
Abstract
Extracellular redox (reduction–oxidation) state is a factor that serves as an important regulator of cell–microenvironmental interactions and is determined by several known variables; including redox-modulating proteins that are located on the plasma membrane or outside of cells, extracellular thiol/disulfide couples, and reactive oxygen species (ROS)/reactive nitrogen species (RNS) that are capable of traveling across plasma membranes into the extracellular space. The extracellular redox state works in concert with the intracellular redox state to control both the influx and efflux of ROS/RNS that may serve to modulate redox signaling or to perturb normal cellular processes or both. Under physiologic conditions, the extracellular space is known to have a relatively more-oxidized redox state than the interior of the cell. During pathologic conditions, such as cancer, the extracellular redox state may be altered, causing specific proteins such as proteases, soluble factors, or the extracellular matrix to have altered functions or activities. Recent studies have strongly supported an important relation between the extracellular redox state and cancer cell aggressiveness. The purpose of this review is to identify redox buffer networks in extracellular spaces and to emphasize the possible roles of the extracellular redox state in cancer, knowledge that may contribute to potential therapeutic interventions. Antioxid. Redox Signal. 13, 449–465.
Introduction: Extracellular Redox State
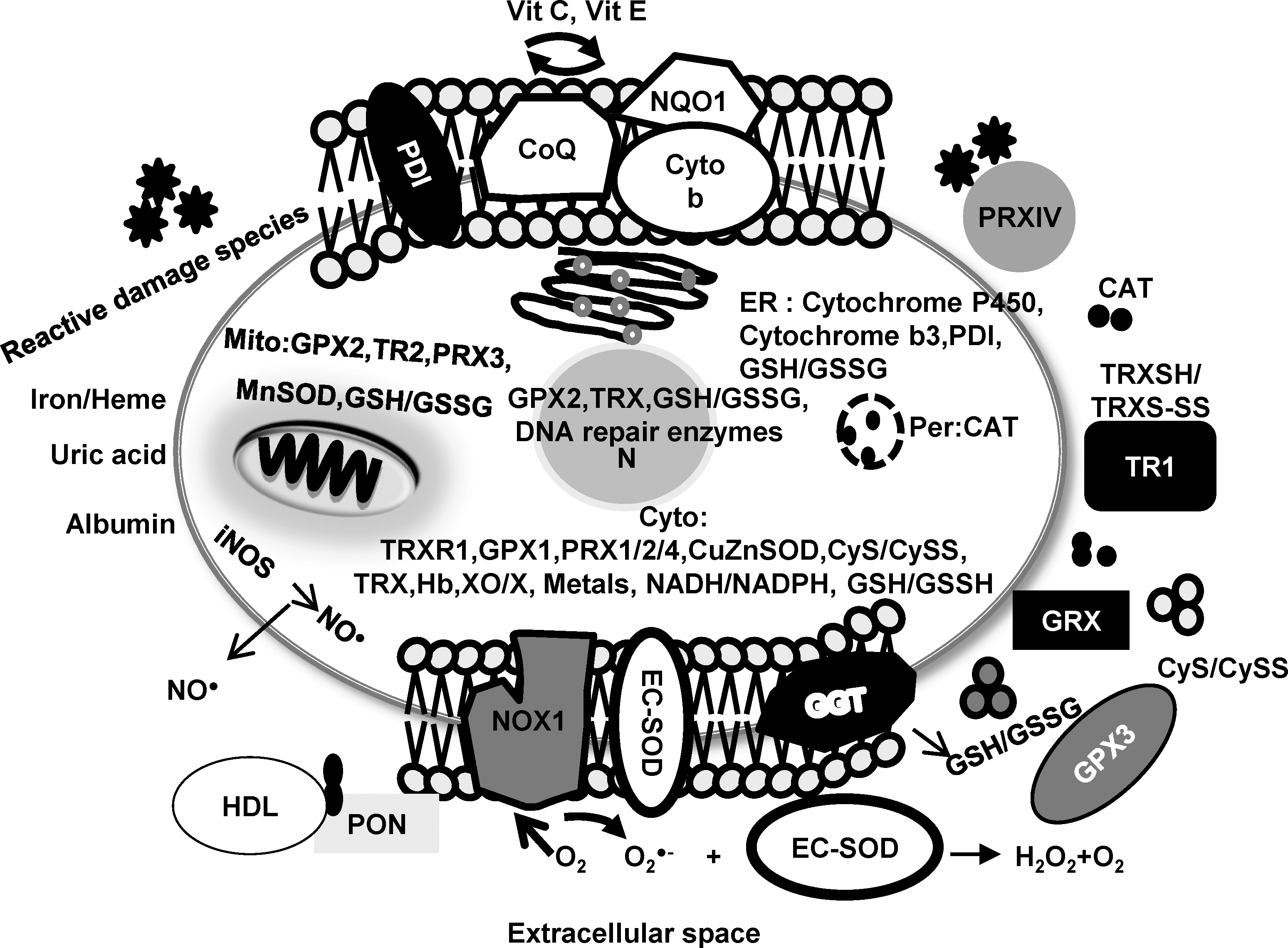
M.W., molecular weight; Ref, reference.
Redox-modulating proteins that are located in the plasma membrane
The plasma membrane protects and separates the cell from its microenvironment. Several physiologic and pathologic signaling(s) pathways are initiated at the plasma membrane. Pathologic conditions resulting in oxidative stress can attack plasma membranes and cause membrane damage.
NADPH oxidase 1
The NADPH oxidase 1 (NOXs) family of NADPH oxidases is composed of proteins that transfer electrons across biologic membranes. In the reaction catalyzed by these enzymes, the electron acceptor is oxygen, and the product of the electron transfer reaction is superoxide radical (O2 •–). The NOX family of NADPH oxidases contains seven structurally related members that are homologous to NOX2 (gp91, the first discovered NOX that is expressed largely in phagocytic cells). These family members include NOX1 through NOX5 and dual oxidases (DUOX) 1 and 2. NOX1-4 isoforms are about 66 kDa in molecular mass, whereas the DUOX(s) are about 180 kDa (8). Among the NOX family members, NOX1 is highly expressed on plasma membranes in several cell types other than phagocytic cells, including colonic epithelium, vascular smooth muscle, placenta, and prostatic epithelium (58, 74, 96).
The immediate product of the NOX1 enzyme is O2 •–, which is generated in both the extracellular and intracellular spaces. The negatively charged O2 •– does not permeate the lipid bilayers of plasma membranes; however, in some cases, O2 •– may pass through the pores of anion channels. Owing to spontaneous and enzymatic dismutation, hydrogen peroxide (H2O2) is also rapidly generated (1). Biochemical effects of NOX-derived O2 •– include reaction with nitric oxide (NO•) leading to NO• degradation or peroxynitrite (ONOO−) formation in extracellular spaces, or reduction of iron centers within enzymes (e.g., extracellular catalase). Biochemical effects of NOX-derived H2O2 include oxidation of extracellular cysteine/glutathione (GSH) and reduced levels of oxidized metal ions (8). Enhanced radical formation in extracellular spaces could lead to imbalance of the extracellular redox state, which may in turn lead to alterations of cell physiologic functions.
γ-Glutamyltransferase
γ-Glutamyltransferase (GGT) is an antioxidant protein catalyzing the first steps in the degradation of extracellular GSH, thus providing cysteine to the cells (91). In contrast, GGT may act as a prooxidant protein by producing extracellular prooxidant species during the GSH degradation process, which is accompanied by several metal-catalyzed redox reactions that may independently affect the proliferative/apoptotic balance of the cell. Moreover, the cysteinyl-glycine products generated through GGT-mediated metabolism of GSH may react with cisplatin (CDDP), a reaction that may lead to formation of CDDP/thiol adducts extracellularly. Thus, upregulation of GGT may result in increased oxidative stress rather than increased intracellular GSH levels. GGT is present in the plasma membranes of almost all cells, but high enzyme levels are expressed in selected cell types, including kidney tubules, biliary epithelium, and brain capillaries (91).
Redox-modulating proteins that are located outside the cell
Unlike the intracellular compartment, extracellular spaces have very low levels of antioxidant enzymes/proteins under physiologic conditions. Little evidence exists of significant levels of primary antioxidant proteins in most extracellular locations, except in plasma, which contains various soluble antioxidant proteins.
Extracellular superoxide dismutase
It is well established that extracellular superoxide dismutase (EC-SOD) is a major extracellular antioxidant protein. It is located in extracellular fluids such as lymph, synovial fluid, plasma, in extracellular matrix, and on cell surfaces (81). EC-SOD is a slightly hydrophobic glycoprotein with an apparent molecular mass of 135 kDa (26). EC-SOD is present in various organisms as a tetramer or, less commonly, as a dimer, or both (84). EC-SOD contains one copper and one zinc atom per subunit (65, 113), and both are required for enzymatic activity. EC-SOD activity can be inhibited by a variety of agents, including azide and cyanide, and inactivated by diethyldithiocarbamate and H2O2 (51, 52). The SOD3 gene is approximately 60% homologous to the SOD1 gene and localizes to human chromosome 4q21 (123).
EC-SOD is found in highest concentrations in blood vessels and lung (64). EC-SOD is synthesized and secreted by a variety of fibroblast and endothelial cell lines (63). One important characteristic of EC-SOD is that it has a strong affinity to heparin (83). Further investigation revealed that the C-terminal domain (positively charged arginines and lysines in amino acids 210–215) is essential for interaction of EC-SOD with heparin and heparin sulfate (90). This region is involved in the binding to type I collagen and is referred to as the ECM-binding region. Plasma EC-SOD can be divided into three types: (a) one with no heparin affinity, (b) one with weak heparin affinity, and (c) one with strong heparin affinity (99). In humans, most tissue EC-SOD exists as type C (99). O2 •– reacts rapidly with NO• to deplete NO• bioactivity and forms a strong oxidant, ONOO−. In addition, O2 •– helps maintain iron in the reduced state, which is then available to participate in Fenton chemistry to produce hydroxyl radical, a highly reactive and toxic ROS. Therefore, EC-SOD limits the levels of these highly reactive ROS/RNS and reduces toxic reactions of these radicals in the extracellular space.
Glutathione peroxidase 3
Like other members of the GPX family, glutathione peroxidase 3 (GPX3) catalyzes the reduction of H2O2 and lipid hydroperoxides by using GSH as a reducing agent. GPX3 is a 23-kDa homotetramer, and its concentration in human plasma is 24–29 mg/L (98). GPX3 has been detected in extracellular body fluids such as thyroid colloid and amniotic fluid (62). To date, the precise reductive cofactor(s) in the extracellular space interacting with GPX3 are unclear. GPX3 has ∼10-fold higher K m for GSH than other isoforms (31). In vitro experiments suggested that thioredoxin (TRXSH) (21) and glutaredoxin (55) may function as electron donors for GPX3 instead of GSH, because its concentration in plasma is only in the micromolar range. GPX3 is expressed in various tissues, with the highest mRNA levels in the kidney. An increase in GPX3 in plasma has been shown to correlate with increased mRNA in the kidney (29). GPX3 has been detected at lower levels in several tissues, such as heart, testis, and liver (66).
Thioredoxin reductase-1
Thioredoxin reductase-1 (TR1), a homodimeric selenium containing flavoprotein, transfers reducing equivalents from NADPH to TRXSH (4, 13). Generally, TR1 is located in the cytoplasm with a molecular mass of 54.6 kDa. Current studies have suggested that TR1 plays an extracellular role (75). An extracellular TR1 form of 116 kDa was found to be secreted from peripheral blood mononuclear cells and monocytes, and this secreted form of TR1 has also been demonstrated in leukemia and melanoma cell lines (105). However, the exact function of the secreted TR1 isoform is unclear. Considered primarily as an antioxidant, it has been suggested that extracellular TR1 may play a role in cell growth and apoptosis by working as a cofactor with other extracellular proteins such as TRXSH1 or the peptide GSH.
Extracellular thiol/disulfide couples
The extracellular supply of thiols is critical for the maintenance of the extracellular space/microenvironment redox state. The importance of cell-surface/extracellular thiols in many cellular functions, including ligand-receptor binding and signal transduction, has been demonstrated in several cell types. Crosstalk between thiol/disulfide couples controls various types of cellular functions.
Cysteine/Cystine
The cysteine (CyS)/cystine (CySS) couple is the major low-molecular-weight redox couple present in the microenvironment and is one of the central redox-control nodes in biologic systems (50). The major oxidized form, CySS, exists predominantly in plasma, and the CyS/CySS ratio is used as a measure of the extracellular redox state. CySS has been found to be approximately 120 μM in the plasma of healthy adults (50), suggesting that influx and efflux of CyS may contribute to plasma redox status and keeps the oxidized form at low concentrations. Evidence from various sources suggests that cells can acquire CyS/CySS mainly by two routes. CySS, the predominant form in the circulation, can be taken up through the xc − cystine/glutamate antiporter, a plasma membrane CySS transporter that cancer cells can express, in particular when they are more aggressive (60). Alternatively, cells can readily take up extracellular CySS as produced, for example, by neighboring activated macrophages, dendritic cells, and fibroblasts. Such stromal cells take up CySS through the CySS transporter, reduce it to CyS, and secrete CySS into their microenvironment (3, 28).
In humans, the physiologic CyS/CySS redox potential (E h CyS/CySS) in healthy subjects is around −80 mV, whereas in subjects with disease, this redox state becomes oxidized to between −62 and −20 mV (46, 49, 69, 80). Several recent studies demonstrated that modification of extracellular E h CyS/CySS could directly regulate or stimulate the differential expression of genes that control behavior, including cell proliferation and cell invasion. Extracellular CyS/CySS may thus act as an oxidant redox switch in several physiologic events.
Glutathione/Glutathione disulfide
Glutathione (GSH) is a strong reducing agent that contributes to key antioxidant metabolic pathways by acting either as a proton donor or as a cofactor of nucleophilic conjugates. This tripeptide of serine, cysteine, and glutamate serves to maintain the redox integrity of the inside and outside of cells. GSH serves as an intracellular thiol redox buffer, maintaining the intracellular environment in a reduced state. Plasma contains only low levels of GSH, 2–4 μM (49), unlike the intracellular space, where it is present at millimolar concentrations (39). Airway-lining fluid is unusual among extracellular fluids in containing GSH at high micromolar concentrations (200–400 μM) (12). Because GSH degradation occurs extracellularly, the export of GSH, glutathione disulfide (GSSG), and GSH adducts is an important step in its turnover.
Thioredoxin/Thioredoxin disulfide
Thioredoxin (TRXSH) is a small 12-kDa redox-active protein that serves as an electron donor to reduce inactive oxidized forms of proteins back to reduced active forms. The active-site sequence (CyS-Gly-Pro-CyS) of the protein is conserved from Escherichia coli to human. By using the two cysteine residues in the active site, TRXSH/thioredoxin disulfide (TRXS-S) acts as an oxidoreductase through the dithiol/disulfide moiety (67). The TRXSH/TRXS-S system plays a major role as an intracellular antioxidant and may also play a significant role in decreasing ROS levels extracellularly. Extracellularly, TRXSH levels are in the 1- to 5-nM range (67). TRXSH has been found at a concentration of 30 ng/ml by using sandwich ELISA technique in human plasma (88). Full-length and truncated TRXSH have been detected in plasma and may provide further clues to the probable role as secretory antioxidant proteins (89). Both the long and truncated forms of the protein have been localized to the cell surface, and it has been suggested to have an antioxidant role in the extracellular environment. In addition, TRXSH may serve as an electron donor for the potential peroxidase activity of albumin (54).
Extracellular ROS/RNS
The source of ROS/RNS in the extracellular space is in the intracellular compartment, as well as in the extracellular space and from surrounding cells. As most ROS/RNS have limited diffusion distances because of their highly active reaction rates with biologic molecules, most of ROS/RNS generated within a cell cannot diffuse out of the cell. The concentrations of types of ROS/RNS generated extracellularly may differ from those generated within a cell.
Hydrogen peroxide
Hydrogen peroxide
Nitric oxide
Nitric oxide (NO•) is synthesized from the amino acid
Initial measurements in plasma reported levels of 3 nM of free NO• and ∼7 μM for NO• present as S-nitrosation derivative molecules (2, 92). NO• at high concentrations is a highly toxic compound; it reacts with O2 •– to yield ONOO−. ONOO− is a potent oxidant that can mediate the oxidation of biomolecules such as thiols. NO• may also have an important role in regulation of redox-sensitive thiols in the intracellular and extracellular environments, linking intra- and extracellular redox states as a network that communicates to regulate cellular function.
Extracellular free radical–damage products
The extracellular space/microenvironment of most cells is significantly less sheltered against free radical damage than that inside the cells because of the limited amounts of antioxidant-related proteins/enzymes. Several studies document oxidative/nitrative modifications of proteins, lipids, or carbohydrates and subsequent accumulation of these modified molecules in extracellular spaces during pathologic-related oxidative stress, an example being accumulation of damage to methionine residues (19). The major consequences of reactive damage in the extracellular space are damage to the ECM and to plasma membranes. Oxidative/nitrative damage may change the ECM and the plasma membrane structure, both of which play a key role in the regulation of cellular adhesion, proliferation, migration, and cell signaling. In addition, ECM and plasma membranes are widely recognized as being key sites of cytokine and growth-factor binding; thus, modifications of these structures alter cell behavior.
Extracellular repair systems
In addition to extracellular antioxidant proteins, the extracellular repair systems also play role(s) in the protection of the extracellular space/microenvironment. However, the levels and activities of extracellular repair systems appear to be significantly limited when compared with intracellular repair systems. Additionally, slow turnover of many extracellular proteins can result in an accumulation of free radical damage modifications. Oxidative carbohydrate damage can be repaired in the extracellular space through catabolism (32). In contrast, extracellular oxidative/nitrative protein damage has a limited capacity to be repaired, because the extracellular space/microenvironment does not contain Met sulfoxide reductase activity (19). However, the extracellular space contains disulfide reductase and protein disulfide isomerase enzymes (PDIs) on cell surfaces (45, 47). These enzymes have been shown to repair or correct protein conformation between reduced and oxidized thiols on cell surfaces/extracellular spaces. Unfortunately, some extracellular protein modifications may not be recognized and repaired by PDIs. Alternatively, extracellular protein damage may be repaired by catabolism. However, protein–protein cross links due to oxidation–reduction may limit catabolism processes by preventing cell uptake of damage products.
Possible Roles of the Extracellular/Microenvironment Redox State in Cancer
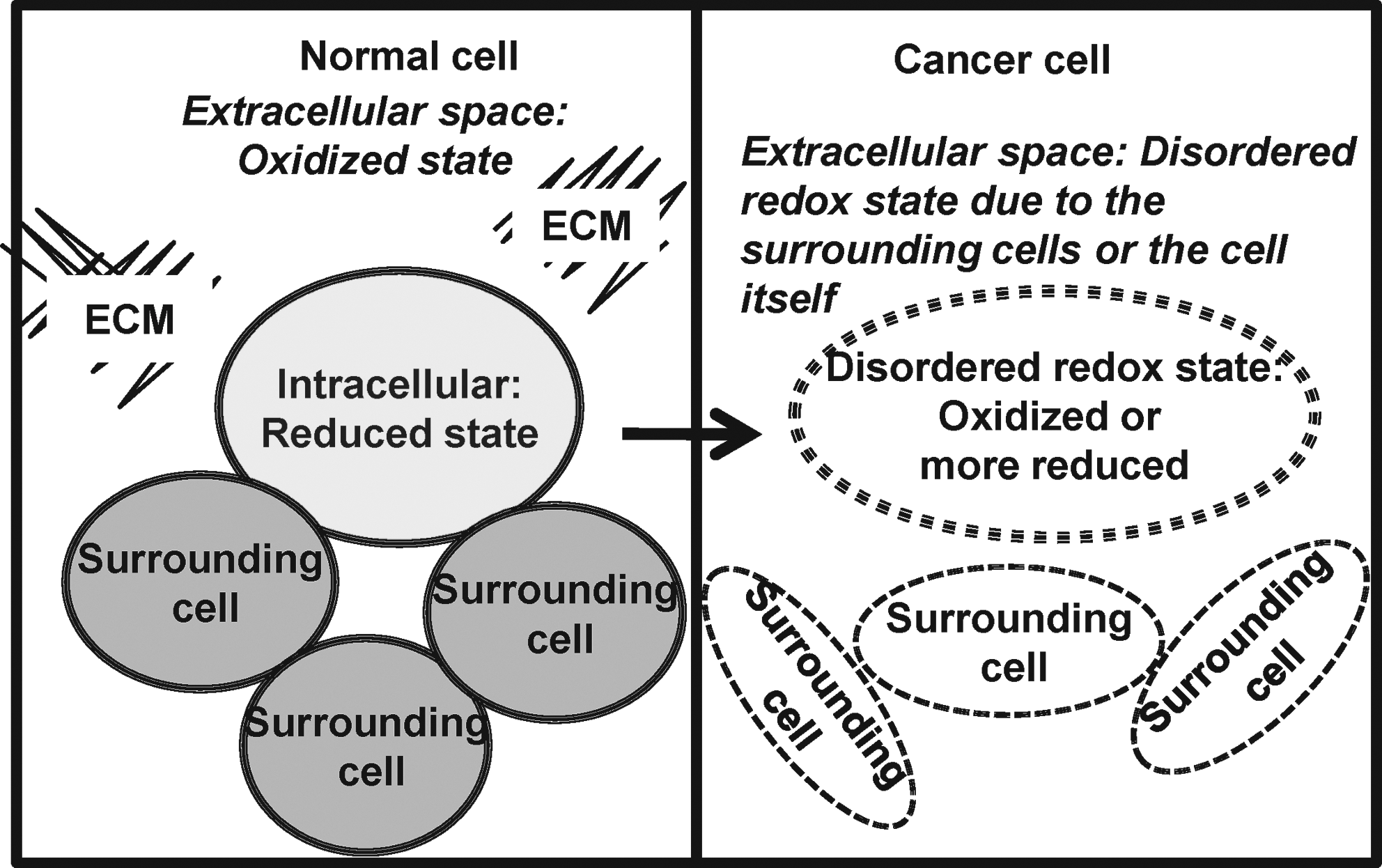
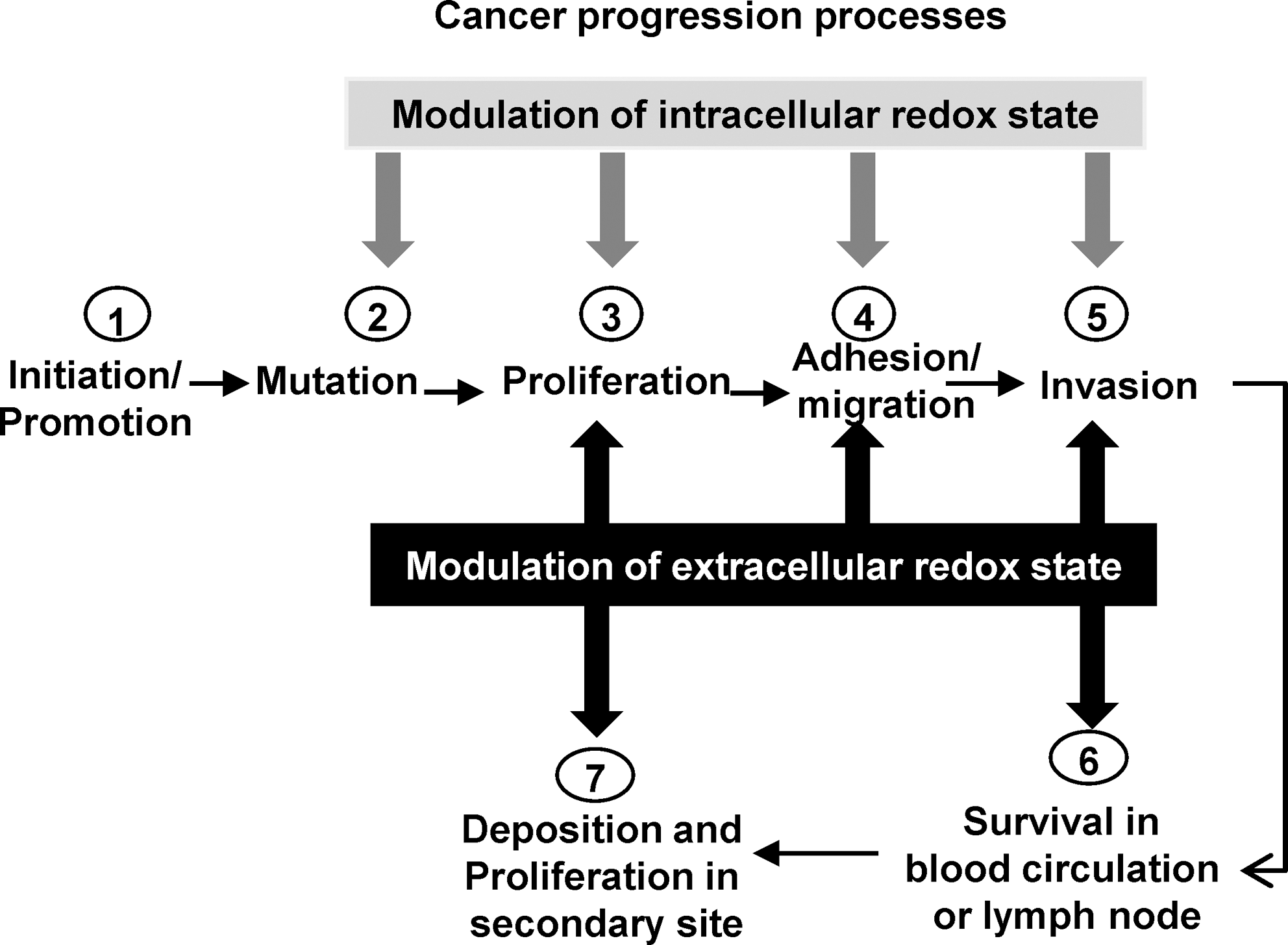
In general, the balance of the extracellular redox state of the cell is determined by the intracellular redox state or signal transduction in surrounding cells or both. Persistent generation of ROS/RNS or alterations of redox-modulated proteins in the extracellular space/microenvironment (or both) may lead to disturbances of the extracellular redox state. These changes have been linked to several pathologic processes, including cancer. We hypothesize that the redox-state balance of the extracellular space/microenvironment is altered during cancer progression (Fig. 2) and may be an important factor for regulation of cancer cell behavior during the metastatic process. By measuring extracellular redox-related proteins, GSH/GSSG levels, and ROS/RNS levels in in vitro studies, our laboratory demonstrated that the redox state of the extracellular space/microenvironment was altered in prostate cancer cells [Figs. 3 –5, (16)]. Different cancer cell types or stages of progression of the same cancer type may demonstrate different and unique extracellular redox states. We hypothesize that cancer cells may exhibit a plasticity of redox state in the extracellular space/microenvironment, so that the conversion between status (reduced or oxidized) in the extracellular space/microenvironment can be accomplished rapidly. Moreover, the redox state of the extracellular space/microenvironment may be an important factor in determining the response of cancer cells to chemotherapy or radiation. This review addresses important findings linking extracellular redox state and cancer. The role of low-molecular-weight antioxidant compounds may be found in other recent reviews (30, 36, 44).
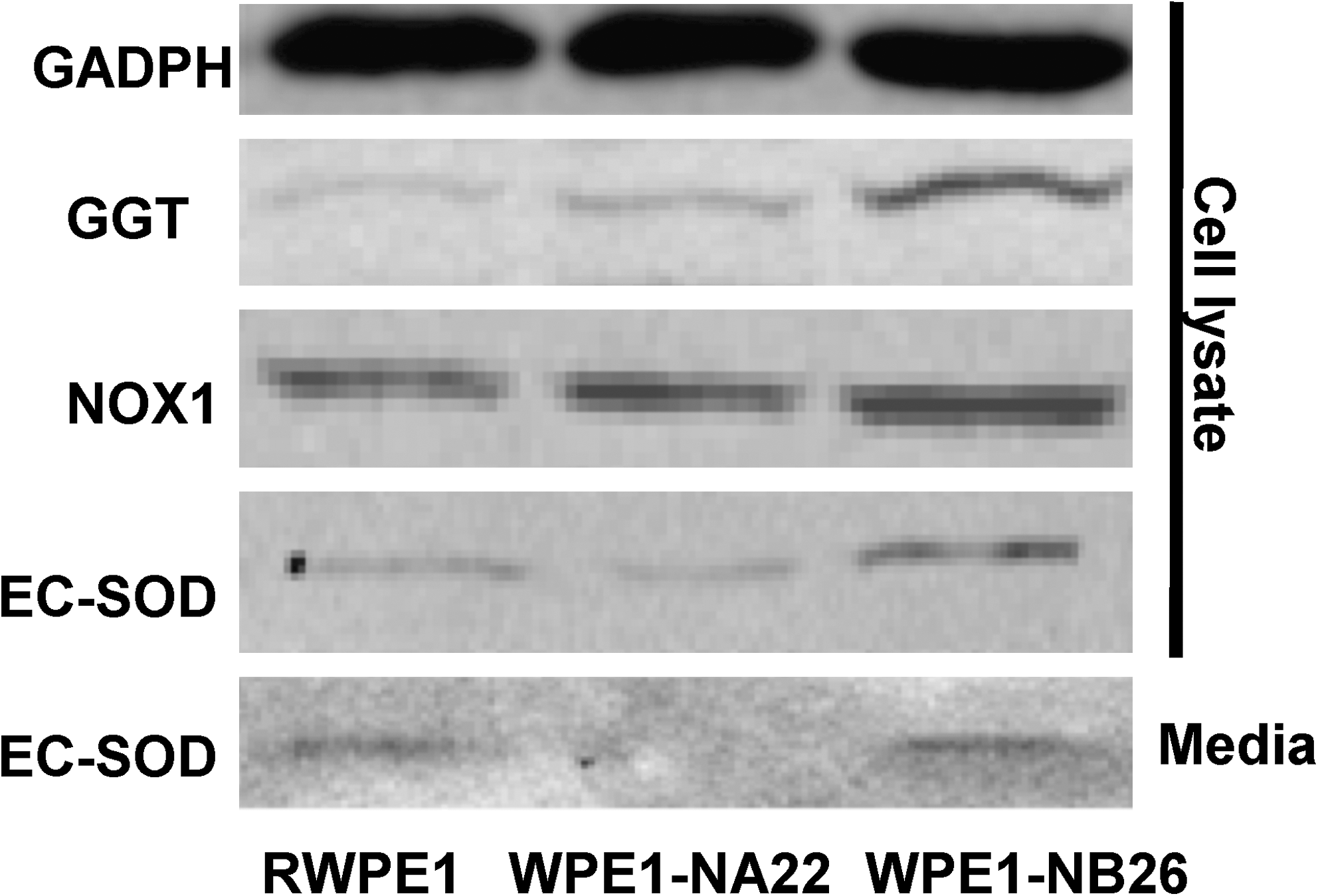

NOX1
Because ROS/RNS are important factors in regulating cancer cell growth, several investigators have studied the expression of NOX1 in cancer cells. NOX1 has been suggested to participate in growth factor–triggered extracellular ROS production (114). Increased extracellular ROS generation (O2 •– and H2O2) is mainly due to NOX1 activity. In a tissue-culture model, less-transformed human keratinocyte cells (EPIs) expressed low levels of Nox1 protein (17), whereas more-transformed cells (FIBs) had higher levels of Nox1 protein expression, as measured by using Western blot analysis (18). Moreover, NOX1 mRNA was absent in normal stomach cells with PCR techniques; in contrast, significant levels of NOX1 mRNA were detectable in gastric cancer cells (114). Arnold et al. (58) investigated the correlation between NOX1 and ROS in prostate cancer and found that human prostate tumors showed marked increases in Nox1 protein and mRNA levels. The induction of Nox1 protein was correlated with an increase in H2O2 levels. In addition, a series of cell lines developed from LNCaP prostate cancer cells demonstrating increasing tumor and metastatic potential showed increased Nox1 protein levels and a parallel increase in H2O2 levels (58). Our laboratory demonstrated induction of Nox1 protein expression in the highly aggressive WPE1-NB26 prostate cancer cells in comparison to immortalized RWPE1 prostate epithelial cells (Fig. 3). The induction of Nox1 protein expression in WPE1-NB26 cells correlated with extracellular H2O2 levels, with higher H2O2 levels in the extracellular space (cell-culture medium) of WPE1-NB 26 cells than in the extracellular space of RWPE1 cells (Fig. 5).
In colon cancer, NOX1 mRNA expression, as assessed by using PCR, was maximal in well-differentiated adenocarcinoma and lesser in poorly differentiated adenocarcinoma, implying that NOX1 expression is inversely correlated with cancer progression (96). Conversely, a study of patient samples showed that NOX1 mRNA expression measured by dot-blot hybridization, real-time PCR, and in situ hybridization was not statistically higher in colon cancer than in normal colon tissues (109). One possible explanation for seemingly contradictory results is that NOX1 may exert cancer-promoting effects but may be less involved in cancer progression. Because studies have shown variable levels of NOX1 in various cancer and model systems, controversy still exists regarding the role of NOX1 in carcinogenesis.
GGT
GGT catalyzes the degradation of extracellular GSH, which is then subsequently metabolized into CyS inside of cells. Many cancer cells require CyS for growth. Therefore, the relation between GGT and cancer formation has been widely studied. In leukemia K562 and MEG-01 cell lines, the proinflammatory cytokine tumor necrosis factor α induced GGT-promoter transactivation, mRNA, and protein synthesis, as well as enzymatic activity (94). Serum GGT has been demonstrated to be associated with a high risk of liver cancer (40); thus, it has been used as a serum marker of hepatocarcinoma. In some tumors, GGT expression has been established as an early marker of neoplastic transformation, an example is skin cancer. It has been demonstrated that there was no expression of GGT protein assessed by immunohistochemistry in benign papillomas (20), basal cell carcinoma, or other benign skin tumors; in contrast, a high expression of GGT was found in squamous carcinoma (34). Thus, GGT expression in cancer may correlate with an aggressive phenotype. Our Western blot analysis in prostate cancer cell lines demonstrated that GGT expression levels in highly aggressive WPE1-NB26 prostate cancer cells was higher than in less-aggressive WPE1-NA22 prostate cancer cells and immortalized RWPE1 prostate epithelial cells (Fig. 3). Analyzed by using immunohistochemistry, GGT protein was shown to be expressed in non–small cell cancer but not in hyperplasic bronchial mucosa (91). Transfection of a murine epidermal cell line with GGT cDNA resulted in a significant increase in transplanted cell-line growth in nude mice (119). Exposure of CC531 colon carcinoma cells to a low dose of gamma radiation resulted in increased GGT activity through RAS activation (86). However, some studies demonstrated a significant reduction of GGT in human tumors. The fact that GGT-mediated metabolism of GSH may be accompanied by metal-catalyzed redox reactions that subsequently produce prooxidant species may explain why certain cancer cells express less GGT.
EC-SOD
Among the extracellular antioxidant proteins, the role of EC-SOD is well established because of its ability to be secreted outside of cells. Changes in EC-SOD protein expression have been associated with various types of cancer. Our laboratory demonstrated that the levels of EC-SOD protein expression in highly aggressive WPE1-NB26 prostate cancer cells was slightly higher when compared with those in immortalized RWPE1 prostate epithelial cells both outside (in the media) and inside the cells (Fig. 3) (16). In contrast, immunohistochemical assessment of EC-SOD in lung carcinoma tissues demonstrated a positive EC-SOD immunoreactivity in only two of 37 cases (108). Lower expression of EC-SOD protein levels also was found in renal cell carcinoma when compared with those in normal renal tissues (106).
GPX3
In cancer cells/tissues, GPX3 is usually downregulated. It has been reported that renal cell carcinoma tissues had no detectable levels of GPX3 in comparison to normal renal tissues when analyzed with two-dimensional polyacrylamide gel electrophoresis (101). Gpx3 protein expression was reported to be downregulated in thyroid cancer (103), colorectal cancer (73), and prostate cancer (122). The downregulation of Gpx3 protein in prostate cancer cells/tissues was associated with hypermethylation of the GPX3 gene (122), whereas in thyroid cancer, decreased levels were associated with downregulation of the transcription factor PAX8 (103). In addition to protein expression, GPX3 activity was demonstrated to be significantly reduced in the blood of patients with breast, gastric, and colorectal cancers (87). A major function of GPX3 is to protect the extracellular space/microenvironment from H2O2, a molecule that can stimulate cancer cell growth at appropriate concentrations. Therefore, GPX3 has been considered to be a tumor-suppressor protein. The study of Nkx3.1 knockout mice demonstrated that GPX3 protein expression was significantly but only transiently induced during transition from normal epithelium to prostatic intraepithelial neoplasia (PIN); once PIN developed into carcinoma, Gpx3 protein expression was absent (85). The absence of GPX3 expression has been correlated with increased invasive ability and oxidative-damage levels in prostate cancer (122).
GSH/GSSG
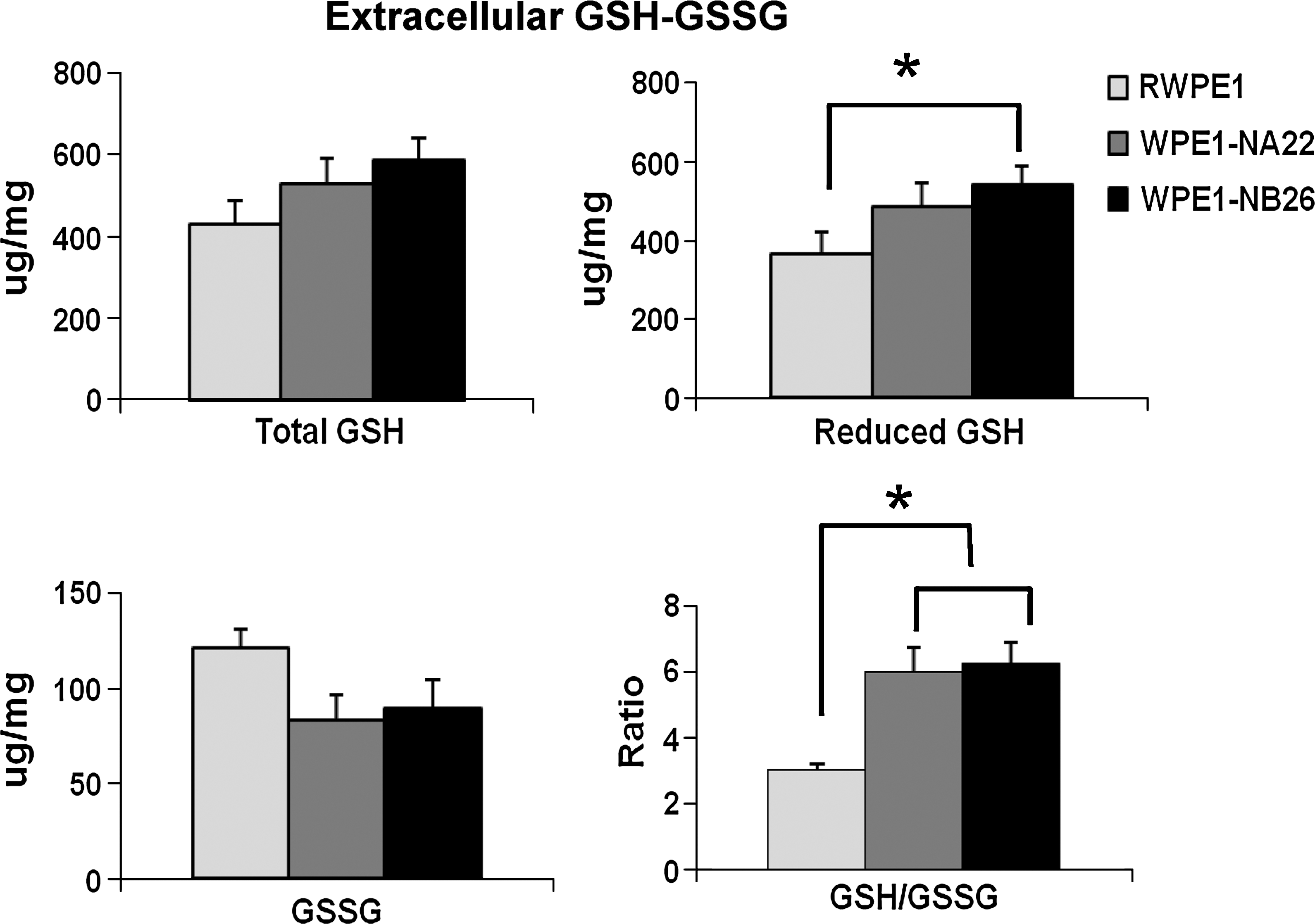
The major role of extracellular GSH/GSSG is to maintain the balance of the extracellular redox state. By measuring extracellular GSH/GSSG ratios (in the culture media), we demonstrated that, in highly aggressive WPE1-NB26 prostate cancer cells, extracellular GSH/GSSG ratios were higher when compared with those in immortalized RWPE1 prostate epithelial cells (Fig. 4) (16). In addition, extracellular GSH/GSSG ratios in the highly aggressive PC3 prostate cancer cells were higher when compared with those in less-aggressive LNCaP prostate cancer cells (14). From these results, we infer that prostate cancer cells had a more-reduced state in the extracellular space/microenvironment to maintain or develop a more-aggressive phenotype.
CyS/CySS
Several cancer cell types are incapable of synthesizing CyS/CySS, including leukemias and lymphomas. CyS is an amino acid often required for cancer growth and viability and is also an important redox-related component in the maintenance of the redox state and GSH levels in the extracellular space/microenvironment. In cancer cells, CyS/CySS pools are not in equilibrium, and a reduction in uptake of extracellular CyS/CySS has been demonstrated in breast carcinoma (76), leukemia (41), and prostate cancer cells (25). It has been demonstrated that PC3 cells compensate for the reduction in uptake of extracellular CyS/CySS by overexpression of the CySS plasma-membrane transporter (xc − cystine/glutamate antiporter) (25). The main functions of this transporter are mediation of cellular CySS uptake for synthesis of GSH essential for cellular protection from oxidative stress and maintenance of redox balance in the extracellular compartment.
It has been demonstrated that alteration of extracellular CyS/CySS levels of Caco-2 colon carcinoma resulted in reduced redox potential (−80 mV) and reduced CyS/CySS concentrations correlating with regulation of proliferation of these cells (48). Alteration of the extracellular CyS/CySS redox state altered phosphorylation of p44/p42 mitogen-activated protein kinase through epidermal growth factor–receptor phosphorylation, with the greatest phosphorylation at the most-reduced condition (−150 mV) (79). The data to date indicate that alterations of extracellular CyS/CySS concentrations may be an important feature of cancer cells during progression. In contrast, a study of lung fibroblasts showed that oxidation of CyS in the media stimulated cell proliferation and ECM expression (43). These latter results suggested that extracellular CyS/CySS redox-dependent cell proliferation may be cancer or cell-type specific.
TRXSH1
TRXSH1 is located intracellularly; however, in patients with hepatocellular carcinoma, serum levels of TRXSH1 have been detected (68). TRXSH1 appears to be released directly from cancer tissues, because serum TRX1 levels decreased after removal of the cancer; thus, measurement of serum TRXSH1 might prove to be a useful clinical parameter for detection of hepatocellular carcinoma. Studies have demonstrated that TRXSH1 is secreted from both normal and Chinese hamster ovary cancer cells; however, because TRXSH1 secretion was elevated in cancer cells, the secretion was hypothesized to stimulate cancer growth by enhancing the activity of other growth factors (110). TRXSH1 was identified as an autocrine growth factor produced by Epstein-Barr virus–transformed B cells or the T-cell hybridoma MP6 (97, 117). TRXSH1 was reported to show mitogenic activity in leukemia cells (118); however, the mitogenic activity was significant only in a reduced extracellular space/microenvironment. In media of cell cultures or in plasma, TRXSH1 is easily oxidized; therefore, it is important for cancer cells to maintain an appropriate redox state in the extracellular space/microenvironment for optimal activity of this cancer-related protein. Cancer cells may rely on TRXSH1 to protect against oxidative stress, and the secretion of TRXSH1 may be an important determinant in resistance of cancer cells to chemotherapy.
H2O2/O2 •–
The role of extracellular H2O2 has been viewed in terms of threshold concentration, serving a dual role as a source of oxidant stress at high concentrations and as a signaling molecule at low concentrations. De Yulia et al. (22) demonstrated that extracellular H2O2 was produced by receptor-ligand interactions in growth-factor signaling pathways (22). Several studies suggest that extracellular H2O2 may play a role in regulation of vascular function, including stimulation of smooth muscle–cell proliferation, and thus, it may play a role in angiogenesis in cancer formation (61). Our study in prostate cancer cell lines demonstrated a slight increase in extracellular H2O2 levels in highly aggressive WPE1-NB26 prostate cancer cells when compared with immortalized RWPE1 prostate epithelial cells (Fig. 5). Although it was initially believed that H2O2 can freely diffuse across membranes, recent genetic evidence suggests that some membranes are poorly permeable to H2O2 and that its transport may be regulated by AQP channel proteins; thus, extracellular H2O2 may remain outside the cancer cell because of limited diffusion, resulting in higher levels of extracellular H2O2 and subsequent mitogenic effects.
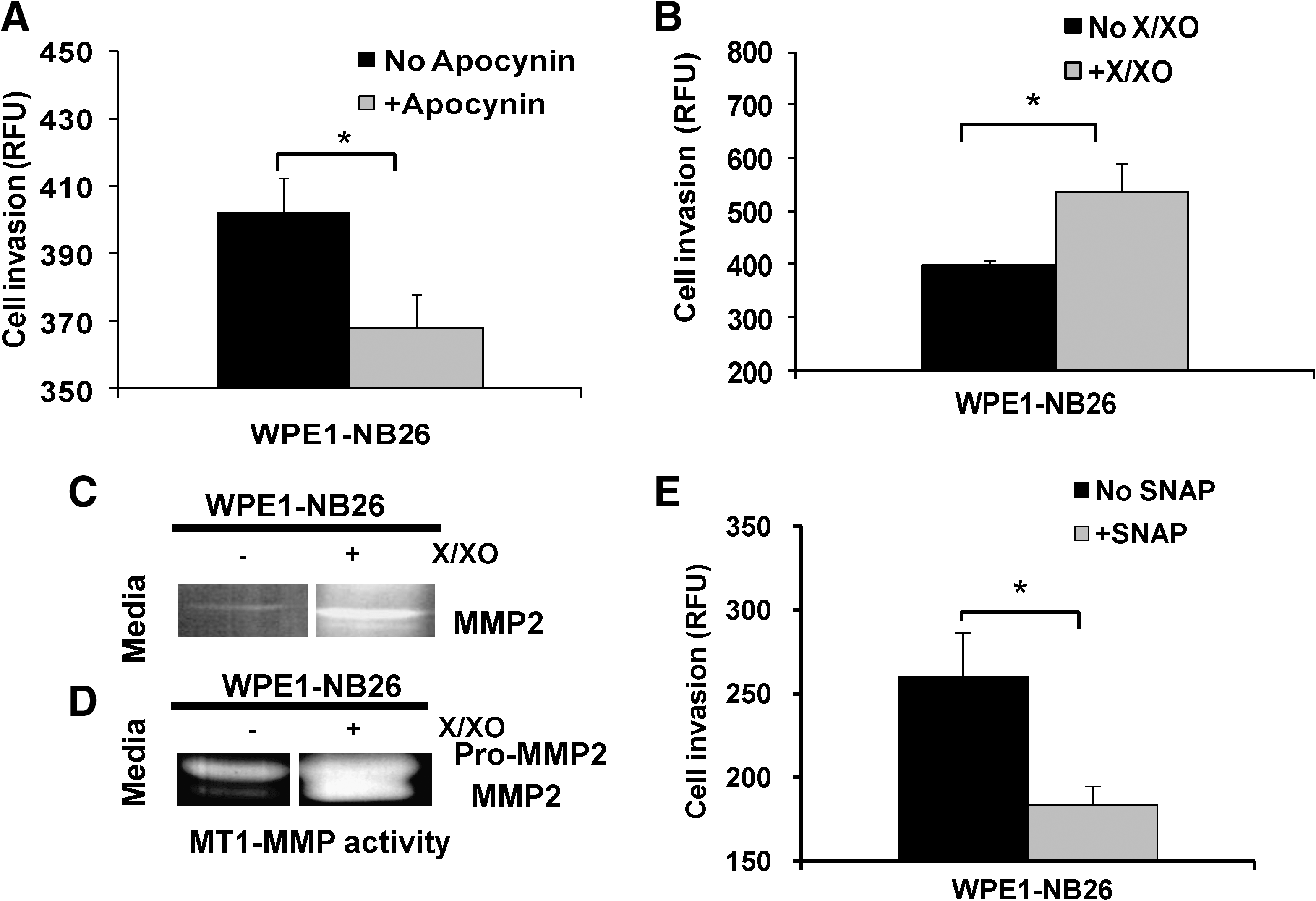
Unlike other ROS/RNS species, extracellular O2 •– has not been widely studied because of its short half-life and extreme difficulty in analyzing levels in the extracellular space. By using xanthine (X)/XO to produce O2 •– in the media, we were able to demonstrate that O2 •– increased the invasive ability of WPE1-NB26 prostate cancer cells, at least partially through induction of matrix metalloproteinase 2 (MMP2) and membrane type 1 (MT1)-MMP activities (Fig. 6B–D) (16). The induction in invasive ability of cancer cells could due to the production of O2 •– at the cell membrane or in the extracellular space or both.
NO•
Recent reports concluded that several human tumor cell lines produce NO• constitutively or in response to exogenous stimuli. Like other biologic events, the production of NO• can be heterogeneous in different tumor cell types. Dong et al. (24) demonstrated that nonmetastatic melanoma cells exhibited high levels of inducible NOS (iNOS) activity and NO•, whereas metastatic melanoma cells did not. In addition, human colon carcinoma cells from the same patient isolated from metastases (SW-620) demonstrated lower NO• levels than cells isolated from the primary tumor (SW-480) (93). In contrast, Gonzalez et al. (27) analyzed expression in 22 primary breast tumors and found a very strong correlation between the presence of iNOS/NO• and axillary lymph node metastasis. Our study in prostate cancer cell lines demonstrated a slight but not statistically significant increase in extracellular NO• levels in highly aggressive WPE1-NB26 prostate cancer cells when compared with those in immortalized RWPE1 prostate epithelial cells (Fig. 5). The role(s) of NO• in cancer progression is complex, with several studies indicating that NO• inhibits tumor cell growth and invasion, whereas other studies suggest that the presence of NO• in the tumor microenvironment promotes tumor angiogenesis and metastasis.
Modulation of Extracellular Redox State and Cancer Behavior
Several extracellular redox-related proteins display tumor-suppressive properties, whereas others display tumor-promotion properties. This section reviews studies of modulation of the extracellular redox state and cancer behavior.
NOX1
Cancer cells expressing NOX1 represent a promising target for selective antitumor therapy. Many cancers are associated with overproduction of ROS due to transcriptional activation of the NOX1 gene. Studies have confirmed the presence of NOX1 on the cell surface of many invasive human cancers (17, 18). Development of anticancer drugs by using NOX1 inhibition as one of the underlying mechanisms must be experimentally tested. We have demonstrated that the invasive ability of highly aggressive WPE1-NB26 prostate cancer cells was decreased after treatment with the NOX inhibitor, apocynin (Fig. 6A) (16). Anti-sense inhibition of Nox1 protein in prostate cancer cells has been demonstrated to suppress proliferation and trigger apoptosis, whereas overexpression of Nox1 protein in human prostate cancer cells has been shown to increase tumorigenicity (5). Treatment with sulfonylurea (LY181984) in patients with solid cancers (e.g., breast, prostate, lung, ovary), leukemias, lymphomas, and in HeLa cells resulted in reduction of NOX1 activity (71, 72). Several NOX inhibitors, including quinine-site inhibitors (70) and antitumor sulfonylureas, are under preclinical development; these compounds inhibit both NOX1 and growth of cancer cells at potentially therapeutic dosage levels without inhibiting the growth of noncancer cells (71, 72).
GGT
Differential expression of GGT occurs in several cancer types and thus might be exploited to increase the therapeutic efficacy of selected treatments. The major function of GGT is the reconstitution of intracellular GSH; thus, GGT expression in cancer cells may participate in mechanisms of drug resistance. Exposure of ovarian cancer cells to cisplatin leads to the appearance of cisplatin-resistant sublines, and the degree of resistance is correlated with increased expression of GGT mRNA levels (56). Drug resistance due to GGT expression in cancer also has been demonstrated in leukemia (1), prostate cancer (38), and melanoma cells (37). It has been demonstrated that GGT enzymatic activity can exert prooxidant effects at the membrane-surface level and in the extracellular microenvironment (91). Generation of prooxidant species could lead to endogenous oxidative stress, which may subsequently result in reduction of GSH levels (91). It has been shown that induction of H2O2 in GGT-expressing lymphoma cells resulted in modulation of thiol groups of cell-surface proteins or receptors or both (23). GGT-expressing preneoplastic nodules induced in rat liver by diethylnitrosamine and phenobarbital demonstrated increased oxidative damage product levels, which may potentially result in the progression of preneoplastic lesions to malignancy (107). The role(s) of GGT activity in cancer biology and therapy must be further analyzed, because it can act as antioxidant and prooxidant.
EC-SOD
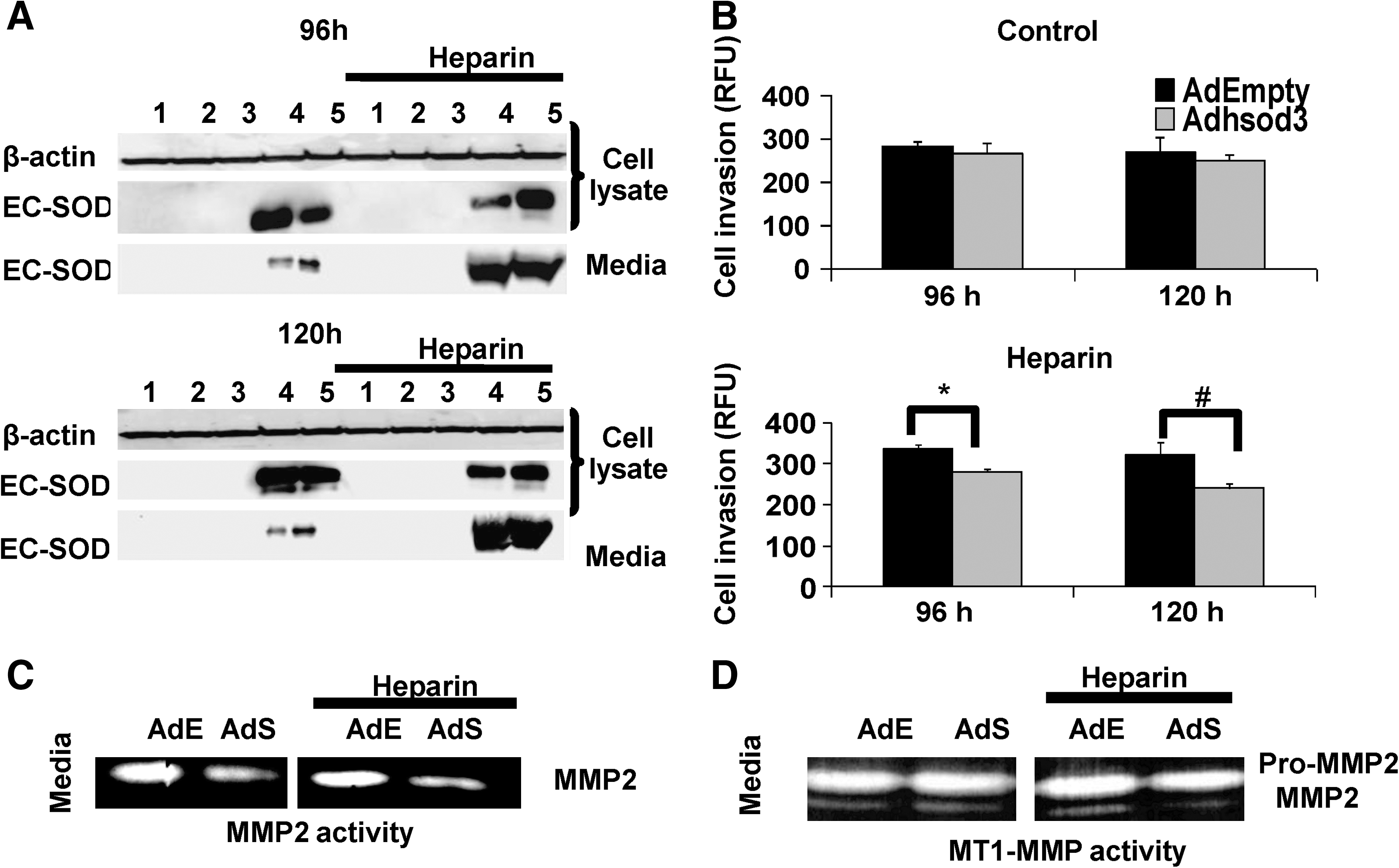
EC-SOD expression has been documented to be altered in several types of cancer. EC-SOD is an antioxidant protein of particular interest in redox-based cancer therapeutics because it can display both antioxidant and prooxidant properties, and a potential role in cancer metastasis has been suggested. Studies demonstrated that EC-SOD has a longer half-life in the circulation relative to the other two SOD isoforms (33, 53). Studies have shown that overexpression of EC-SOD protein inhibited in vivo growth of melanoma (120) and suppressed breast carcinoma growth and invasion (111). A recent study indicated that overexpression of EC-SOD protein significantly inhibited pancreatic cancer cell growth both in vitro and in vivo, whereas manganese superoxide dismutase (MnSOD) and copper zinc superoxide dismutase inhibited only in vitro growth (112). Our laboratory documented that overexpression of extracellular EC-SOD resulted in inhibition of prostate cancer cell growth and invasive ability. We used adenoviral vectors to transfer the functional human SOD3 gene into prostate cancer cells and examined the roles of intra- versus extracellular redox states on prostate cancer cell behavior. To modulate the extracellular redox state, we added heparin to stimulate secretion of EC-SOD protein into the culture media; without heparin treatment, the majority of EC-SOD protein remained inside the cells, thus primarily affecting the intracellular redox state. Western blot analysis of EC-SOD protein documented successful overexpression with two bands identified at ∼33 kDa and ∼28 kDa, the latter band thought to be the result of intracellular proteolysis of the C-terminal end of EC-SOD (Fig. 7A) (16). We analyzed cell growth and invasive ability by using an in vitro cell-invasion assay; we found that overexpression of extracellular EC-SOD protein decreased WPE1-NB 26 cell-invasive ability >50%. Inhibition of cell-invasive ability by overexpression of EC-SOD protein correlated with reduction of MMP2 and MT1-MMP activities (Fig. 7B–D) (16). We also examined the effect of extracellular redox on cell invasion in the immortalized RWPE1 prostate epithelial cells and found no effect of EC-SOD protein on the minimal in vitro cell invasion observed in this cell line. Proposed role(s) of overexpression of extracellular EC-SOD protein in regulation of cancer cell behavior is illustrated in Fig. 8.

GPX3
The distinctive feature of GPX3 is that it is readily detectable in plasma, suggesting that it provides the first line of defense against extracellular ROS. GPX3 was found to be one of the most consistently downregulated genes in prostate cancer with microarray analysis; methylation analyses indicated that the GPX3 gene was widely methylated in prostate cancer. Forced overexpression of Gpx3 protein in prostate cancer cell lines has been demonstrated to suppress tumor growth and metastasis both in vitro and in vivo (122). Overexpression of Gpx3 protein downregulated the expression of c-met, a receptor tyrosine tumor-transforming gene involved in a variety of cellular processes; this study suggests that GPX3 contains tumor-suppressor activity (122). Restoration of wild-type Gpx3 protein expression in tumor cells by gene-targeted therapy should be considered in cancer models of treatment or prevention.
CyS/CySS
Plasma extracellular CyS/CySS becomes more oxidized during several oxidative stress–related pathologic events, including chemotherapy. Thus, modulation of extracellular CyS/CySS could be important in developing systemic cancer therapeutic agents. We have performed a series of experiments modulating concentrations of CyS/CySS in tissue-culture media and studied the resultant effects on normal PrEC prostate epithelial cells or highly aggressive DU145 and PC3 prostate cancer cells. We found that media with oxidized CyS/CySS enhanced prostate cancer cell growth, whereas media with reduced CyS/CySS increased prostate cancer cell invasion. Conversely, neither oxidized nor reduced CyS/CySS affected normal prostate epithelial cell growth or altered behavior in in vitro invasion assays (unpublished data). Similarly, Nkabyo et al. (80) found that media with more-oxidized CyS/CySS resulted in less proliferation of colon cancer Caco-2 cells. The CyS/CySS couple may function as an oxidant–reductant in redox switching, thus providing a means to oxidize–reduce proteins without direct involvement of more potent oxidants–reductants.
A growth requirement of extracellular CyS/CySS by cancer cells can be readily demonstrated by transferring them to culture medium specifically deficient in the amino acid cysteine and monitoring their growth (unpublished data, 25). The apparent importance of the xc − cystine/glutamate antiporter in the supply of CyS to various cancer cells led to an early suggestion that the xc − cystine/glutamate antiporter represents a potential target for therapy. A specific inhibitor of the xc − cystine/glutamate antiporter, sulfasalazine, decreased GSH levels (>90%) in human DU145 and PC3 prostate cancer cell lines; additionally, intraperitoneal administration of this drug substantially reduced growth of xenografts of these cell lines in SCID mice (25). Similar results were obtained in Mia PaCa-2 and Panc-1 pancreatic cancer cells (59), with reduction of intracellular GSH leading to cancer growth arrest. Controlling the extracellular redox state through modulation of CyS/CySS concentrations and CyS/CySS transport could be an important determinant in the development of new cancer therapeutics.
TRXSH1
Extracellular TRXSH1 expression has been established as a marker in several cancer tissues. Drugs identified as TRXSH/TRXS-S inhibitors, including auranofin and nitrosoureas, have already been used in the clinic (7, 102). However, the efficacy of these inhibitors has not been established because of their lack of specificity and cytotoxicity. In contrast, TRXSH has shown antiinflammatory effects in the circulation; thus, the use of recombinant human TRXSH1 (rhTRXSH1) in inflammatory-related cancers has been proposed. Although elevation of TRXSH1 in the extracellular space/microenvironment is associated with cancer progression, it was demonstrated that administration of rhTRXSH1 had no effect on tumor growth in a cancer model (115). This discrepancy may be explained by the fact that TRXSH1 may become an oxidized form, TRXS-S, which then has no mitogenic effect. Thus, inhibition of TRXSH1 may potentially attenuate cancer growth, whereas rhTRXSH1 has potential in the treatment of inflammatory-related cancer.
H2O2
For redox-based therapy to be successful, it is important to use the appropriate ROS and ROS-generating system. H2O2 is a possible candidate for redox-based therapy because of its relative stability and mild cytotoxicity. Many antitumor agents exhibit antitumor activity through H2O2-dependent activation of apoptotic cell death (104); thus, use of an extracellular H2O2-generating system has been proposed as an alternative approach for development of H2O2-dependent antitumor therapy. H2O2 used as a single agent in the extracellular space may not be effective because of decomposition of H2O2 by GPX3, TRX1, or other antioxidant-related proteins in the extracellular space/microenvironment. Administration of a H2O2-generating system in combination with GPX3 or TRXSH1 inhibitor compounds may improve antitumor efficacy.
NO•
In cancer cells, NO• production may increase blood flow, resulting in increased oxygen delivery and increased angiogenesis; thus, treatment of cancer by using NO• inhibitors has been proposed. Conversely, NO• production in cancer cells may inhibit tumor cell growth and invasion through inhibition of MMP activity (16). Treatment of cancer cells with NO• donor compounds, such as SNAP, have been tested. Our laboratory demonstrated that the invasive ability of highly aggressive WPE1-NB 26 prostate cancer cells was decreased after treatment with the NO• donor, SNAP (Fig. 6D) (16). However, administration of NO•-donor compounds to animals causes a prompt decrease in blood pressure, which becomes problematic for in vivo studies. It is not clear at present whether increasing or decreasing NO• levels would be more beneficial to cancer patients; further studies should be performed.
Approach to Redox Compartment Therapy in Cancer
Over the past years, several researchers have put great effort into studying the relations between redox imbalance and cancer. Recently, redox-based therapies have been proposed for treatment of cancer, with the aim of targeting the redox-related proteins that are involved in the carcinogenesis process. The optimal targets are those present or altered only in tumor cells. Multiple redox-related proteins are involved in the process of carcinogenesis, proliferation, and metastasis.
Although most attention has been focused on the intracellular redox state as a target for cancer therapy, increasing evidence suggests that extracellular redox-related proteins may be potential therapeutic targets for cancer treatment. Extracellular redox-related proteins as cancer drug targets is a relatively new concept, but one that may be successful with further development. Although some studies described herein analyzed the role of extracellular redox state in in vitro cultured cells, direct in vivo evidence has not been obtained. The lack of in vivo evidence is problematic, considering the complexity of in vivo tumor progression, involving not only oncogenes and tumor-suppressor genes but also stromal cells constituting the tumor microenvironment. More firmly to establish the involvement of the extracellular redox state in initiation and promotion of tumors, animal-model experiments such as knockout and knockin mice are critical, in which the expression and activity of extracellular redox-related proteins are genetically altered. Possible effects of modulation of intracellular and extracellular redox states during cancer progression are proposed in Fig. 9.
In addition to intracellular versus extracellular redox therapies, each compartment in individual cell types has distinct redox characteristics; thus, a need exists to obtain and integrate knowledge of redox compartmentalization into a broader context in the development of cancer therapeutics. Current evidence shows that the concept of a single redox balance within biologic systems is inadequate, because each compartment of each different cell type has unique redox characteristics. The redox characteristics of subcellular compartments have evolved in conjunction with the functions of each organelle. Thermodynamic disequilibrium of redox couples, antioxidant proteins, and ROS/RNS levels within and between compartments is considered to be determinant factors of redox balance in each compartment. Redox communication between compartments is poorly understood and represents an important subject for further investigation. Systemic treatment of cancer by using nonselective redox-related low-molecular-weight compounds may not be effective, an example being treatment of cancer cells with antioxidant compounds that are not site specific. Modulation of compartment-specific redox-related proteins such as MnSOD (mitochondria) or EC-SOD (extracellular space) has demonstrated better tumor-suppression properties than systemic SOD-mimic compounds.
A differential response of normal versus tumor cells to redox therapy has been proposed because of their different intrinsic redox states. We propose that each type of cancer and each step of cancer progression has a unique redox profile. Thus, it would be important to establish the redox state of each subcellular compartment in each cancer-cell type and at each step of cancer progression to allow the design of appropriate redox therapies. Studies have demonstrated that more than one redox-related protein in different compartments is altered during cancer progression, an example being prostate cancer cells, which demonstrated increased GPX3 (extracellular space) expression in PIN but decrease of GPX3 expression in neoplasia; these results were in contrast to GPX2 (mitochondria), which was downregulated in both PIN and neoplasia (85). PIN might then be prevented with an anticancer drug that inhibits extracellular GPX3 or induces mitochondrial GPX2 activity, whereas neoplasia might be treated with an anticancer drug that induces extracellular GPX3 or mitochondrial GPX2 activities or both. Nevertheless, alteration of extracellular redox-related proteins may not result in change of overall extracellular redox status; instead, it may result in disruption of redox signaling or cascade downstream signaling control, which may result in alteration of cancer cell behavior. Thus, using antioxidant supplements solely as anticancer therapy may not be sufficient; combination therapy with agents that modulate redox-signaling proteins may be more beneficial (39).
Much interest surrounds redox-compartment therapies in cancer and, in particular, in their potential to enhance further the efficacy of current treatments. However, considerable thought is needed to maximize their usefulness. Preclinical models are very important for better understanding of the benefit and utility of novel redox-based therapeutics.
Footnotes
Acknowledgments
This material is based on work supported by the Office of Research and Development, Biomedical Laboratory Research and Development Service, and Department of Veterans Affairs (TDO).