
Abstract
With the evolution of fish, systems appeared for the disposal of the hemoglobin (Hb) that was inevitably released from erythrocytes. Thus, a plasma protein that bound free Hb with great affinity, haptoglobin (Hp), evolved from a protease of the innate immune system. In parallel, other proteins appeared (for example, hemopexin and α1-microglobulin), which bound and mediated the removal of free heme groups. Remarkably, Hp later disappeared in some vertebrate lineages, suggesting that it could also be disadvantageous. In the avian lineage, a soluble protein evolved, possibly from a scavenger receptor, which in some birds seems to have replaced Hp. Among mammals, multimeric forms of Hp appeared independently at two discrete times, suggesting that this form of the protein confers an advantage on the bearer, possibly by improving resistance to infection. Antioxid. Redox Signal. 12, 249–260.
Introduction
Hemoglobin-Binding Proteins
Methods of detection
Gel electrophoresis under non-denaturing conditions has been used extensively for the detection of Hb-binding proteins. With this technique, an excess of Hb is added to a serum sample, and, after electrophoresis, bands containing Hb (and Hb-binding proteins) are visualized through the peroxidase activity of Hb (10, 62). Although this method is ideal for the simultaneous analysis of many samples, it provides little information about the identity of the binding protein. This goal is better achieved by using affinity chromatography with immobilized Hb, followed by SDS/PAGE and amino acid sequencing/mass spectrometry (91).
Mammalian haptoglobin
The first Hb-binding protein was detected in human plasma in the late 1930s and was given the name haptoglobin from the Greek word haptein (to bind) (65). Sera from many animals were subsequently analyzed by using the electrophoretic method, which showed that Hp levels varied greatly among species (42, 82). Furthermore, variation among individuals also was large, reflecting the fact that Hp is an acute-phase protein (54, 82).
The primary translation product of mammalian Hp mRNA is a polypeptide of ∼45 kDa that quickly dimerizes and is proteolytically cleaved while still in the endoplasmic reticulum (27). The mature protein therefore consists of four chains; two of 9 kDa (the α-chains) and two of 35 kDa (the β-chains) (Fig. 1). We have shown that the enzyme mediating the cleavage is complement component 1r-like protein (C1r-LP) (92), the product of a gene formed by partial duplication of the gene for complement component 1r (15). In all investigated mammalian species, the α-chains and β-chains originating from the same Hp polypeptide are linked by a disulfide bond, and in most species, a disulfide bond links the α-chains of the two αβ subunits (Fig. 1A). When the first amino acid sequence of human Hp was determined, it became clear that the major part of the β-chain is related to serine proteases (SPs), such as trypsin and chymotrypsin (38). This part of Hp is therefore also referred to as the SP domain. It was also noted, however, that the serine and histidine residues of the catalytic triad had been replaced by other amino acids, making the protein proteolytically inactive. Furthermore, various biochemical experiments showed that Hb binds to a site on Hp distinct from the pseudo-protease active site (2, 43, 64, 88).
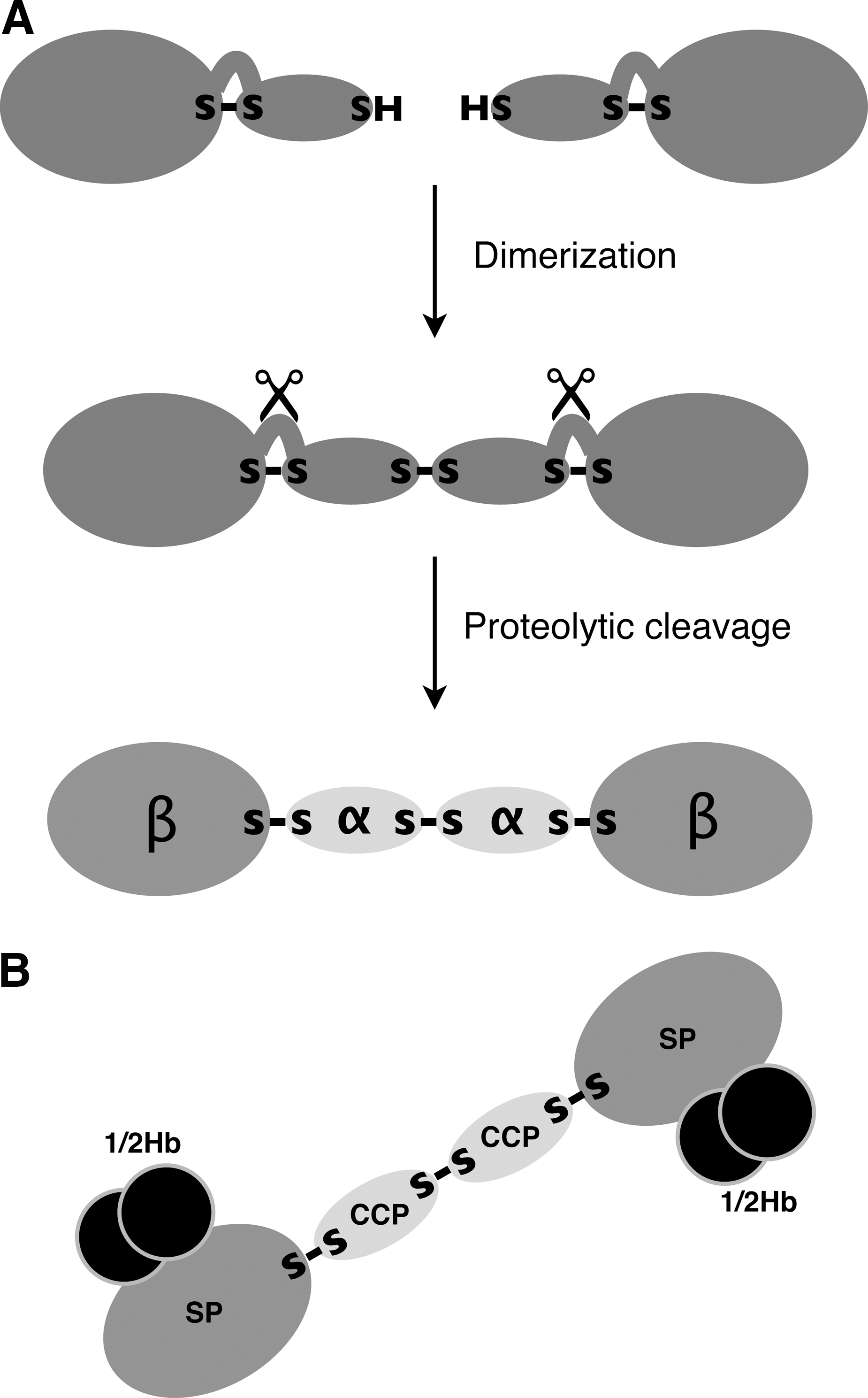
The two dimers making up an Hb molecule are bound to each other in such a way that each α-chain of a dimer binds to the β-chain of the other dimer (84). At the high Hb concentrations existing within erythrocytes, most of the Hb molecules are tetrameric, whereas on release from the cells, the protein dissociates into dimers (57). Each SP domain of Hp binds to one Hb dimer (Fig. 1B; denoted 1/2Hb), apparently to both α- and β-chains (48, 93).
To date, the function of the α-chain of Hp is unknown. Sequence analysis indicates, however, that it contains a complement-control protein (CCP) domain, also known as the short complement-like repeat or Sushi domain (81). These structures occur both in complement and in noncomplement proteins, where they mediate binding to other proteins or carbohydrates (61).
Mammalian haptoglobin variants
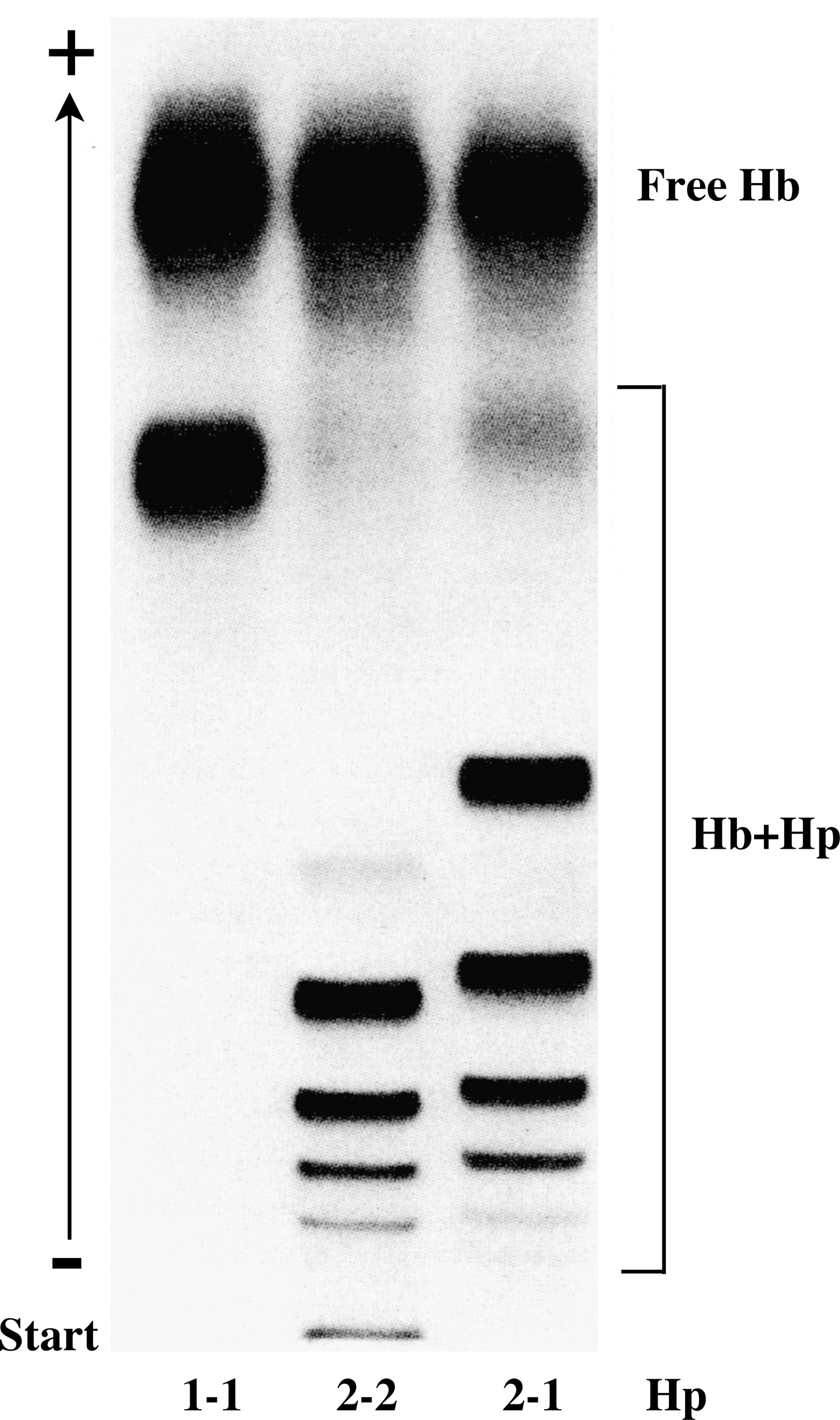
In humans, Hp exists in two major allelic forms, Hp1 and Hp2, differing in the size of the α-chain. In Hp1, the α-chain comprises one CCP domain, whereas in Hp2, it comprises two CCP domains (Fig. 2). The shorter α-chain (α1) has one free sulfhydryl group, and the longer α-chain (α2) has two free sulfhydryl groups, binding to one and two other α-chains, respectively. The two allelic forms of Hp, therefore, give rise to three different serotypes in which the Hp molecules comprise either two α1β units (Hp1-1), a variable number of α2β units (Hp2-2), or two α1β units and a variable number of α2β units (Hp1-2) (Fig. 2). On electrophoresis under non-denaturing conditions and in the absence of a reducing agent, Hp1-1 will yield a single band, and Hp2-2 and Hp1-2 will yield multiple bands (Fig. 3). Densitometric analysis of the bands shows that the most abundant forms of Hp2-2 and Hp2-1 contain four and three multiples of the αβ-subunits, respectively (88).
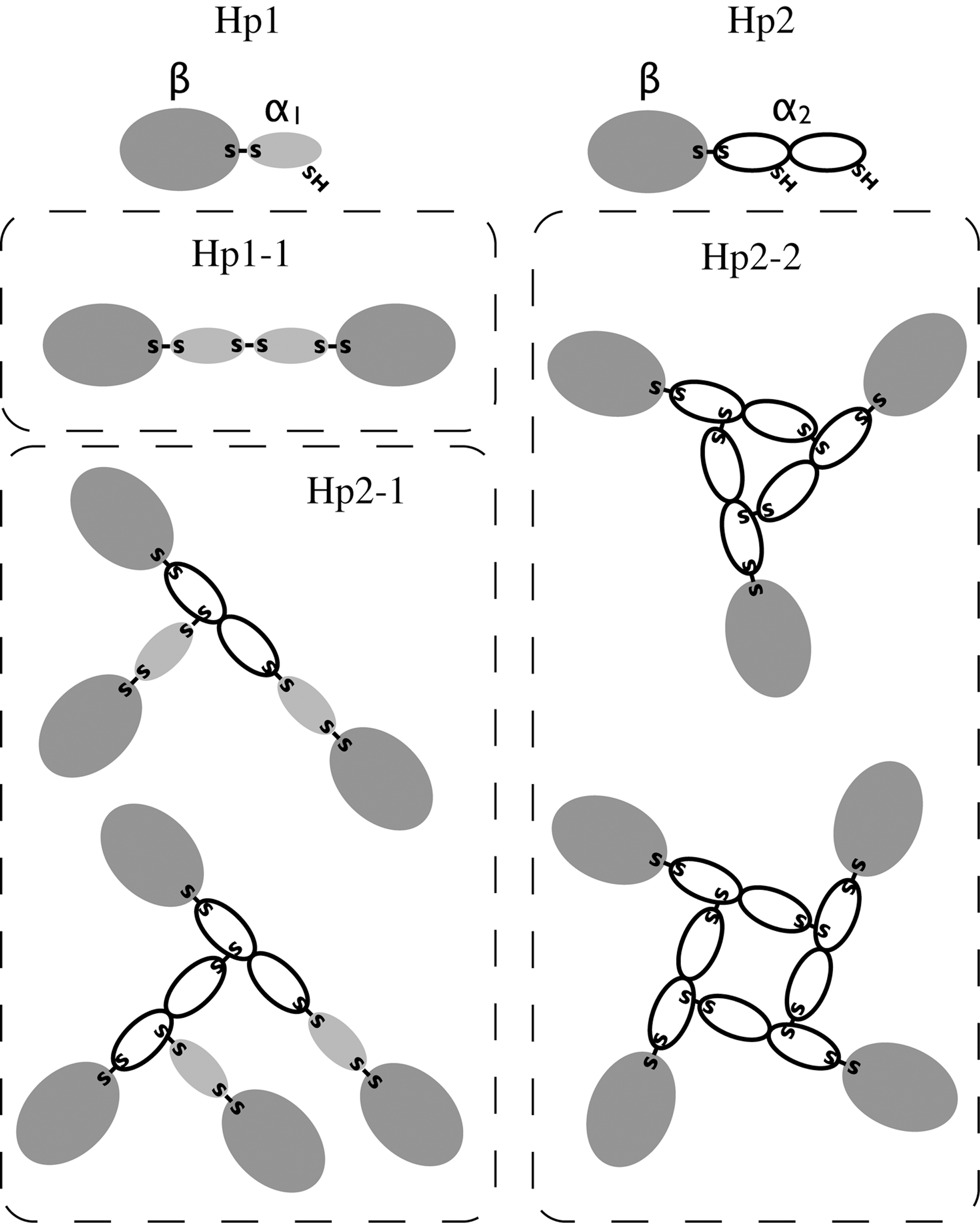
For most mammals, the electrophoretic mobility of Hp has been found to be similar to that of human Hp1-1 (13). The known exceptions are species of the suborder Ruminantia, such as cattle and sheep, for which multiple bands or large complexes have been observed (52, 82). The gene for bovine Hp (bHp) was recently identified, and the predicted amino acid sequence suggested a domain structure similar to that of human Hp2 (40, 90). The two CCP domains of bHp are essentially the same size as the human counterparts, but their amino acid sequence similarity is lower: 68% as opposed to 98% (90). The inner CCP domain of bHp contains an additional free Cys residue, which probably accounts for the high degree of cross-linking (40). Analysis of available sequences encoding Hp from sheep (90), deer (39), and goat (unpublished data) shows that their α-chains also comprise two CCP domains.
The plasma of humans and some other primates contains a protein with more than 90% amino acid sequence similarity to Hp, called Hp-related protein (Hpr). Hpr, which occurs in association with high-density lipoprotein (53), binds Hb, but unlike Hp bound to Hb, the complex is not removed from the bloodstream (60). However, Hpr bound to Hb is endocytosed by the parasite Trypanosoma brucei brucei, and once in its lysosomes, the associated lipoprotein together with Hpr causes the death of the parasite (51, 63); see also the contribution by Nielsen et al. in this Forum Review.
Evolution of the mammalian haptoglobin gene
In 1962, Smithies et al. (77) discovered an Hp-related gene (Hpr) located 2.2 kb downstream from the human Hp gene (77) and which was later found to produce Hpr, described earlier (7). Further analysis revealed that New World monkeys have only one Hp gene, whereas Old World monkeys and apes have three (49). The simplest explanation for these findings is that the original Hp gene was triplicated after the separation of the lineage leading to New World monkeys ∼30 million years ago. After the divergence of the chimpanzee lineage ∼2 million years ago, one of the Hp genes was subsequently deleted from the human genome (49).
The two major Hp alleles (Hp1 and Hp2) in the human population give rise to Hp proteins with α-chains of different lengths. Based on a comparison of Hp2 with two minor alleles of Hp1 (Hp1S and Hp1F), Maeda et al. (46) suggested that Hp2 arose by an internal duplication of the gene segment encoding the CCP domain caused by an unequal crossing-over between the two Hp1 alleles (46). This event must have occurred after the separation of the human and great ape lineages, because Hp2 does not exist in the latter (45). Ruminants have an Hp2-like gene, but apparently no Hp1-like gene. The mechanism of duplication in these animals, therefore, cannot be deduced simply by comparison of two alleles, as was done for the human Hp gene. However, a comparison of the chromosomal regions comprising bHp and human hHp2 indicates that the former gene was formed by duplication of the ancestral bHp gene segment covering a region homologous to the duplicated segment in hHp2 (90). In cow, unlike in humans, the duplication seems to have been followed by various rearrangements, suggesting a more ancient origin. The bovine Hp2-like gene has been found in species from three different ruminant families: Bovidae, Cervidae, and Giraffidae (50, 70), implying that the duplication took place in a common ancestor, probably 30 to 40 million years ago (29).
Statistical methods can be used to determine whether a protein domain has evolved under selective pressure (67, 68). By using this approach, we found that, of the first 54 codons in the CCP domains from several animals, strong statistical evidence for selection existed in only four codons, and that this selection was negative (90). This indicates that the CCP domains have not evolved to gain a new function. Possibly, their only purpose in Hp may have been to allow for the creation of large complexes, preventing the protein from diffusing from the bloodstream.
Avian hemoglobin-binding proteins
The presence of an Hb-binding protein in the plasma of different birds has been demonstrated by the use of gel electrophoresis (42). In chicken, however, the protein was detected only after the plasma level was increased by turpentine-induced inflammation (56). Subsequently, an Hb-binding protein was isolated from chicken serum, and it was noted that it bound chicken and turtle Hb, but not Hb from various mammals; in contrast, human Hp was found to bind both chicken and human Hb with great affinity (19, 55). These findings suggest that the chicken protein binds Hb in a different way from human Hp. By using the BLAST program, we analyzed the newly available chicken genome with the sequence of the β-chain of human Hp as query. The predicted protein sequences with the highest scores were then examined for the presence of Hp characteristics: an SP domain lacking the essential serine and histidine residues of the active site as well as the “histidine-loop” cysteine pair (81). No gene encoding a protein similar to mammalian Hp was found (91). By using affinity chromatography with immobilized chicken Hb (18), we isolated the Hb-binding protein from chicken plasma. Analysis with SDS-PAGE revealed a 69-kDa band, and mass-spectrometric analysis showed that it contained a protein originally identified as an inflammation-induced protein in chicken plasma (79). This protein had been isolated by Iwasaki et al. (35), who also cloned and sequenced its cDNA; they referred to the protein as 18-B in their publication but as PIT54 in the GenBank database. Furthermore, they showed that 18-B/PIT54 has antioxidative properties in that it inhibits superoxide production by stimulated phagocytes (35).
To find out the distribution of Hb-binding proteins among birds, we also analyzed the plasma of domestic goose (Anser anser) and found a protein of ∼70 kDa. A partial amino acid sequence was obtained with mass spectrometry, which indicated that the protein was an orthologue of PIT54 (91). Similarly, we investigated the serum of ostrich, a representative of paleognaths, a separate phylogenetic line of modern birds (26). This experiment revealed the presence of both Hp and an orthologue of PIT54 (91). A PIT54-like protein also has been reported in another paleognath, the emu (47). Furthermore, Hp has been found to occur in cormorant (58), a bird belonging to a different order from either chicken or goose. Our analysis of nonavian genomic sequences did not reveal any PIT54 orthologues (91). The simplest interpretation of these findings is that PIT54 evolved as a complement to Hp in the ancestors of all birds, and that the Hp gene was later deleted in the ancestor of at least one avian order.
Amphibian hemoglobin-binding proteins
Electrophoretic analysis of serum from a frog (species not specified) by Liang (42) revealed an Hb-binding protein. However, our search in the genomic database of Xenopus tropicalis (African clawed frog) for a gene encoding a protein homologous to the mammalian Hp β-chain did not reveal an Hp gene (91). Analysis of serum from X. laevis with Hb-affinity chromatography yielded a protein similar to the μ-chain of IgM (91). This protein has not previously been assigned any function, and it remains to be seen if the fact that it binds Hb in vitro is of any physiologic significance.
Reptilian hemoglobin-binding proteins
Electrophoresis has provided evidence for the presence of an Hb-binding protein in sera from snake, turtle, and lizard (42, 74). Our DNA sequence analyses indicate that no Hp gene is found in anole lizard, whereas it is present in alligator (unpublished data). Furthermore, we detected Hp in the serum of Chinese soft-shell turtle with affinity chromatography (91).
Hemoglobin-binding proteins in bony fish
Electrophoretic analysis revealed the presence of Hb-binding proteins in the plasma of some species of fish (36, 83). In a study of the acute-phase response in trout, bacteria were injected, and the change in plasma protein levels was analyzed by using 2D-gel electrophoresis. With Edman degradation, a 15-amino acid sequence similar to one found in human Hp was detected in one protein that increased in level (24). By analyzing genomic or EST sequences from several fish species, we found a gene encoding a protein with an amino acid sequence 32–34% identical to that of the β-chain of human Hp (91). The protein apparently comprises an SP domain preceded by a 16- to 22-residue extension and a putative signal peptide. Furthermore, at its C-terminal end, the extension contains a consensus cleavage sequence for subtilisin-like proprotein convertases: R-X-(R/K)-R (Fig. 4). These enzymes, which are located in the Golgi complex, seem to occur in all eukaryotic cells (75). With Hb-affinity chromatography followed by SDS-PAGE, we obtained two major protein bands with apparent molecular masses of ∼45 and 75 kDa from the serum of Takifugu rubripes (puffer fish). The proteins were identified with mass spectrometry as Hp and IgM heavy chain (μ-chain), respectively (91). The cDNA of Hp from T. rubripes was cloned, and the protein was expressed in COS-7 cells with and without an inhibitor of proprotein convertases. This experiment showed that the protein was cleaved by proprotein convertases during secretion (unpublished data).

The dimeric subunits of fish Hb are bound to each other much more strongly than those of other vertebrates (11, 21), suggesting little dissociation of Hb on hemolysis. Because Hp binds exclusively to Hb dimers (57), it is likely that only a small part of the Hb released on hemolysis in fish is captured by Hp.
Hemoglobin-binding proteins in cartilaginous fish
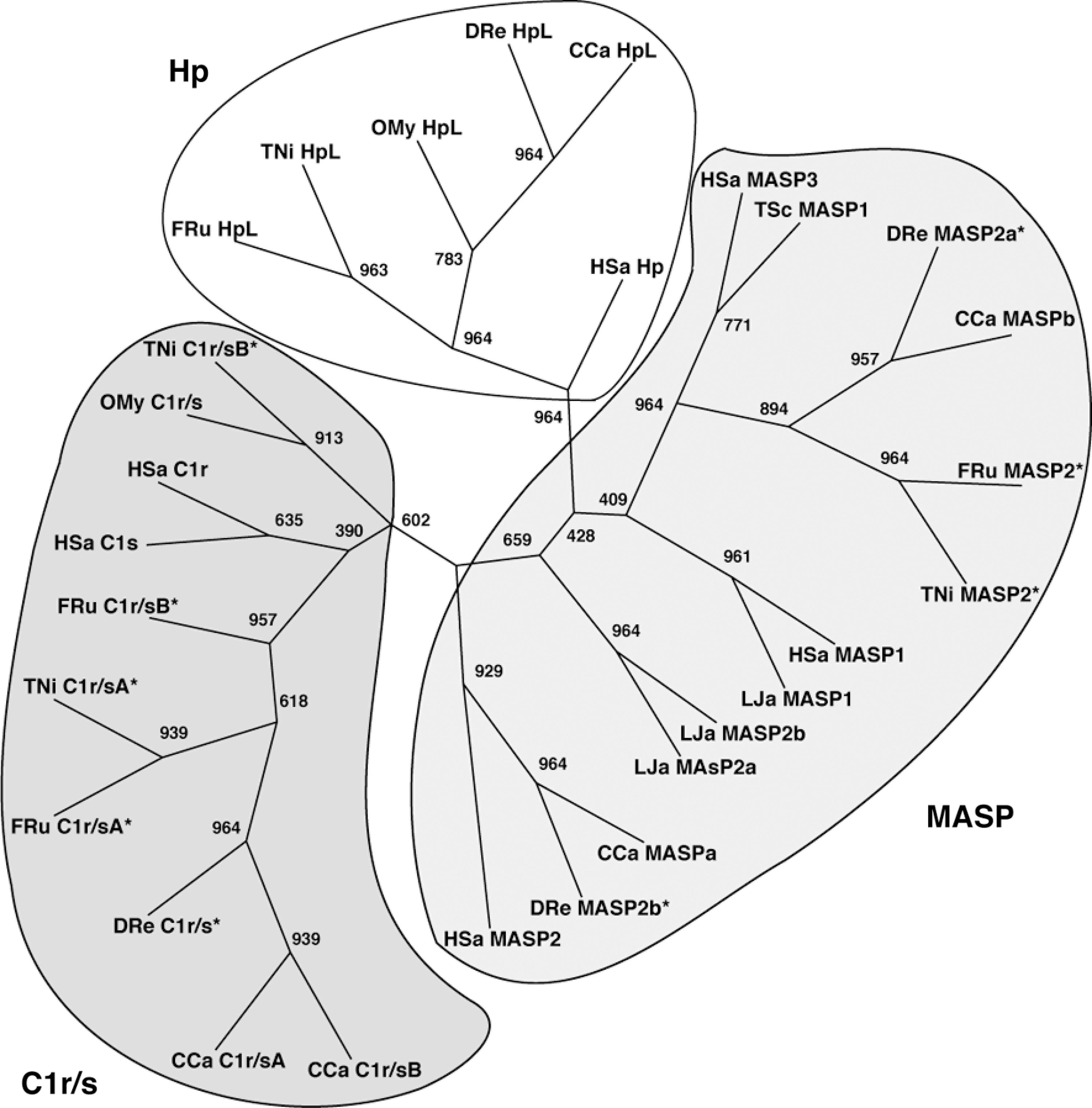
Cartilaginous fish seem to represent an evolutionary lineage preceding that of bony fish. Our analysis of EST libraries of two fish belonging to this group, dogfish shark (Squalus acanthias) and little skate (Leucoraja erinacea), did not reveal an Hp gene (unpublished data). The proteins most closely related to Hp that we found were mannan-binding lectin–associated serine proteases (MASPs). These proteins are part of the innate immune system and are found in both invertebrates and vertebrates. MASPs are proteolytically activated on binding of an associated lectin to the surface of a microorganism. Similarly, the complement components, C1r and C1s, in complex with C1q, are proteolytically activated on binding of C1q to a target (23). Clear structural similarities exist between Hp, MASP-2, C1r, and C1s, as shown by Tosi et al. (81).
We also analyzed the genomes of five invertebrates: Saccharomyces cerevisiae (baker's yeast), Coenorhabditis elegans (a nematode), Drosophila melanogaster (fruit fly), Ciona intestinalis (sea squirt), and Brachiostoma floridae (lancelet), as well as the primitive vertebrate Petromyzon marinus (lamprey). We were unable to find a gene encoding a protein similar to mammalian Hp in any of these species (unpublished data); the closest relatives were orthologues of vertebrate MASP in P. marinus.
Origin and evolution of the haptoglobin gene
The difference in domain structure between fish and mammalian Hp raises the question of whether the two proteins have a common ancestor. Figure 5 shows a phylogenetic tree for the SP domains of human and fish Hp, as well as MASP proteins and C1r/s. Clearly, human and fish Hp are on a branch distinct from those containing the complement proteins, indicating a common origin. Ancestries can also be studied through analysis of the chromosomal regions comprising the respective genes. As shown in Fig. 6, both human and fish Hp genes are located next to DHODH (a single-copy gene), suggesting a common origin. This figure also shows that the human Hp gene is flanked by DHODH and DHX38, and that in X. tropicalis and chicken, DHODH and DHX38 are located close together. These chromosomal arrangements indicate that the whole Hp gene was deleted in an ancestor of X. tropicalis and chicken. Given the fact that amphibians emerged before the divergence of reptiles/birds and mammals, this deletion must have occurred independently in the amphibian and avian lineages.

Based on the observations described above, the evolution of fish and mammalian Hp could be explained by the sequence of events shown in Fig. 7. It is likely that C1r, C1s, and MASP in both mammals and fish have a common ancestor with the same modular composition as these proteins have today: two CUB domains, one epidermal growth factor (EGF)-like domain, two CCP domains, and one SP domain (23). Through partial duplication of the gene for this ancestor, the Hp prototype (prtHp) appeared (step I). This protein had a domain organization similar to that of mammalian Hp (consisting of both a CPP and an SP domain) but was still an active protease. Subsequently, the essential serine and histidine residues in the active site of its SP domain were substituted with alanine and either lysine or arginine (step II), which resulted in a proteolytically inactive protein (ancHp). All these events occurred in a common ancestor of bony fish and mammals. In the fish lineage, the CCP domain was then deleted (step III), giving rise to the present fish Hp.
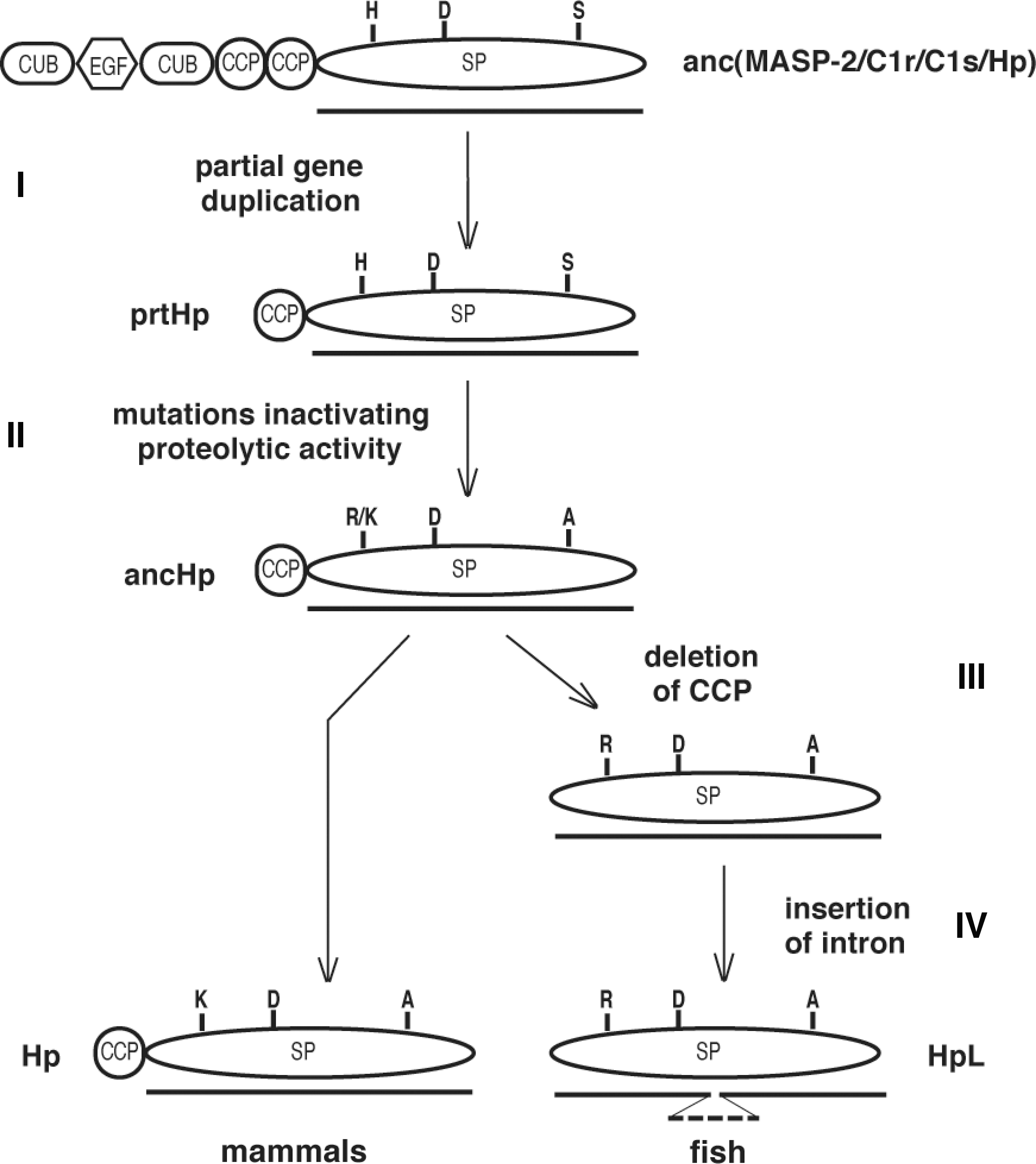
Intracellular Cleavage of Haptoglobin
As described earlier, mammalian Hp is cleaved in the endoplasmic reticulum by a complement-related protein (C1r-LP), whereas fish Hp is cleaved in the Golgi complex by proprotein convertases. How can this difference in processing be explained in evolutionary terms? As shown in Fig. 7, Hp seems to have evolved by partial duplication of a gene encoding a complement protease similar to today's MASP2/C1r/C1s. This type of protein contains an SP domain connected to a CCP domain, which is also the case for mammalian Hp. MASP, as well as C1r/C1s, is activated extracellularly by a proteolytic cleavage between the CCP and the protease domains, either autocatalytically or by another protease of the complement system (76). The fact that human Hp is cleaved in vivo by a protein derived from C1r (91) and in vitro by C1r (unpublished data) strongly supports the idea that the ancestral Hp was also cleaved in this fashion. It is possible that a need for constantly active Hp in the blood, in the absence of activated complement components, appeared later. The appearance of already active C1r-LP in the endoplasmic reticulum might have fulfilled this need in an ancestral mammal; we have found the gene for C1r-LP in opossum (a marsupial) but not in platypus, probably the most primitive living mammal (unpublished data). Similarly, an alteration of the amino acid sequence at the cleavage site, making the protein a substrate for already existing proprotein convertases, could have served the same purpose in an ancestral bony fish.
Cleavage of Hp has been reported to increase the affinity for Hb (30), but the physiological relevance of this observation remains to be determined. However, Hp has been shown to have biologic activities other than Hb binding, such as suppressing lipopolysaccharide-induced proliferation of T cells and B cells (3, 5) and stimulating angiogenesis (14). It is not known whether proteolytic processing is necessary for any of these activities.
Cell-Surface Receptors
After free Hb binds to Hp, the complex is rapidly taken up by a receptor, CD163, residing on the surface of monocytes/macrophages (37). The receptor and its ligand are then endocytosed and degraded (59). CD163 only binds Hp in complex with Hb, whereas Hb can bind alone to CD163, but with low affinity (73). CD163 belongs to group B of the family of scavenger-receptor cysteine-rich (SRCR) proteins. Members of this family consist of one or several SRCR domains, which are ancient conserved modules of 100 to 110 amino acid residues (69, 71). Human CD163 comprises nine SRCR domains, one of which has been shown to account for the binding of Hp-Hb complexes (44). PIT54, the Hb-binding protein of chicken plasma, also is a member of this family and consists of four SRCR domains. The sequence identity between the different domains of PIT54 and the chicken orthologue of CD163 is 47–51% (unpublished result). The corresponding value for PIT54 and another member of the SRCR family, an orthologue of human deleted in malignant brain tumors-1 protein (DMBT-1) (71), is 63–67% (unpublished observation). These findings suggest that other members of the SRCR family might also have the capacity to bind Hb (72). Whether PIT54, like CD163, actually mediates plasma clearance of Hb remains to be tested. For a comprehensive review of hemoglobin-scavenger receptors, see the contribution by Nielsen et al. in this issue.
Heme-Binding Proteins
On release of Hb from erythrocytes, part of the protein is oxidized to ferrihemoglobin, which dissociates into ferriheme and globin (12). Initially, most of the free heme binds to lipoprotein, but this fraction is gradually transferred to hemopexin and serum albumin (4). Heme-binding proteins can be detected by using the same electrophoretic method as that described earlier for Hb, or isolated by binding to immobilized heme.
Hemopexin
Of all plasma proteins, hemopexin has the highest affinity for heme (4, 80). Hemopexin is an acute-phase protein synthesized mainly by hepatocytes and has been detected in the plasma of all mammals analyzed to date. An orthologous heme-binding protein has been found in different fish species, such as medaka and trout (31). With BLAST analysis, we found hemopexin in the cartilaginous fish (skate, dogfish, and shark) but not in the more primitive jawless fish (lamprey and lancelet) (unpublished data). The hemopexin-heme complex is taken up by a multiligand receptor (low-density lipoprotein receptor–related protein 1 or CD91), which is present on many cell types, such as macrophages and hepatocytes (34). For a detailed review of hemopexin, see the contribution of Tolosano et al. in this issue.
Serum albumin
When the extracellular Hb level exceeds the binding capacity of Hp and hemopexin, heme will bind to serum albumin, forming methemalbumin. With electrophoresis, methemalbumin was shown to form in amphibians, reptiles, and anthropoid primates but not in fish, birds, and nonprimate mammals (6, 42). It has been suggested that the occurrence of methemalbumin in primates may be of importance during hemolytic conditions occurring in prenatal life, when Hp and hemopexin levels are either low or absent (6). Heme bound to albumin has been shown to be taken up by hepatocytes (78), possibly through specific receptors (22).
α1-Microglobulin
α1-Microglobulin is a 26-kDa protein synthesized mainly in the liver. After secretion, part of α1-microglobulin becomes covalently linked to other proteins, such as IgA, and is partly filtered through the kidneys (8). α1-Microglobulin belongs to the lipocalin family, whose members contain an eight-stranded β-barrel surrounding a pocket that can bind hydrophobic ligands. It was recently shown that a truncated form of α1-microglobulin binds and degrades heme (1) and that the protein can act as an antioxidant (2). α1-Microglobulin orthologues with the capacity to bind heme have been found in birds, amphibians, and bony fish (41).
Conclusions
In this review, we describe how systems for scavenging Hb and free heme seem to have evolved with the appearance of cartilaginous and bony fish, respectively. An increase in the amount of Hb in the blood and a higher oxidizing capacity of Hb (9, 20) might have been factors behind this development. It should also be noted that genome duplications have apparently occurred on either side of the divergence of jawless fish, greatly facilitating the evolution of proteins with new functions (17). The occurrence of Hp in animals belonging to different phylogenetic lineages is shown in Fig. 8. Our analysis indicates that Hp evolved from the ancestor of the complement components MASP2/C1r/C1s by the removal of four or five of the original six domains. It is possible that the need for an Hb-binding protein was the driving force behind the evolution of Hp. However, Hp also has antiinflammatory functions (33), and it is conceivable that these evolved before Hb binding. This hypothesis is supported by the existence of two other Hb-binding proteins: CD163 in mammals and PIT54 in birds. The ancestor of these proteins was probably a scavenger receptor, which might also have had the capacity to bind Hb. Further studies of the Hb-clearance mechanism in animals representing different evolutionary stages should clarify this issue.

It is remarkable that some animal groups lack Hp, a phenomenon suggesting that a disadvantage is associated with its expression. This is the case for at least one type of amphibian and one type of lizard; animals with a relatively low metabolic rate and presumably a low need for Hb scavenging. At least one class of birds, including chicken, also lacks Hp, but here a completely unrelated protein, PIT54, seems to have replaced its function. Although all mammalian species seem to have Hp in their blood, a small number of human individuals have genetically based anhaptoglobinemia. Evidence suggests that this condition provides partial resistance to malaria (16). The appearance of multimeric Hp molecules among humans and ruminants is another evolutionary change in mammals. Here again, the change seems to provide partial protection against certain pathogens, at least in humans (85). Unfortunately, diabetic individuals with the multimeric form of Hp are at a much higher risk for cardiovascular complications; see the contribution of Levy et al. in this issue.
Footnotes
Acknowledgments
This work was supported by the Swedish Research Council and the O.E. and Edla Johansson Scientific Foundation. We thank B. Tomkinson for comments on the manuscript.