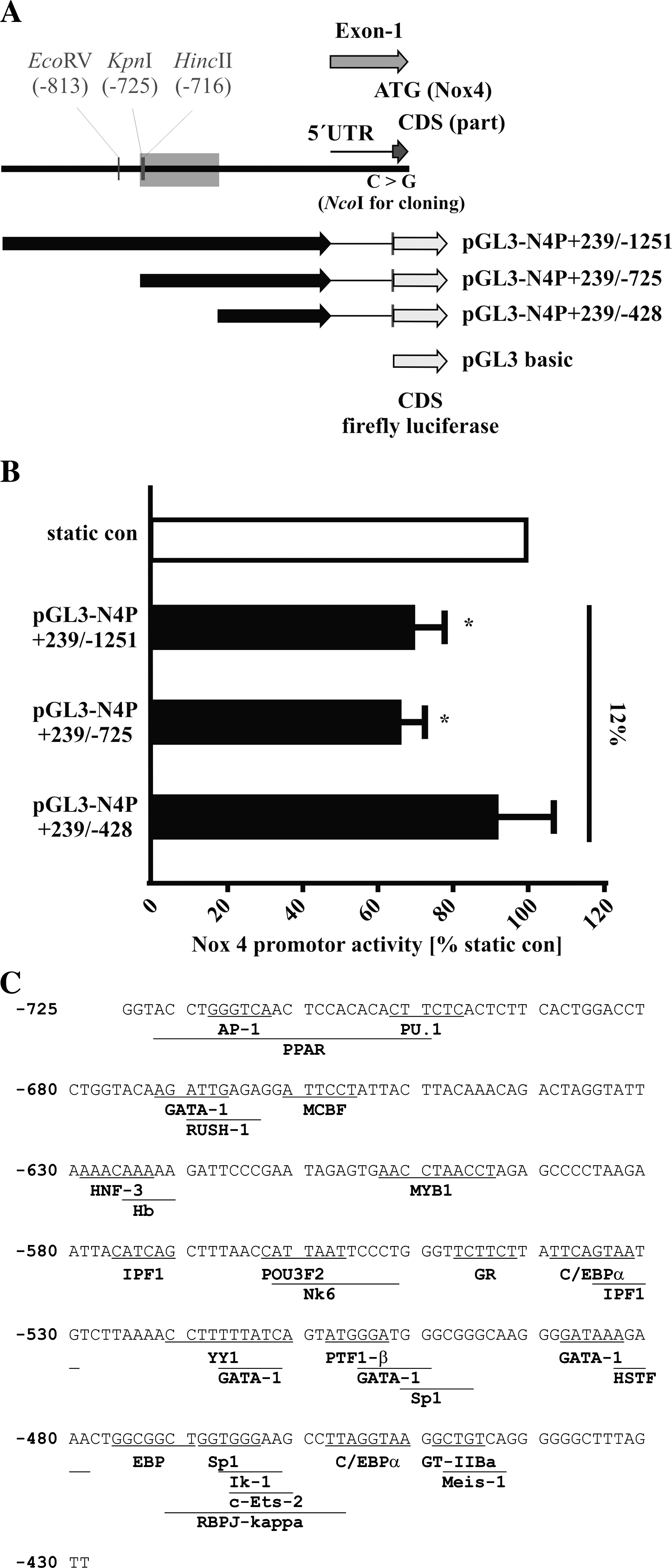
In contrast, cyclic strain did not reduce the activity of Nox4 promoter construct pGL3-N4P + 239/-428. Therefore, the transcription factor binding site responsible for the downregulation of Nox4 by cyclic strain is most probably located between position −428 and −725 bp upstream of the transcription start site. Computer-based analysis of possible transcription factors using two independent programs (Fig. 5C) resulted in potential transcription factor binding sites of six factors known to be regulated by cyclic strain, like C/EBP (27, 55), RBPJκ (38), SP1 (24), AP1 (59), PU.1 (60), or PPAR (55). In addition, potential bindings sites of 18 factors currently unrelated to cyclic strain response were predicted (GATA-1, HNF-3, RUSH-1, MCBF, Hb, MYB1, IPF1, POU3F2, Nk6, GR, YY1, PTF1-β, HSTF, EBP, Iκ-1, C-ETS-2, Meis-1, and GT-IIBa).
Cyclic strain-dependent release of nitric oxide by human endothelial cells
Because ROS and nitric oxide (NO) formation are regulated in a tight balance, we analyzed the NO release in our experimental system. Application of 12% cyclic strain caused a time-dependent release of NO with a maximum after 24 h (Fig. 6A). A strain-dependent upregulation of NO release was observed starting at 5% cyclic strain (24 h) (Fig. 6B). Long-term application of cyclic strain (12%, 24 h) was accompanied by a threefold increase in endothelial nitric oxide synthase (eNOS) protein expression (Fig. 6C).

Impact of NO inhibition on cyclic strain-dependent downregulation of Nox4
Finally, we analyzed the impact of increased NO release in response to cyclic strain on Nox4 downregulation. NO-Synthase was blocked by the inhibitor L-NAME (500 μM). L-NAME per se had no effect on Nox4 expression, while the combination of L-NAME with cyclic strain partially blocked stretch-dependent downregulation of Nox4 (Fig. 7A). In contrast, stimulation of HUVECs with the NO donor DETA-NO did not change Nox4 expression (Fig. 7B). The strain-dependent NO production can not be altered by SOD/CAT or apocynin, while rotenone partially blocked NO formation (data not shown).

Discussion
This is the first study describing a regulation of the major NAD(P)H oxidase subunit Nox4 by cyclic strain in endothelial cells. While short-term application of 12% had no effect on Nox4 expression, long-term application of 5%–12% cyclic strain resulted in a strain-dependent downregulation of Nox4 mRNA expression that is normalized at higher cyclic strain (16%). Using a novel specific Nox4 antibody, we were able to confirm the downregulation at the protein level. This downregulation of Nox4 is regulated on the transcriptional level, because the promoter activity of human Nox4 deletions starting at position −1251 and −725 is downregulated by cyclic strain in endothelial cells as well. This is the first report analyzing Nox4 promoter constructs in response to cyclic strain. Since the activity of a further deletion construct starting at −428 is not affected by cyclic strain, the binding site of the transcription factors mediating this downregulation is most probably located between position −725 and −428 of the human Nox4 promoter. The mechanism of cyclic strain-induced decrease in ROS production might include downregulation of Nox4 on the transcriptional level as shown in this study. The identified strain responsive region of the human Nox4 promoter contains binding sites of several transcription factor known to be regulated by cyclic strain including C/EBP (27, 55), RBPJκ (38), SP1 (24), AP-1 (59), PU.1 (60), or PPARγ (55). Other predicted candidates include GATA-1, RUSH-1, MCBF, HNF-3, Hb, MYB1, IPF-1, POU3F2, Nk6, GR, YY1, PTF1-β, HSTF, EBP, Iκ-1, c-Ets-2, Meis-1, and GT-IIBa). Which transcription factors are responsible for this downregulation in response to cyclic strain will be the focus of future studies. The observed upregulation of NO release could at least partially contribute to this downregulation of ROS formation in response to long-term cyclic strain as well. Our data suggest a beneficial effect of physiological values of cyclic strain on vascular function. This is in agreement with the reduced Nox4 expression, ROS formation, and improved endothelial function in response to increased blood flow and subsequent biomechanical stimulation of vascular cells in vivo by physical exercise training (1).
Because Nox4 is forming an NAD(P)H oxidase complex with p22phox, we analyzed the regulation of this subunit in response to cyclic strain as well. The p22phox mRNA expression was not affected by short-term application, but downregulated in response to long-term application of cyclic strain of 24 h. Endothelial cells treated with the NAD(P)H oxidase inhibitor diphenyleneiodoniumchloride (19) or a dominant-negative mutant of Rac (58) resulted in reduced superoxide anion generation in response to cyclic strain. In a previous study, cyclic strain increased the endothelial mRNA expression of p22phox (35). Recently, it has been shown that physiological cyclic strain (5%) increases but pathological strain (20%) decreases endothelial cell survival and tubulogenesis by a pathway involving ROS that is produced by an NAD(P)H oxidase complex containing p22phox (26). Even while the role of p22phox in this later study was not accompanied by a determination of its expression in response to cyclic strain, it further supports an important role of NAD(P)H oxidase complexes in response to biomechanical forces. Using human endothelial cells, we found that exposure to laminar shear stress (30 dyn/cm2) induced a transient upregulation of p22phox mRNA expression after 12 h, which returned to baseline after 24 h. In these samples exposed to laminar shear stress, the p22phox protein expression did not change after 24 h as well (10).
Here, we observed a strain-dependent downregulation of superoxide anion radical formation in response to long-term cyclic strain reaching a maximum inhibitory effect at 12%. The same regulation was observed for Nox4 mRNA and protein expression. By EPR measurements with the cell-permeable spin probe CMH, we can confirm the downregulation by 12% long-term cyclic strain. Using cell-impermeable spin probe CPH we detected a trend to increased H2O2 generation combined with a trend to decrease in ·O2 − formation. These differences of intercellular vs. extracellular ROS detection in combination with the observed strain-dependency of ROS formation might explain the different results reported by different groups after application of cyclic strain. Cyclic strain for up to 6 h increased superoxide production (35, 58), but after 24 h formation of reactive oxygen species returned to control levels (19). In vitro cultures of vascular smooth muscle cells exposed to 6% cyclic strain failed to induce production of superoxide anion, whereas 10% cyclic strain induced superoxide anion release (19, 20). Furthermore, while application of cyclic strain of 10% and 20% to vessels ex vivo activated ROS generation, cyclic strain of 5% failed to induce ROS production (30). Oxidative stress induced by 6 h cyclic strain could be prevented by the mitochondrial inhibitor rotenone, whereas inhibitors of NO synthase, xanthine oxidase, or NAD(P)H oxidase had no effect (4). However we found no influence of rotenone or apocynin-inhibited enzymes in cyclic strain-dependent (12%) reduction of ROS formation.
Short-term application of bovine aortic vascular smooth muscle cells cyclic strain (10%) stimulated NAD(P)H oxidase activity after 15 min (6). In murine vascular smooth muscle cells, cyclic strain resulted in a rapid ROS formation and p47phox membrane translocation that is followed by an increase in Nox1 transcripts (18). Short-term (2 h) cyclic stretch (10%) increased ROS generation and translocation of p47phox to the membrane in sheep pulmonary arterial smooth muscle cells (34). In addition, hypertension and high pressure induced phosphorylation and translocation of p47phox resulting in activation of NAD(P)H oxidase (40, 56). Therefore, the Nox1- and p47phox-containing NAD(P)H oxidase complex appears to be crucial for the ROS formation in response to cyclic strain in vascular smooth muscle cells across various species. Application of cyclic strain increased ROS production in pulmonary epithelial cells after 2 h, returning to baseline after 24 h (5). In neonatal rat ventricular myocytes, cyclic strain caused an amplitude-related increase in ROS formation (43). Furthermore, cyclic stretch induced apoptosis in porcine retinal pericytes by activation of reactive oxygen species-c-Jun NH2-terminal kinase-caspase cascades (53). In summary, most studies in nonendothelial cells show an increase in ROS formation in response to cyclic strain. Increasing evidence support a crucial role of Nox4-derived ROS in signal transduction and differentiation (12, 14, 42). Therefore, feedback loops could be disrupted by either Nox4 or catabolic enzyme decreases and thus has an opposite effect on each. The downregulation of antioxidative enzymes could represent such an adaptive response to the decreased generation of reactive oxygen species after long-term cyclic strain. As radicals produced by Nox4 seem to be more important as intracellular signaling molecules (45) than to generate an extracellular oxidative burst, we speculate that regulation of Nox4 by cyclic strain is important for adaptation of endothelial cells to strain.
However, less is known about the impact of biomechanical forces on H2O2 generation in endothelial cells. Our data show a trend to increased H2O2 formation between static controls and cells exposed to 12% cyclic strain for 24 h. However, we were not able to detect difference in H2O2 production of endothelial cells overexpressing Nox4, compared to endothelial cells expressing a lacZ-control vector. In previous studies, overexpression of Nox4 in HEK293 cells resulted in elevated H2O2 but failed to induced O2- production (33, 46). In Nox4-depleted rat aortic smooth muscle cells, production of H2O2 was significantly lower while basal superoxide production was not affected compared to control cells (9). However in a previous study we could show increased O2- production by Nox4 overexpression in human endothelial cells (14). Cyclic strain elevated H2O2 release after 6 h to 24 h in porcine aortic endothelial cells (21). In contrast, long-term application of high laminar shear stress caused downregulation of superoxide anion formation in HUVEC (10). Combination of flow conditions with a 6% cyclic stretch value did not affect NAD(P)H oxidase activity induction in bovine aortic endothelial cells (47).
Reactive oxygen species derived from xanthine oxidase or cytochrome P-450 could contribute to the observed changes in ROS formation in response to cyclic strain. Moreover, xanthine oxidase can generate H2O2 (28) and rat xanthine oxidase activity is increased after exercise (16). Xanthine oxidase is also responsible for increased H2O2 generation by oscillatory shear stress in BAEC (36). However, in previous studies showing a transient upregulation of ROS formation by short-term application of cyclic strain, inhibitor studies do not support a major role of xanthine oxidase in the regulation of reactive oxygen species in response to cyclic strain (4, 58). The amount of ROS generated by cytochrome P-450 might represent a physiologically relevant, but smaller portion of the endothelial ROS production (11). Because endothelial cells in vivo are exposed to shear stress and cyclic strain simultaneously, complex interactions of both biomechanical forces might occur and to regulate different ROS producing enzymes.
Each cell has developed antioxidative defense mechanisms. Initial studies described an upregulation of endothelial SOD activity by cyclic strain (7). The heterogeneity in response to cyclic strain might include the endothelial cell type studied. HUVECs of higher passages increased intracellular H2O2 levels, catalase and SOD activity in response to cyclic strain, while in human aortic endothelial cells the opposite effects were observed (52). In the present study, we observed a downregulation of antioxidative enzymes MnSOD, Cu/ZnSOD, and catalase in response to cyclic strain of 12% for 24 h. Therefore, the reduced ROS formation in response to cyclic strain in our study cannot be explained by an upregulation of antioxidative enzymes. This might rather reflect an adaptation to the reduced ROS generation in response to long-term cyclic strain in our cells.
In our study, the reduced ROS formation was accompanied by an increase in NO release and eNOS expression after long-term cyclic strain of 12%. Recently, NO production was similarly stimulated by 5% cyclic strain, but inhibited by 20% cyclic strain in a human endothelial cell line (26). Several other studies described in agreement with our results an increased eNOS mRNA expression, promoter activity and functional activity in bovine endothelial cells and human endothelial cell lines (54, 62). We found a partial inhibition of strain-dependent Nox4 downregulation by eNOS inhibitor L-NAME. In contrast DETA-NO was not able to reduce Nox4 expression in endothelial cells. Therefore we conclude that eNOS is involved in Nox4 regulation but a NO donor alone is not sufficient to reduce Nox4 expression. Furthermore, rotenone treatment resulted in a reduced strain-dependent NO production. Thus, a direct scavenging of NO by superoxide anion radicals is not likely to be solitary responsible for increased eNOS activity.
Nevertheless, the observed upregulation of NO release and downregulation of ROS formation suggest a protective impact of long-term cyclic strain of up to 12% in primary cultures of human endothelial cells. In line with this hypothesis, application of cyclic strain by pulse perfusion can improve endothelial function (31).
In conclusion, physiological levels of cyclic strain downregulate Nox4 expression and superoxide anion formation. This novel mechanism might contribute to a vasoprotective balance between NO and superoxide anions in response to physiological mechanical stimulation of endothelial cells.
Footnotes
Acknowledgments
We thank Melanie Brux for her excellent technical assistance and Bruno Fink for the EPR measurements.
This work was supported by the German Federal Ministry of Education and Research program, NBL3, of the University of Technology Dresden (PhD program Metabolism and Endothelium to C.G.; Professorship of Vascular Endothelium and Microcirculation to H.M.) the MeDDrive program of the Medical Faculty Carl Gustav Carus of the University of Technology Dresden, Germany (to C.G. and W.G.), the Doktor Robert Pfleger Foundation, Bamberg, Germany (to H. M. and W.G) and the DIGS-BB program (to A.A.)
Author Disclosure Statement
No competing financial interests exist.