
Abstract
Hydrogen sulfide (H2S) is an important signaling molecule produced from L-cysteine by cystathionine β-synthetase (CBS) or cystathionine γ-lyase (CSE). Here we examined the role of H2S in the oxygen-sensing function of the carotid body chemoreceptors, where the large conductance Ca2+-activated potassium channel (BKCa) plays a key role. In the isolated mouse carotid body/sinus nerve preparations, the H2S donor, NaHS, excited the chemoreceptor afferent nerves in a concentration-dependent manner that was reversed by carbon monoxide donor. The NaHS-evoked excitation was abolished by removing extracellular Ca2+, or using Cd2+, pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid and hexomethonium, suggesting that H2S evokes release of ATP/ACh from type I glomus cells of the carotid body. The chemoreceptor afferent activation by hypoxia was decreased remarkably using CBS inhibitors, amino oxyacetic acid (AOAA) and hydroxylamine, but not CSE inhibitors, propargylglycine and β-cyano-L-alanine, despite expression of both enzymes in type I glomus cells. In these cells, the BKCa currents were inhibited by hypoxia and such inhibition was mimicked by NaHS and diminished by AOAA. Finally, mice hyperventilated in response to hypoxia, which was prevented by CBS inhibitors. These data suggest that H2S plays a crucial role in mediating the response of carotid body chemoreceptors to hypoxia via modulating the BKCa channels. Antioxid. Redox Signal. 12, 1179–1189.
Introduction
Hypoxia is known to decrease the potassium conductance in type I glomus cells of the carotid body (8, 14, 19) and other oxygen-sensitive cells (13, 23, 27, 38). In the carotid body, it has been proposed that heme oxygenase-2 (HO-2) functions as an oxygen sensor through primary production of carbon monoxide (CO), which in turn activates the BKCa channel (12, 41). However, a recent study has shown that the chemoreceptor response to hypoxia in HO-2 deficient mice remains intact (26), suggesting other signaling molecule(s) exist and mediate the chemoreceptors response to hypoxia via modulation of the BKCa channels.
Hydrogen sulfide (H2S) is present in many mammalian tissues and endogenously synthesized from L-cysteine by cystathionine-β-synthetase (CBS) or cystathionine-γ-lyase (CSE) (11, 15, 33). Previous studies have demonstrated that H2S is an important signaling molecule exerting a wide spectrum of biological effects in the nervous, cardiovascular, and immune systems (21, 29, 33, 36). Inhalation of large quantities of H2S results in cessation of ventilation, disturbance of oxygen homeostasis, and neurological dysfunctions (2, 9). However, it is unknown how H2S regulates the chemoreceptor function and particularly what role it plays in the physiology of oxygen homeostasis. In the present study, we investigated the effects of H2S on the responses of the carotid body chemoreceptor to hypoxia. We found that the chemoreceptors were activated by exogenous H2S, as were in response to hypoxia. Blocking endogenous H2S production remarkably attenuated the chemoreceptor responses to hypoxia. Furthermore, exogenous H2S resembled the strong inhibition by hypoxia of the BKCa channel currents in type I glomus cells, whereas lowering endogenous H2S prevented the inhibition of BKCa channel currents by hypoxia. Finally, mice exhibited hyperventilation under hypoxia, and this compensatory mechanism was abolished by inhibiting endogenous H2S generation. These results taken together suggest that H2S plays a crucial role in oxygen sensing of the carotid body via modulating the BKCa channel activity.
Materials and Methods
Adult Kunming mice (25–30 g) of either sex were used. All procedures were performed in accordance with the institutional guidelines of Shanghai Jiaotong University on the use of experimental animals.
Isolated mouse carotid body and chemoreceptor afferent recording
Isolated carotid body/sinus nerves were prepared, and the effects of H2S on the function of the chemoreceptors were examined as described previously (31). Briefly, mice were deeply anesthetized by intraperitoneal injection of pentobarbital (100 mg/kg) and killed by cervical dislocation. The carotid bifurcation region containing carotid body and attached sinus nerve was carefully dissected, and placed into a recording chamber (3 ml) perfused with oxygenated (21% O2 + 5% CO2) Krebs solution at a rate of 15 ml/min and kept at 34°C. The sinus nerve was carefully cleared by removing connective tissues, and recorded using a suction electrode. Nerve activity was amplified (20,000X) and filtered (200–3,000 Hz), and recordings were stored by a personal computer using a Spike 2 data acquisition and analysis program (Cambridge Electronic Design, Cambridge, UK). The sensitivity of chemoreceptor afferents to hypoxia was examined by switching the superfusate to Krebs solution bubbled with hypoxic gas mixture (5% O2 + 5% CO2 + 90% N2) for 3 min at an interval of 15 min. The effects of exogenous H2S were tested by switching to Krebs solution containing NaHS. To examine the role of endogenous H2S in oxygen sensitivity, the tissues were challenged with hypoxic solution with or without CBS or CSE inhibitors.
Type I glomus cell culture and whole-cell recording
The carotid bodies were cut into pieces and incubated in 1 ml phosphate saline buffer (PBS) containing 0.05% collagenase type II (Sigma, Shanghai, China) and 0.025% trypsin at 37°C for 20 min. After extensive wash with PBS, cells were dispersed by gentle agitation with a Pasteur tube in F-12 culture medium. Dissociated cells were plated onto glass coverslips and cultured in F-12 medium (supplemented with 5% fetal calf serum, 100 U/ml penicillin G, 0.1 mg/ml streptomycin, and 84 U/ml insulin) at 37°C in an incubator circulated with air and 5% CO2, and used within 24 h.
Whole-cell recordings were made using an EPC10 amplifier (HEKA, Lambrecht/Ptalz, Germany), using pipettes with a resistance of 3–5 MΩ. The holding potential was −60 mV. Extracellular solution contained (in mM) 141 NaCl, 10 HEPES, 4.7 KCl, 1.2 MgCl2, 1.8 CaCl2, 10 glucose, pH 7.4. Pipette solution contained (in mM) 125 KCl, 4 MgCl2, 10 HEPES, 5 MgATP, 5 Na3GTP, and 1.1 EGTA. A microperfusion tube was positioned about 25 μm away from the patched cells to allow rapid administration of normoxic (95% O2 + 5% CO2) and hypoxic solutions (95% N2 + 5% CO2) or drugs. Type I glomus cells are known to express multiple potassium channels, including voltage-gated potassium channels (KV1-3) (20, 28), twin pore acid sensitive potassium channels (3), and BKCa (30). We used a protocol consisting of a pre-pulse (0 mV, 100 ms) followed by test pulses (400 ms) from −80 to 80 mV with 10 or 20 mV increments, and analyzed cells that exhibited minimal outward currents during the pre-pulse.
CBS and CSE immunostaining
Immunohistochemistry was performed on formalin-fixed, paraffin-embedded 5 μm sections of the carotid bifurcation region. The tissues were deparaffinized, and then dehydrated with graded alcohol and xylene. After extensive wash with PBS, endogenous peroxidase was blocked using 0.3% (vol/vol) hydrogen peroxide for 10 min, followed by washing with PBS. The sections were treated with 0.3% BSA for 30 min to block nonspecific binding, and then incubated at 4°C overnight with monoclonal mouse anti-CBS (a dilution of 1:200; ABNOVA, Taipei; Taiwan) or anti-CSE antibody (1:400; ABNOVA). Control sections were without the primary antibodies. After wash in PBS, the sections were incubated with biotinylated rabbit anti-mouse IgG antibody (1:200; Dako, Glastrup, Denmark) for 30 min at 37°C, followed by extensive washing in PBS and incubation with streptavidin/HRP (1:300; Dako) at 37°C for 30 min. Reaction product was visualized with 3,3-diaminobenzidine for 5 min. After brief exposure to hematoxylin stain, the sections were rinsed with water and then dehydrated by sequential immersion in gradient ethanol and xylene. Images were obtained under a light microscope equipped with a digital camera.
For confocal microscopy, sections were washed in PBS with 0.2% Triton and incubated with 10% (vol/vol) goat serum in PBS for 30 min. The sections were then incubated at 4°C with a mouse anti-CBS or anti-CSE antibody. After several washes with PBS, the sections were incubated with tetramethyl rhodamine isothiocyanate-conjugated goat anti-mouse IgG (1:100 in PBS; Sigma) and co-stained at 37°C for 2 h with fluorescein isothiocyanate-conjugated anti-PNA antibody (1:200; Sigma). After being thoroughly rinsed in PBS, the sections were viewed under a Zeiss (Jena, Germany) Axioplan 2 microscope or inverted laser-scanning confocal microscope (model 510 CLSM).
Whole body plethysmography
Mice were placed individually in a Plexiglas recording chamber (500 ml) that was flushed continuously with a mixture of 79% nitrogen and 21% oxygen at a rate of 300 ml/min. The pressures within the chamber were monitored via a pressure transducer connected to a bridge amplifier and the signal was filtered, recorded, and analyzed offline using Spike 2 software (CED, Cambridge, U.K.). The animals were allowed to acclimatize to the chamber environment in normoxia for 30 min before measurement of the baseline ventilation. The chamber was then flushed with a hypoxic gas mixture (5% O2 + 95% N2) at the same rate (300 ml/min) for 2 min. This was repeated after 15 min. The animals were then injected i.p. with either saline or test drugs, and challenged again with hypoxic gas mixture 15 min later. The respiratory rate (Fr) and tidal volume (Vt) were determined by the pressure trace, and minute ventilation (ml/min) was derived from Fr × Vt.
Statistics
The data are, where appropriate, presented as mean ± SEM. Statistical analysis was performed using paired Student's t-test or Wilcoxon signed rank test with results being considered significant for p < 0.05.
Results
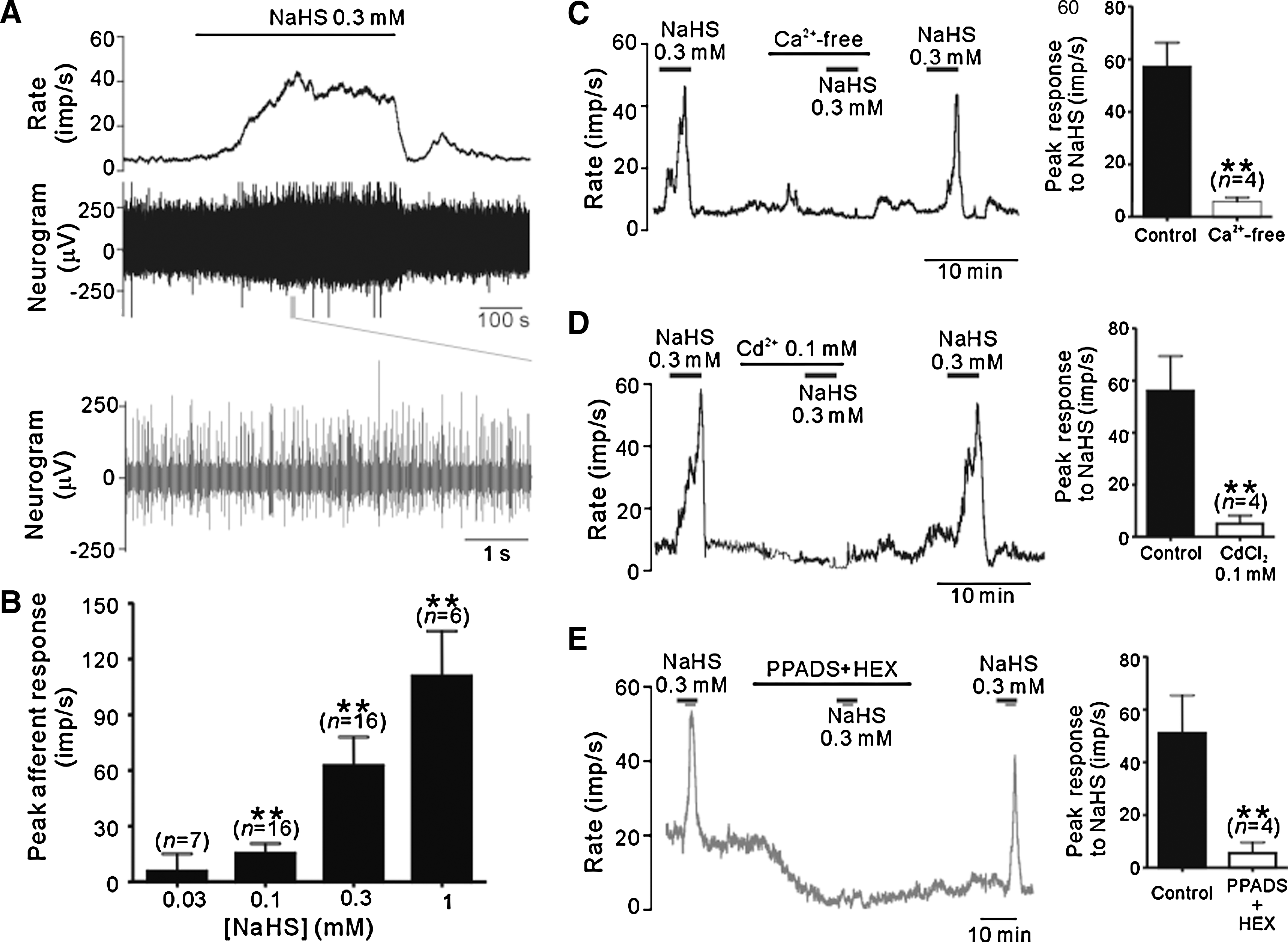
Effects of H2S on spontaneous afferent activity
Figure 1 shows the effects of exogenous H2S on the chemoreceptors in isolated carotid body/sinus nerve preparations. NaHS, a H2S donor, induced remarkable spontaneous afferent activity in a concentration-dependent manner; the peak afferent activity was increased by 15.3 ± 5.4 (n = 16, p < 0.05), 62.5 ± 15.3 (n = 16; p < 0.01) and 110.7 ± 24.2 impulses per second (imp/s) (n = 6; p < 0.01), following application of 0.1, 0.3, and 1 mM NaHS, respectively, while there was no significant effect by 0.03 mM NaHS (n = 7) (Fig. 1B). The NaHS-evoked afferent activity was almost completely abolished in the presence of Ca2+ free extracellular solution (Fig. 1C) or 100 μM Cd2+ (Fig. 1D), a nonselective Ca2+ channel blocker. Adenosine-5’-triphosphate (ATP) and acetylcholine (ACh) are the two major transmitters released from type I glomus cells (22). Consistently, the afferent excitation by NaHS was prevented by co-administrating pyridoxalphosphate-6-azophenyl-2',4'-disulfonic acid (PPADS, 30 μM), a P2X receptor antagonist, and hexomethonium (HEX, 30 μM), a nicotinic receptor antagonist (Fig. 1E). These results taken together suggest that NaHS primarily facilitates release of ATP/ACh from type I glomus cells that subsequently excite the afferent terminals.
Effects of H2S on hypoxia-evoked afferent activity
We next examined the role of H2S in oxygen sensing in the carotid body. We first compared the afferent response to hypoxia in the presence and absence of NaHS (Fig. 2A). In response to hypoxia in the absence of NaHS, the afferent activity started to increase after a latency of 132.8 ± 6.9 s (n = 14), and reached a peak value of 123.9 ± 22.3 imp/s (n = 14). While there was no significant effect on the peak afferent response to hypoxia, NaHS considerably shortened the response latency; the mean latency was 84.2 ± 8.5 s (n = 6; p < 0.05, Wilcoxon signed rank test) and 49.5 ± 3.4 s (n = 8, p < 0.01) in the presence of 0.03 and 0.1 mM NaHS, respectively, suggesting that H2S and hypoxia have additive or synergistic actions.

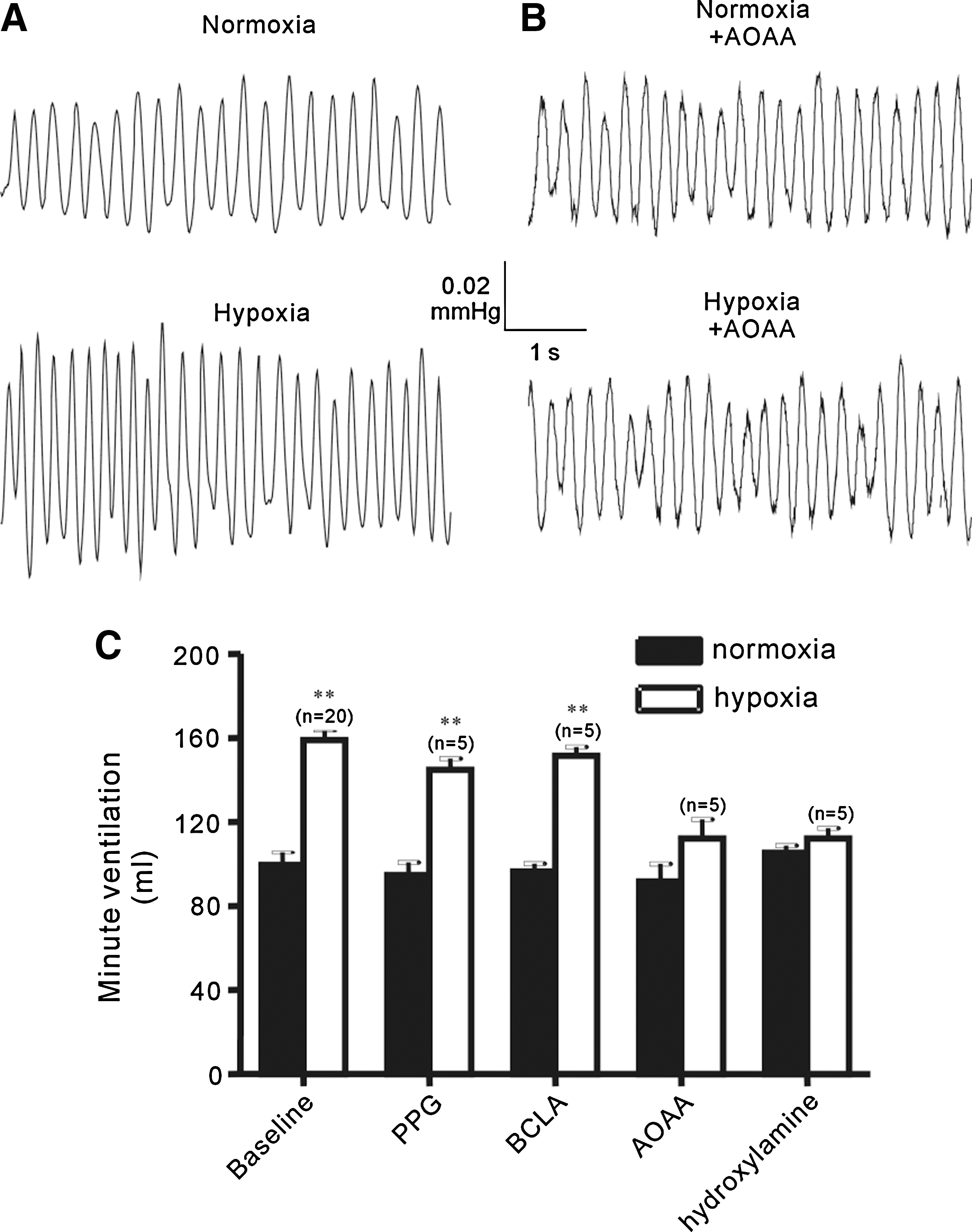
We then used CBS and CSE inhibitors to prevent generation of H2S to explore the role of endogenous H2S in oxygen sensing in the carotid body. Pretreatment with the CBS inhibitors, amino oxyacetic acid (AOAA, 0.3 mM) or hydroxylamine (0.3 mM) (1, 47), remarkably decreased the afferent responses to hypoxia (Fig. 2B). Pretreatment with CSE inhibitors alone (D,L-propargylglycine, PPG, 0.1–1 mM, or β-cyano-L-alanine, BCLA, 0.1–1 mM) (1, 47) had no significant effect on the hypoxia-evoked afferent responses (Fig. 2C). However, PPG appeared to increase the efficacy of CBS inhibitors (AOAA and hydroxylamine). Thus, although 100 μM AOAA only resulted in a small decrease in hypoxic response and PPG at concentrations of up to 1 mM had no effect, combined use of 100 μM AOAA and 100 μM PPG led to marked reduction in the hypoxia-evoked afferent activity (Fig. 2D). These data suggest that endogenous H2S is important in mediating the chemoreceptor responses to hypoxia.
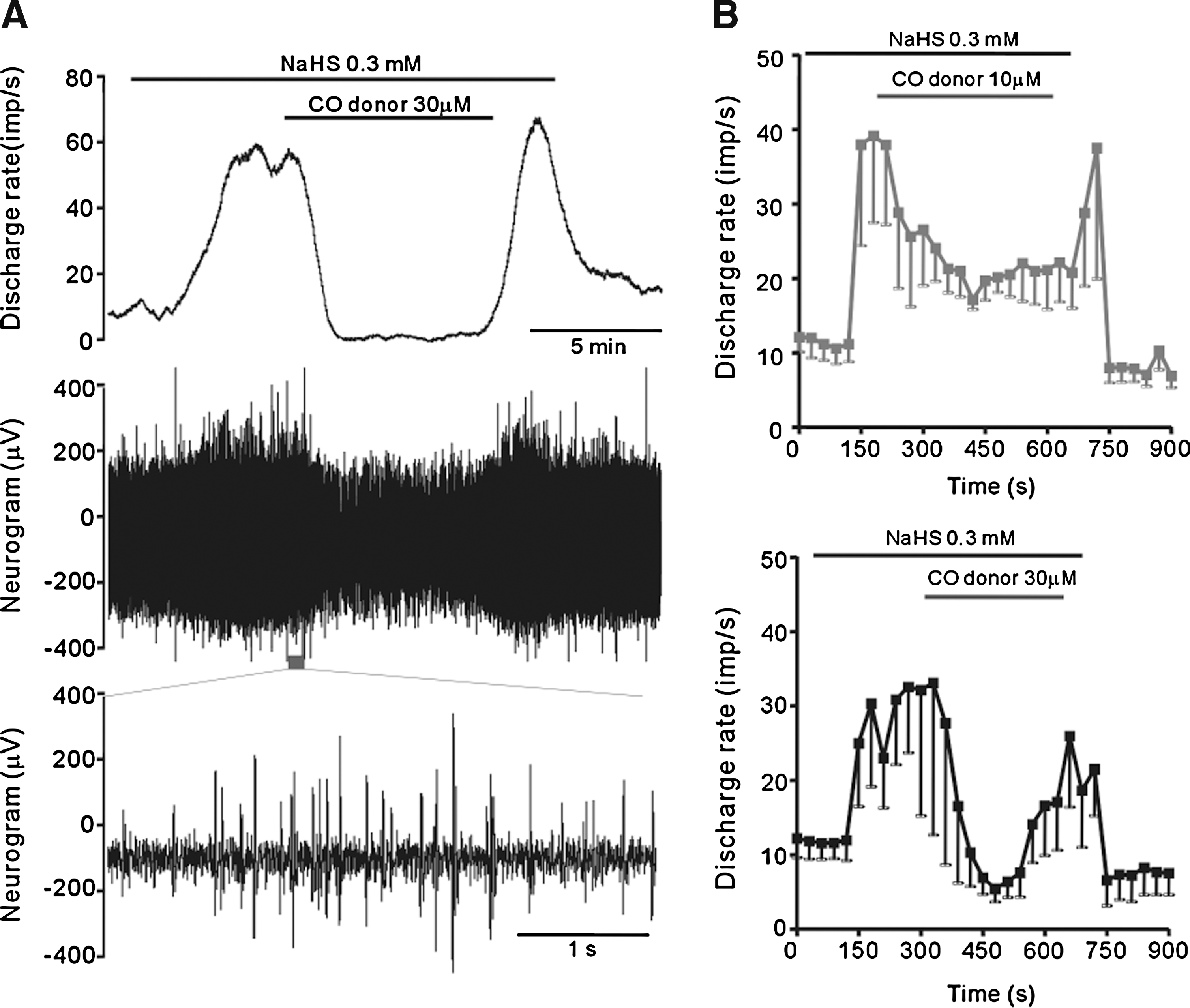
Effects of CO donor on H2S-evoked afferent responses
Previous studies by Kemp and colleagues implicated CO as a major player in oxygen-sensing via interacting with the BKCa channels in type I glomus cell (41). We therefore carried out experiments to examine the possible effect of CO donor on NaHS-evoked afferent responses. When the CO donor, [Ru(CO3)Cl2]2 (41), was applied at 30 μM, there was no significant change in the baseline spontaneous afferent activity (data not shown). However, the CO donor consistently reversed the stimulatory effect on the carotid body stimulated with NaHS (0.3 mM) (Fig. 3), and at 30 μM was able to bring the nerve activity back to the baseline. These results led us to speculate that H2S might act via interacting with the CO-modulated BKCa channels in type I glomus cells.
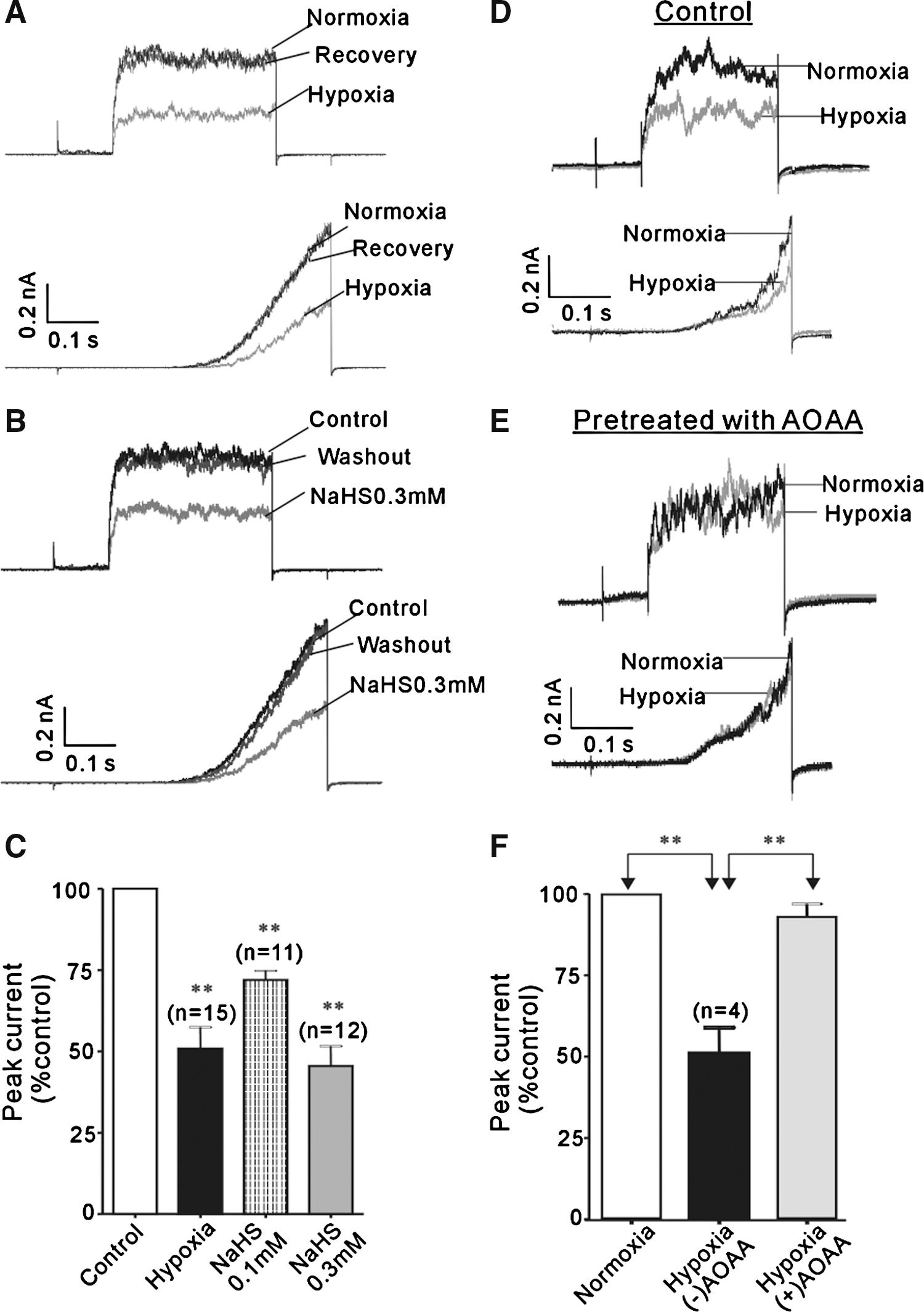
Effects of H2S on BKCa channel currents in type I glomus cells
We made whole-cell recordings on type I glomus cells to examine whether H2S acts on the BKCa channels. Type I glomus cells were round cells with a diameter of < 10 μm. The BKCa channel currents in these cells were almost completely abolished by 3 μM paxiline, a selective BKCa channel blocker (data not shown). Hypoxia strongly inhibited the BKCa channel currents (49.2 ± 6.6%; n = 15) in a voltage-independent manner (Fig. 4A), as previously described (41). NaHS mimicked the inhibition by hypoxia of the BKCa channels (Fig. 4B); the mean currents were reduced by 28.1 ± 2.8% (n = 11) and 54.5 ± 6.1% (n = 12) in the presence of 0.1 and 0.3 mM NaHS, respectively (Fig. 4C). Noticeably, the inhibition by hypoxia and NaHS were both fully reversible. We also tested whether endogenous H2S plays a role in the sensitivity of BKCa channels to hypoxia by using AOAA (0.3 mM). In the presence of AOAA, hypoxia had negligible inhibition (6.9 ± 4.0%; n = 4) (Fig. 4D–F). Consistent with the observation that the CO donor could reverse the excitatory effect of H2S on the chemoreceptor afferent activity, the BKCa current inhibition by NaHS was nicely reversed by the CO donor (Fig. 5). These data taken together suggest that H2S mediates the inhibitory action of hypoxia on the BKCa channels.

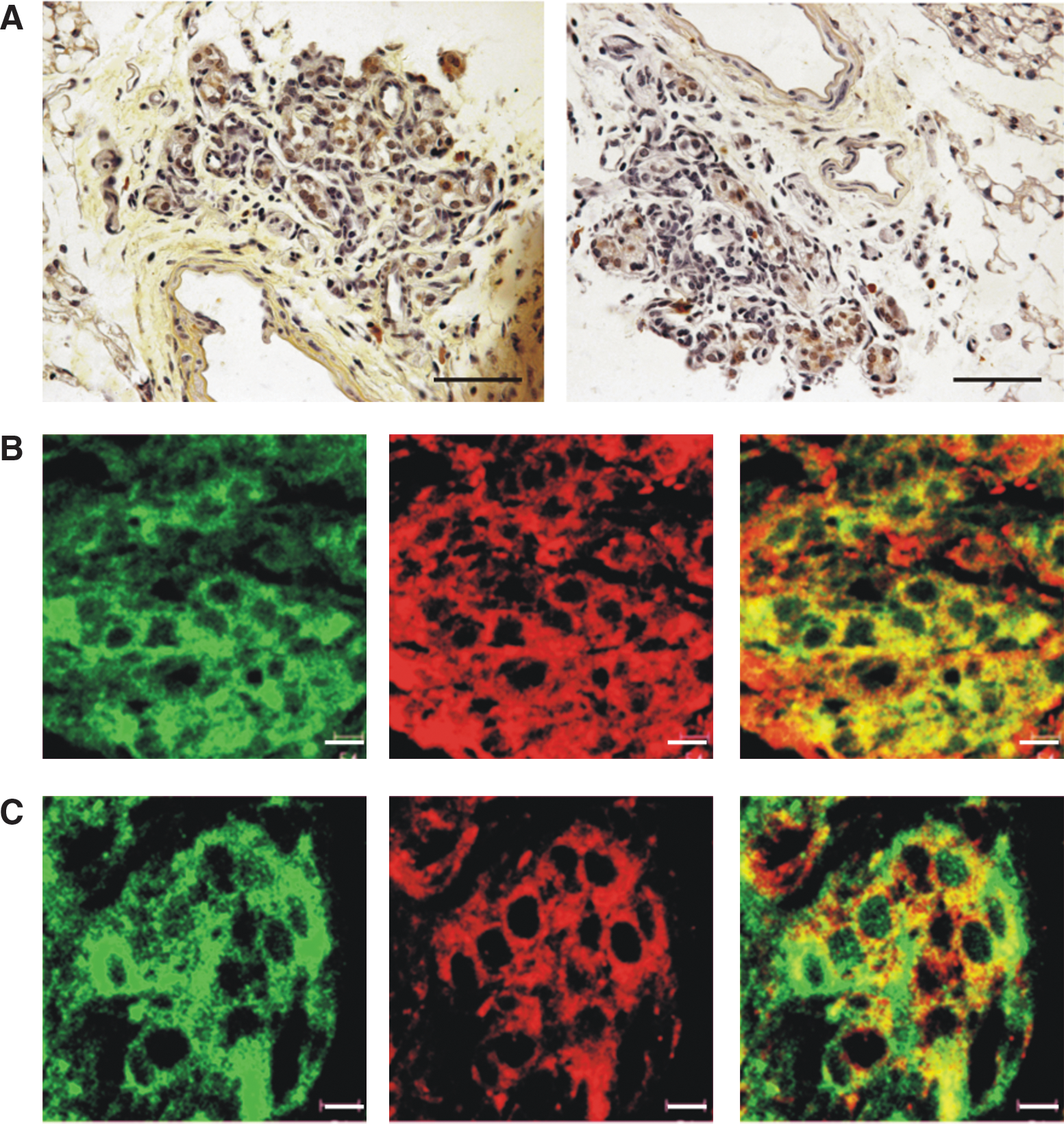
Expression and localization of H2S-producing enzymes in carotid body
We carried out immunostaining to examine whether CBS and CSE enzymes are expressed in the carotid body. CBS and CSE immunoreactivity was present within lobules of the chemoreceptive organ of carotid body (Fig. 6A). Further double labeling using an antibody recognizing the peanut agglutinin receptor (PNA), a protein marker for type I glomus cells, show that virtually all PNA-positive glomus cells within lobules were co-stained strongly with CBS and CSE (Fig. 6B and C). The staining pattern indicates that CBS and CSE immunoreactivity is localized closely to the cytoplasmic side of the membrane of the chemoreceptive cells.
Effects of H2S-producing inhibitors on the hyperventilation induced by hypoxia
Finally, we used the CBS and CSE inhibitors to study the role of endogenous H2S in the hyperventilatory responses to hypoxia in vivo by recording minute ventilation of mice using single chamber plethysmography (Fig. 7). Under control conditions, the average minute ventilation was 125 ± 12 ml in normoxia and increased to 199 ± 14 ml (n = 20, p < 0.01) in response to hypoxia (Fig. 7A and C). Hypoxia-induced hyperventilation was abolished in mice after i.p. injection with AOAA (300 μmol/kg); the minute ventilation in hypoxia (140 ± 11 ml) was not significantly different from that in normoxia (115 ± 11, n = 5, p > 0.05) (Fig. 7B and C). Similar results were obtained with hydroxylamine (300 μmol/kg) (Fig.7C). Consistent with lack of significant effect on hypoxia-induced afferent activity (Fig. 2C), mice injected with PPG or BCLA (both 300 μmol/kg) exhibited normal hyperventilatory responses to hypoxia (Fig. 7C). Of notice, there was no change in the minute ventilation in normoxia by any of these inhibitors (Fig. 7C).
Discussion
The major findings of the present study are two fold. First, H2S excites the carotid body chemoreceptors and inhibits the BKCa channels in type I glomus cells. Second, H2S is essential for hypoxia-evoked excitation of the carotid body chemoreceptor in vitro and hyperventilation in vivo. Therefore, H2S plays a crucial role in oxygen-sensing mechanism of the carotid body by modulating the BKCa channels.
Numerous lines of evidence suggest that H2S is an important gaseous signaling molecule, as the third gasotransmitter after nitric oxide (NO) and CO, regulating a wide spectrum of physiological functions (see Introduction). In the present study, we showed that H2S production was essential in oxygen sensing of the carotid body. In the isolated carotid body/sinus nerve preparations, exogenous H2S activated the afferent nerves in a concentration-dependent manner (Fig. 1A), and at lower concentrations facilitated the response of afferent nerve to hypoxia (Fig. 2A). Similarly to hypoxia, H2S appeared to primarily act on type I glomus cells rather than afferent terminals, because the effects were abolished by application of Ca2+-free extracellular solution or cadmium to prevent transmitter release from glomus cells (Fig.1B and C). The chemoreceptor excitation by H2S was also abrogated by blocking synaptic transmission within the chemoreceptor organ by co-application of PPADS and hexomethonium (Fig. 1D), consistent with the fact that ATP and ACh are the two major transmitters in type I glomus cells (22). We further showed a critical role of endogenous H2S in oxygen-sensitivity of the carotid body by inhibiting H2S-producing enzymes (Fig. 2B). CBS and CSE are the two enzymes that catalyze generation of endogenous H2S in many mammalian tissues; CBS is predominantly expressed in the nervous systems and CSE is mainly found in the vascular smooth muscles (6, 49). We performed immunohistochemistry to show expression of both CBS and CSE in type I glomus cells, located closely to the cytoplasmic side of the cell membrane. In the isolated carotid body/sinus nerve preparations, hypoxia-induced afferent discharge was effectively abolished by the CBS inhibitors, AOAA and hydroxylamine, but not by the CSE inhibitors, PPG and BCLA (Fig. 2B and C). PPG had no significant effect but seemed to enhance the efficacy of CBS inhibitors in attenuating the hypoxia-evoked afferent discharge (Fig. 2D). Moreover, injection of CBS but not CSE inhibitors abolished the hyperventilatory responses to hypoxia in mice (Fig. 7). These data provide strong evidence indicating that endogenous H2S is necessary for the responses of the carotid body chemoreceptors to hypoxia. These results also suggest that in our preparations CBS is the predominant H2S generating enzyme or alternatively, CBS is closely associated with the BKCa channels.
The mechanisms underlying the majority of H2S-induced effects are not fully understood, with a few exceptions. For example, relaxation of smooth muscles and inhibition of insulin secretion from pancreatic β-cells primarily result from direct activation of the KATP channels (10, 46, 48, 49). Apparently, this cannot explain the excitatory effects of H2S on the chemoreceptors. Since the afferent excitation evoked by H2S was nicely reversed by CO donor, it is tempting to speculate that H2S might target the CO-modulated BKCa channels. Whole cell current recordings from acutely dissociated type I glomus cells clearly showed exogenous H2S resulted in strong inhibition of the BKCa channels, resembling the action of hypoxia (Fig. 4A and B) and this effect was reversed by the CO donor (Fig. 5). More importantly, inhibition of the BKCa channels by hypoxia was remarkably attenuated by inhibiting production of endogenous H2S (Fig. 4C). Therefore, our results suggest that H2S production is required for functional inhibition of the BKCa channels by hypoxia.
It has been estimated that approximately one-third of NaHS forms H2S in physiological solutions, and therefore the concentrations of exogenous H2S applied in this investigation would be in the range of 10–100 μM, which is comparable to the level previously reported in blood (10–100 μM) (49) and brain (150 μM) (37). However, a very recent study has showed that circulating free sulfide level is considerably low as it is readily oxidized to inactive metabolites (i.e., sulfite and/or sulfate) in the presence of O2 (40). On the other hand, the same study has clearly demonstrated that hypoxia is sufficient to increase the local H2S concentrations. The same group also reported that hypoxia and H2S produce temporally and quantitatively identical responses in blood vessels and that inhibition of H2S synthesis inhibited the hypoxic response, suggesting that H2S may serve as an O2 sensor/transducer in vascular responses to hypoxia (24). More recently, they reported evidence that H2S is an O2 sensor in trout chemoreceptors (25). The present study has extended their findings to the mammalian chemoreceptors and has further demonstrated that H2S acts via inhibiting the BKCa channels.
HO-2 was previously proposed to function as an oxygen-sensor via a mechanism involving CO generation from oxygen and increase in the BKCa channel activity (41), although a subsequent study has shown that the chemoreceptor response to hypoxia in HO-2 deficient mice remains intact (26). Meanwhile, other oxygen sensing mechanisms have been put forward (see Introduction). In this study, we found that despite negligible effect on its own, the CO donor could antagonize H2S-evoked afferent chemoreceptor excitation and BKCa channel inhibition (Figs. 3 and 5), confirming the stimulating effects of CO on the BKCa channels. In a recent study, Telezhkin et al. have examined the effects of H2S on recombinant human BKCa α channels (34). Interestingly, they showed that NaHS inhibited the BKCa channel composed of the pore forming α subunit by reducing open state probability without altering its conductance and that the inhibitory effect of H2S was reversed by a CO donor. However, unlike the activation by the CO donor, the inhibition of the BKCa channel by H2S was not sensitive to KCN, suggesting that the actions of H2S and CO are noncompetitive. Regardless of the relationship between H2S and CO signaling pathways in regulating the BKCa channels, the in vitro and in vivo results from this and previous studies support the notion that H2S is crucial in oxygen sensing of the carotid body.
In summary, we provide evidence that H2S activates the carotid body chemoreceptors via inhibiting the BKCa channels and endogenous H2S plays a crucial role in oxygen sensing. Such a mechanism may have widespread physiological and pathological implications, given broad expression of the BKCa channels and strong biological relevance of acute oxygen-sensing in both health and diseases.
Footnotes
Acknowledgments
This work was supported by the Science and Technology Commission of Shanghai Municipal Government (Grant 05PJ14053) and Natural Science Foundation of China (Grant No 30770797 and 30700269).
Author Disclosure Statement
No competing financial interests exist.