
Abstract
In mitochondria, two mono heme c-type cytochromes are essential electron shuttles of the respiratory chain. They are characterized by the covalent attachment of their heme C to a CXXCH motif in the apoproteins. This post-translational modification occurs in the intermembrane space compartment. Dedicated assembly pathways have evolved to achieve this chemical reaction that requires a strict reducing environment. In mitochondria, two unrelated machineries operate, the rather simple System III in yeast and animals and System I in plants and some protozoans. System I is also found in bacteria and shares some common features with System II that operates in bacteria and plastids. This review aims at presenting how different systems control the chemical requirements for the heme ligation in the compartments where cytochrome c maturation takes place. A special emphasis will be given on the redox processes that are required for the heme attachment reaction onto apocytochromes c. Antioxid. Redox Signal. 13, 1385–1401.
Introduction
Characteristics of c-type cytochromes
Cytochromes are redox active hemoproteins. Their activity is based on the change of their heme-iron valence. Mitochondria and plastids possess different types of cytochromes characterized by their heme prosthetic groups. While hemes A, B, and C are found in mitochondria, plastids only contain hemes B and C.
Mitochondrial cytochromes
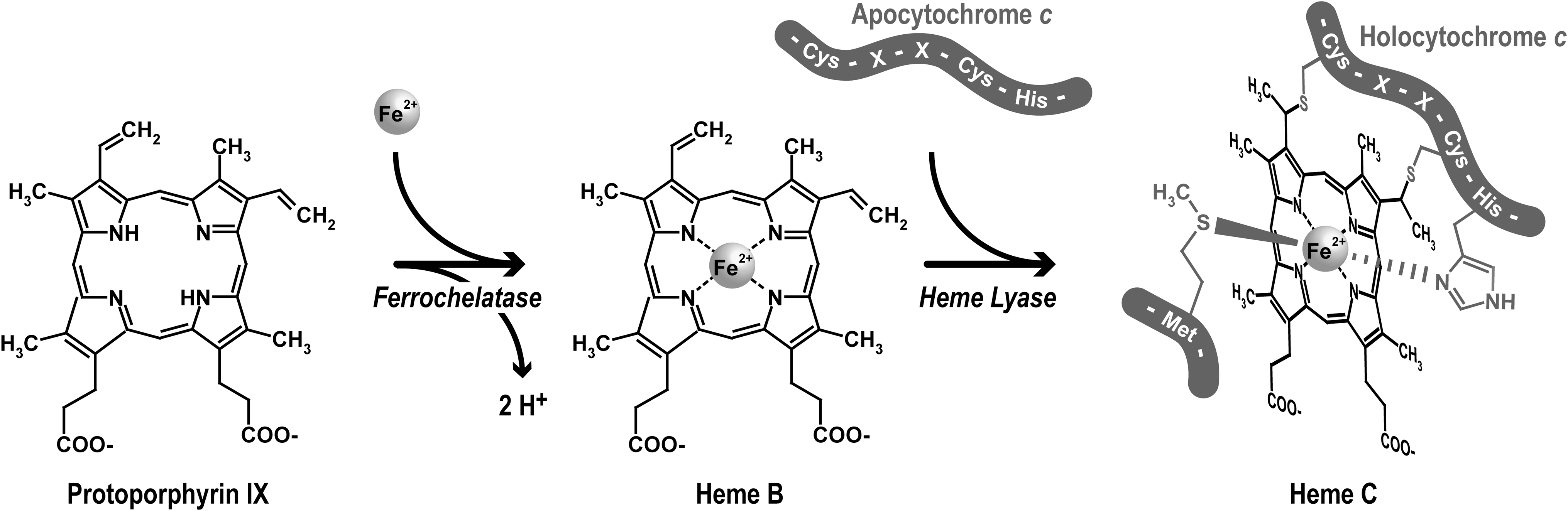
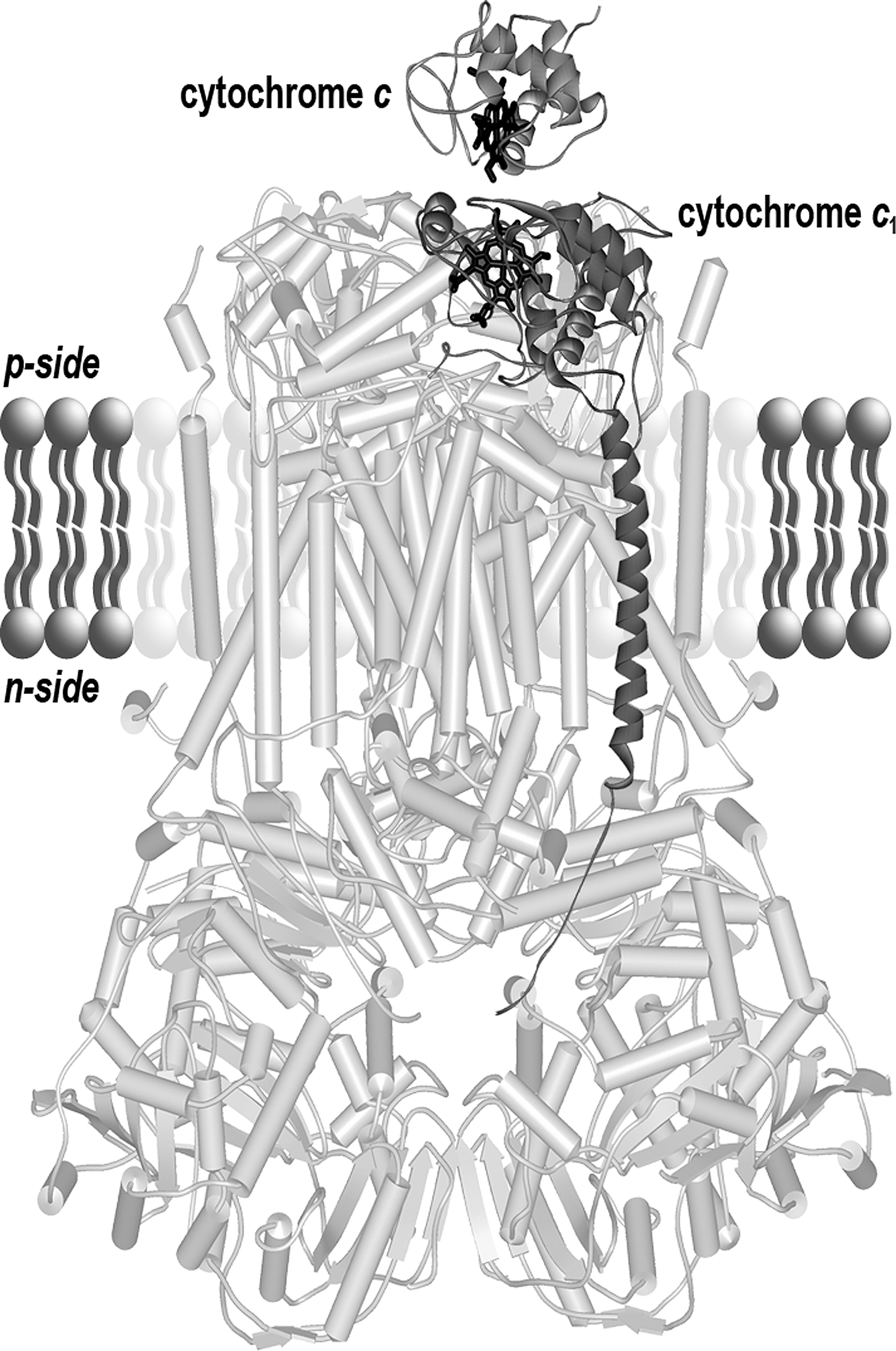
The mitochondrial complex IV (cytochrome c oxidase) contains two A-type hemes (in cytochrome a and cytochrome a 3). In complex III (ubiquinol: cytochrome c oxidoreductase or bc 1 complex), electrons flow from ubiquinol following two paths, via the Rieske iron–sulfur protein, cytochrome c 1 and to cytochrome c, and via two b-type cytochromes to ubiquinone. Cytochromes b and cytochromes c possess an iron protoporphyrin IX or protoheme as a prosthetic group. Iron protoporphyrin IX is the direct product of the ferrochelatase (Fig. 1). In contrast, heme A is a protoheme modified by addition of a hydroxyethylfarnesyl chain and a formyl group. Aside from the heme structure, another criterion for cytochrome classification is the type of ligand involved in the iron-atom coordination. The central iron atom is coordinated by the four nitrogen atoms of the tetrapyrrole ring and by one or two so-called axial ligands that participate in a noncovalent bond between the heme and the protein. In the case of c-type cytochromes, in addition to the axial ligands, heme is covalently bound to the apoprotein (Fig. 1). In most organisms, two thioether bonds stereospecifically connect the cysteines of a CXXCH motif in the apocytochrome to the vinyl lateral chains of the porphyrin rings A and B. In euglenozoans, the XXXCH heme binding motif of cytochromes c provides a unique covalent bond for the heme (4, 39). For mitochondrial c-type cytochromes, the histidine of the heme binding motif provides the proximal axial ligand. The sixth axial ligand is the sulfur of a methionine residue. Whereas hemes A and hemes B are embedded within the respiratory chain complexes, cytochrome c and cytochrome c 1 expose their heme to the mitochondrial IMS (Fig. 2). Cytochrome c is a soluble protein of the IMS. Cytochrome c 1 is anchored in the inner membrane via a C-terminal alpha helix. The N-terminal domain carrying the prosthetic group is located in the IMS.
Plastid cytochromes
In photosynthetic eukaryotes, c-type cytochromes are also found in the chloroplast with the heme containing domain facing the lumen side of the thylakoid membrane. Cytochrome c 6 is a soluble protein in the thylakoid lumen, which shuttles electrons from cytochrome b 6 f complex (analogous to the cytochrome bc 1 complex) to photosystem I. Cytochrome c 6 is only found in green algae and cyanobacteria (55). Cytochrome f, the functional equivalent of mitochondrial cytochrome c 1 is found in all photosynthetic eukaryotes. Recently, a novel c-type cytochrome with an unknown function, cytochrome c 6A was recognized in the green lineage (reviewed in (104)). Interestingly, an extra covalently bound heme, heme Ci, has been characterized on cytochrome b 6, a b-type cytochrome that is the counterpart of mitochondrial cytochrome b in the bc 1 complex (60, 94). Unlike the classical c-type cytochromes described above, this atypical heme is attached by a unique thioether bond to a cysteine and has no axial amino acid ligand. Furthermore, the heme is located near the Qi quinone-binding site (i.e., on the stromal face that is the n-side of the membrane). Hence, cytochrome b 6 is a tri-heme cytochrome with two B-type hemes imbedded in the membrane and the Ci-type heme facing the stroma.
Eukaryotic c-type cytochromes do share a number of features: they are mono-heme proteins and located on the p-side of the energy transducing membrane where they function as electron carriers. In addition, both the apocytochrome and the heme are synthesized in a different compartment and need to be transported to the site of assembly.
Three different pathways for c-type cytochrome biogenesis in mitochondria.
The covalent linkage of heme to apocytochrome requires dedicated machineries. Up to now, four different biogenesis pathways (I, II, III, IV) have been identified and the existence of another one (V) has been inferred from analysis of predicted proteomes (45, 57). These systems operate in prokaryotes, and eukaryotic organelles (Table 1). Their distribution do not follow the evolutionary tree of life and lateral transfer of complete biogenesis pathways was proposed (6, 15, 43).
The proteins involved in System I to System IV biogenesis pathways and sites of assembly are indicated.
Rhodobacter and Arabidopsis CcmH correspond to the N-terminal domain of E. coli CcmH. Rhodobacter Ccml is equivalent to the C-terminal domain of E. coli CcmH.
CcmG corresponds to DsbE in some organisms.
CcsA and ResC are orthologs.
CcsB and ResB are orthologs of Ccs1.
HCF 164, ResA and CcsX are orthologs.
Ccm, cytochrome c maturation; Ccs, cytochrome c synthesis; Res, respiration; CcdA, cytochrome c defective; Dsb, disulfide bond; CCB, cofactor binding, cytochrome b 6 f complex, and subunit petB; CCHL/CC1HL, cytochrome c/C1 heme lyase; Cyc2, locus 2 controlling cytochrome c assembly; HCCS, holo cytochrome c synthase; p and n, the positive and negative side of the energy transducing membrane, respectively; n.i., not identified; -, absent; /, not relevant.
System I
Mitochondria from plants and some primitive protozoans follow System I that is inherited from their α-proteobacteria ancestor, while in most other eukaryotes cytochrome c and c
1 are assembled through System III (41). System I operates in α-, most γ-proteobacteria, and archea. In Escherichia coli, a model organism that has been used extensively for the study of System I, it is composed of 8 Ccm (
System II
Other bacteria (β- and ɛ- proteobacteria, Gram positive, cyanobacteria) use this other multicomponent pathway also referred to as the Ccs pathway (58). At least four Ccs (
System III
This system defined by the prototypical heme lyase, is restricted to the mitochondria of fungi, some protozoans, animals, and green algae (6, 13). The evolutionary history of this pathway is unclear as there is no obvious bacterial ortholog of the mitochondrial heme lyase (6).
System IV
The System IV is specific to the assembly of heme Ci in cytochrome b 6 in both prokaryotes and photosynthetic eukaryotes (60, 84). Contrary to the other biogenesis pathways, the heme assembly occurs on the n-side of the membrane, therefore it will not be further described in this review.
System V
This additional pathway has been inferred to account for the biogenesis of single cysteine containing cytochromes c and c 1 found in species of the Euglenozoa phylum (such as Trypanosoma brucei and Euglena gracilis) where no ortholog of any Systems I, II, III, and IV components could be identified from the predicted proteomes (4). For Systems I, II, and III, the assembly is known to occur on the p-side of the membrane. It is likely that this is also the case for System V.
Our current knowledge of the different maturation pathways came from genetic and biochemical studies on model organisms. The experimental systems cited in this review are listed in Table 1.
Features of c-type cytochrome biogenesis proteins
Based on our understanding of the biochemical requirements for apo to holocytochrome c conversion, it is expected that the cytochrome c biogenesis proteins fulfill at least one of the following tasks: transport of heme across a membrane, delivery of reduced heme to the site of assembly, maintenance of apocytochrome cysteines in a reduced form, and catalysis of the heme ligation. Therefore, conserved motifs or amino acids speaking to one of this above mentioned functions were identified and their functional importance analyzed via site-directed mutagenesis. A common characteristic of the biogenesis proteins is their attachment to the energy-transducing membrane. Indeed, the majority of these proteins possess transmembrane (TM) domains (from 1 to 13). For several of them, the topological arrangement has been experimentally deduced and conserved domains/motifs could be positioned with respect to the membrane (Fig. 3).
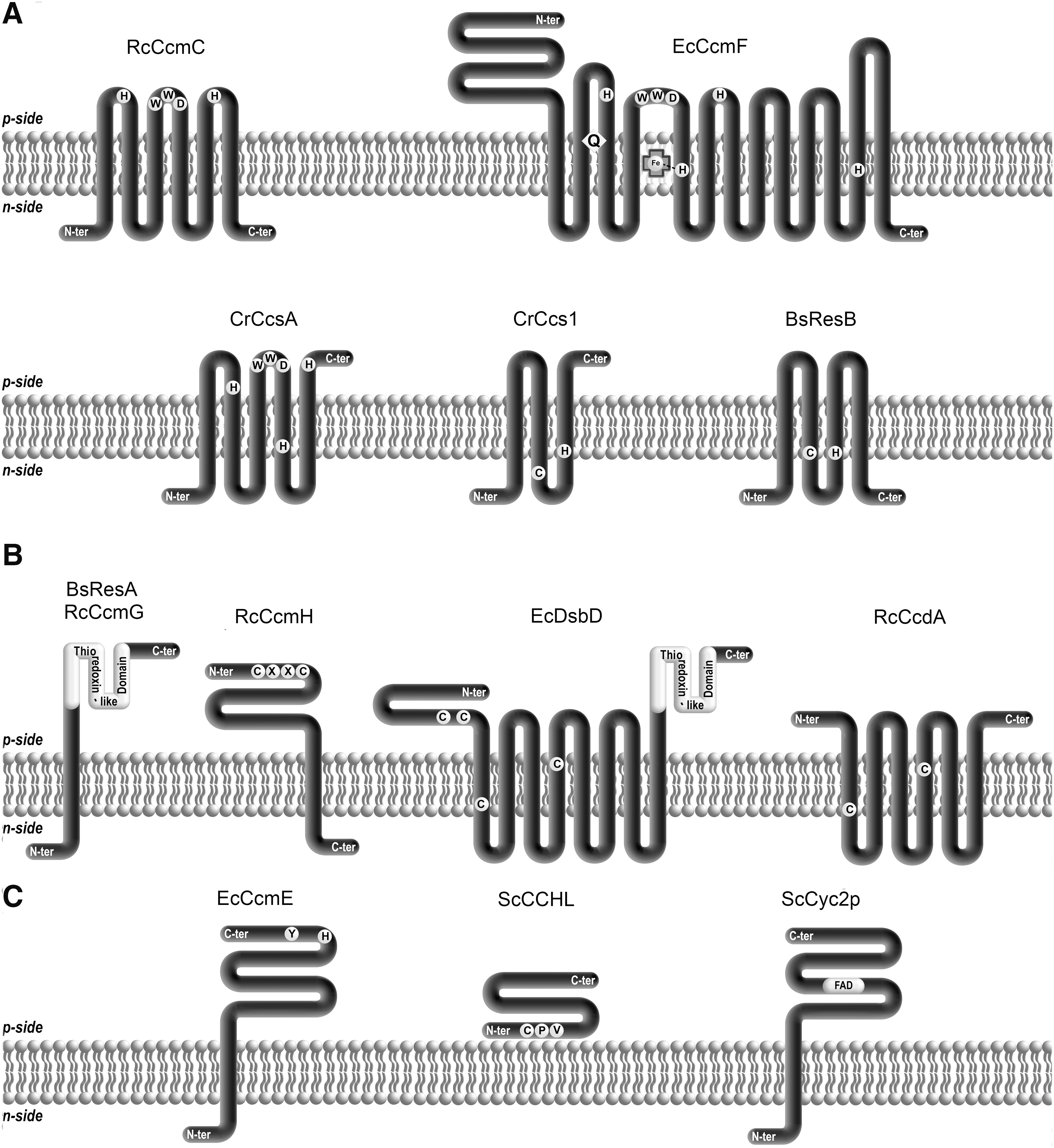
Important motifs and residues indicative of heme handling/delivery including hydrophobic residues and potential heme axial ligands have been identified and localized on both sides of the membrane. Some of these features appear to be common to System I and System II components (Fig. 3A) (59, 98). The tryptophan-rich or so-called “WWD” motif is found in two System I proteins (CcmC and CcmF) and one System II protein (CcsA). This motif is located between two transmembrane domains in a loop oriented toward the p-side of the membrane (11, 26, 42, 45, 78). The CcsBA protein from Helicobacter hepaticus, a natural fusion between CcsB and CcsA, contains histidines located within transmembrane domains close to the n-side of the membrane (38). These histidines are conserved in the corresponding ResB and ResC proteins from Bacillus subtilis. In addition, a cysteine was identified as a heme binding residue in ResB (1). In CcmF, a conserved histidine was shown to be the axial ligand of a B-type heme on the n-side of the membrane (78). The role of the WWD domain and the two pairs of conserved histidines in heme transport and delivery will be presented below.
As for heme delivery, common motifs indicative of a thioredox activity are found in both System I and II components. Putative thiol–disulfide oxidoreductase motifs and a thioredoxin-like domain have been recognized in protein domains facing the p-side of the membrane (Fig. 3B). Thioredoxin domains are present in CcmG and ResA from System I and II, respectively (20, 34, 61). These proteins are membrane anchored and their active domain is extracytoplasmic. Structures of E. coli, B. japonicum, and M. tuberculosis CcmG (29, 44, 71) and of B. subtilis ResA (19) have revealed a typical thioredoxin-like fold for these components. CcmH exhibits a RCXXC motif, but the protein does not fold like a thioredoxin (2, 25). CcmH is the unique component of System I thioredox pathway that is conserved in plant mitochondria. DsbD and CcdA are polytopic membrane proteins that contain conserved cysteines in their central hydrophobic domain (53, 66).
Some biogenesis proteins are specific of a single pathway. This is the case for the ABC transporter subunits CcmA and CcmB, CcmD and the heme chaperone CcmE from System I (Fig. 3C). CcmE is a unique heme chaperone that binds heme transiently in a covalent fashion on a conserved histidine with axial ligation via a conserved tyrosine (31, 48, 87, 102). CcmC is required to load heme on CcmE, while the activity of the CcmABCD ABC transporter complex is needed to release holoCcmE from CcmC, allowing the transfer of heme for ligation onto the apocytochrome (17, 37). CcmD is involved in the stability of the heme delivery protein complex.
The System III pathway involves unique proteins (CCHLs) that do not share any features with System I or II components. CCHLs are not integral membrane proteins, although they are bound to the mitochondrial inner membrane (13, 28, 69). A heme responsive motif (HRM) defined by CPX (where X is I, V, M, L) is present in CCHLs (Fig. 3C). In other proteins, the cysteine in HRM was shown to be necessary for heme binding (106) but it does not appear to be strictly essential for CCHL function in cytochromes c biogenesis (unpublished studies). In yeast, an additional protein Cyc2p is associated to CCHL for holocytochrome c formation. Cyc2p is anchored to the inner membrane and contains a FAD molecule in its C-terminal domain located in the IMS (14). Cyc2p is proposed to play a role in heme or apocytochrome reduction. Cyc2p seems to be restricted to mitochondria of fungi where two heme lyases CCHL and CC1HL operate. There is no structural homolog of Cyc2p in multicellular eukaryotes where HCCS that diplays specificity towards both apocytochrome c and c 1 is the only heme lyase (13, 14).
Requirements for the covalent linkage of heme to apocytochrome c
The covalent attachment of heme to the apocytochrome occurs through two thioether bonds formed between the heme vinyl groups and the thiols of the CXXCH binding motif. This reaction is stereospecific (i.e., the α2 and α4 carbon are linked with the sulfur of the first and second cysteines of the motif, respectively (Fig. 1). In vivo this post-translational modification requires dedicated machineries, however uncatalyzed holocytochrome c synthesis has been obtained and the analysis of the conditions required for correct stereochemical attachment has provided important information.
Holocytochrome c 552 of the thermophilic bacteria Hydrogenobacter thermophilus can be produced in the cytoplasm of E. coli (52, 85). Due to this ectopic expression, apocytochrome c 552 is not accessible to the action of the periplasmic System I and the correct heme attachment occurs independently of this pathway in the cytoplasm. It was proposed that holocytochrome c 552 is assembled spontaneously, due to the partial folding of the apoform of the molecule, in contrast to the unfolded state of mitochondrial apocytochrome c (103). Indeed, when other apocytochromes c are expressed in E. coli cytoplasm, the heme attachment does not take place or is nonstereospecific (54, 85). Nevertheless, in vitro formation of a c-type cytochrome has been achieved with several cytochromes c, including cytochrome c 552 of H. thermophilus (21), but also with Paracoccus denitrificans cytochrome c 550 and the horse mitochondrial cytochrome c (22). The in vitro reaction occurs in two steps. First, a b-type cytochrome is formed involving coordination of heme iron with amino acids in the protein. Next, the thioether bonds are spontaneously formed, provided that the heme in the b-type cytochrome intermediate is in the correct orientation. The divalent iron heme and the free thiol groups are required to avoid the formation of incorrect products and acidic pH appears to be needed to promote the reaction (21). If exogenous reductant is omitted from the in vitro reaction, intramolecular disulfide between the cysteines of the CXXCH motif is detected and holocytochrome c formation does not take place.
Delivery of Reduced Apocytochrome c, the Competent Substrate for the Heme Ligation Reaction
As presented above, the reduction of apocytochrome c is a biochemical requirement for the heme ligation reaction. In this section, we will review the experimental evidences that support the in vivo operation of a pathway that maintains apocytochrome c sulfhydryls under the reduced form.
Evidence for the implication of thiol–disulfide chemistry in bacterial System I and II maturation pathways
Rational for thioredox chemistry in the context of cytochrome c assembly
In the bacterial periplasm, thiol–disulfide homeostasis is under the control of several redox enzymes that promote disulfide bond formation and the isomerization of proteins with incorrect disulfide bonds (50, 51, 56, 66, 67). The thiol-oxidizing pathway is defined by the Dsb system that consists in thiol oxidase DsbA and recycling enzyme DsbB.* DsbA catalyzes disulfide bridge formation on cysteine-containing substrates that are translocated across the plasma membrane into the periplasm. DsbB operates by recycling reduced DsbA to its oxidized form. The electrons derived from DsbA reoxidation are shuttled to the quinone/menaquinone pool via the electron transport chain (Fig. 4). Several hundred proteins are substrates of the Dsb system, but genetic inactivation of this system is surprisingly not lethal but yields redox disturbance in the periplasmic space (50, 51, 56, 66, 67). Because apocytochromes c contain cysteines at their heme binding sites, they are believed to be substrates of the Dsb machinery and need to be consequently reduced to provide free sulfhydryls for the heme ligation.
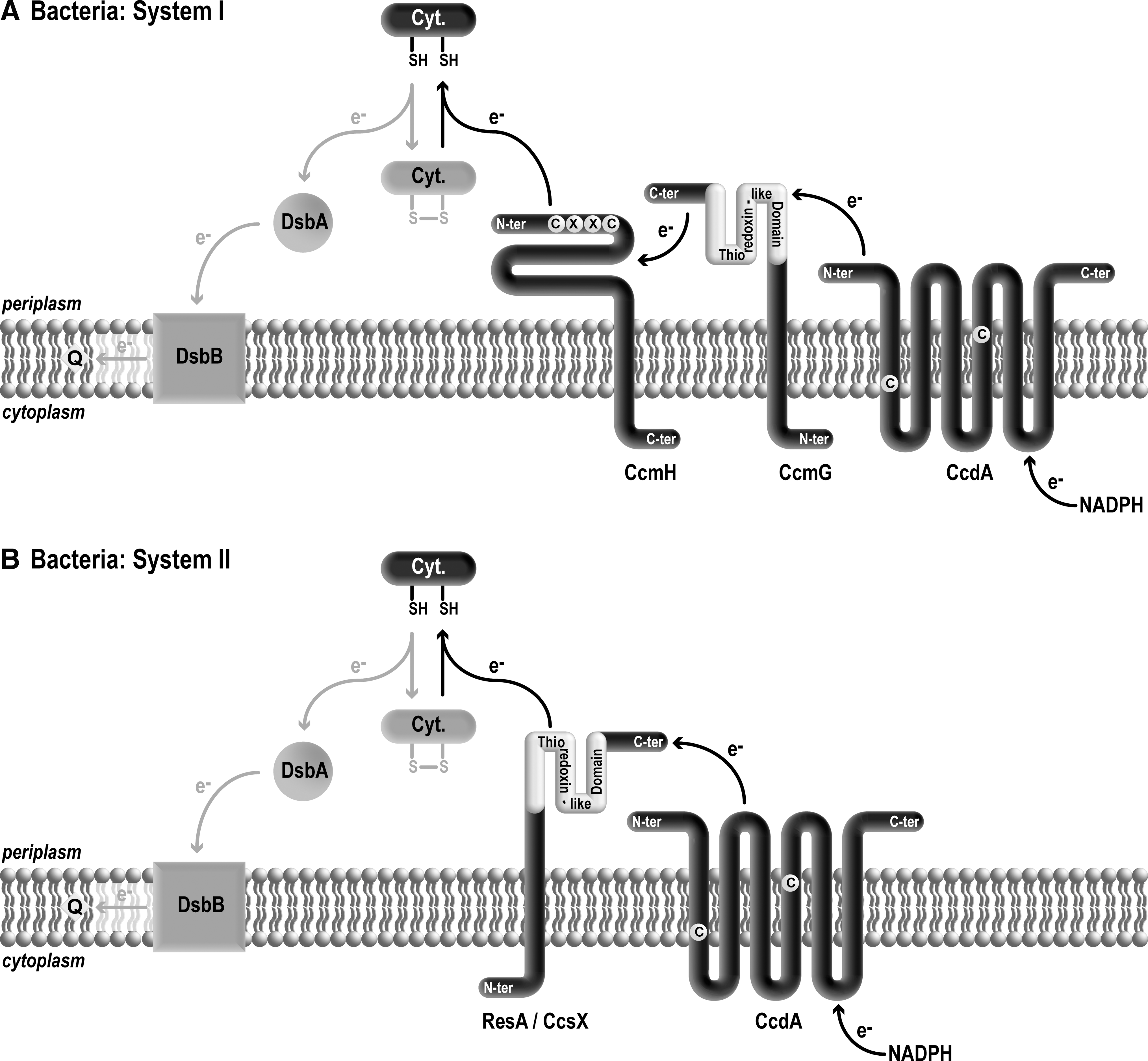
The formation of a disulfide bond at the CXXCH motif was believed initially to be a necessary step in the formation of holocytochrome c (reviewed in (45)). The proposed model was that apocytochrome c needs to be oxidized in order to be consequently reduced and reacted upon with heme. However, there is now multiple experimental evidence indicating that the disulfide bond forming catalysts are not essential for cytochrome c maturation (7, 24, 32, 36, 58). The fact that dsbAB mutants accumulate reduced levels of holocytochrome c suggests that the disulfide bond forming machinery is nevertheless required for optimal production of cytochromes c (101).
The reduced state of apocytochrome c is an absolute requirement for the heme ligation reaction. Consistent with this view is the finding that cytochrome c maturation requires the operation of several thiol–disulfide oxidoreductases that define a transmembrane subpathway for the delivery of reducing equivalents from the cytosol to the CXXCH targets in the periplasm (7, 34, 51, 66, 81). The current view is that the oxidized apocytochrome c CXXCH motif is reduced by the activity of thiol–disulfide oxidoreductase, CcmH in System I or thioredoxin-like ResA/CcsX in System II (Fig. 4). The observation that cytochrome c deficiency due to an inactivation of the thio-redox pathway can be compensated by exogenous thiols is the most convincing evidence that the disulfide in the CXXCH motif is the relevant target of action of this pathway (8, 10, 24, 32, 33, 36). The demonstration that recombinant forms of CcmH (65) and ResA (32) are active in vitro in thiol–disulfide exchange reactions solidifies this proposal. However, there is still no direct in vivo evidence that apocytochromes c are reduced by the proposed redox enzymes CcmH or ResA/CcsX. When the thio-reduction pathway does not operate, apocytochromes c are believed to become disulfide bonded by the Dsb enzymes and are unable to be acted upon by heme. In accord with this scenario is the finding that cytochrome c deficiency due to loss of the thio-reducing pathway can be compensated by mutations inactivating the thio-oxidation pathway (24, 33, 101). So it appears that the thioreduction branch in the context of cytochrome c assembly is only required to counteract the activity of the thio-oxidation pathway.
A multicomponent transmembrane thioreduction pathway
In both bacterial Systems I and II, a transmembrane thioredox pathway operates to deliver reducing equivalents to apocytochromes c by sequential thiol–disulfide exchange reactions. A central component of this redox relay is the CcdA/DsbD thiol–disulfide transporter (56, 66, 92). CcdA and DsbD are polytopic membrane proteins with conserved cysteines involved in transducing the reducing power across the membrane (56, 66, 92). While CcdA or DsbD occurs in System I or System II and are functionally interchangeable (53), they are in fact distinct molecules (Fig. 3B). Both proteins display a conserved central domain composed of six transmembrane domains but DsbD contains additional N- and C-terminal domains that expands its redox activity (53). Whereas CcdA is believed to be restricted to cytochrome c biogenesis, DsbD is also involved in reducing protein disulfide isomerases, the thio-reducing branch of disulfide bond formation (56, 66, 92). However, this view might be simplified as the role of CcdA in other processes unrelated to cytochrome c assembly has now become apparent (56, 66).
In System I, two models have been proposed for the reducing pathways: in the first one, the electrons conveyed by CcdA or DsbD are sequentially transferred to thioredoxin-like protein, CcmG, oxidoreductase CcmH, and apocytochrome c (35). In the second model, CcmH and the apocytochrome form a mixed disulfide that is resolved by CcmG, resulting in a reduced apocytochrome and a mixed disulfide between CcmG and CcmH. This mixed disulfide has been proposed to be reduced by CcdA/DsbD (75). In System II, the electrons transferred via CcdA/DsbD are relayed to thioredoxin-like ResA/CcsX and onto apocytochrome c (Fig. 4).
The detection of mixed disulfide between CcdA and CcmG is a solid proof that CcmG is reduced by the activity of the thiol–disulfide transporter in vivo (53, 93). An electron transfer from CcmG to CcmH and apocytochrome c was deduced from the measurement of the redox midpoint potentials that are an indication of the thermodynamical likelihood of electron transfer reactions (88). Moreover, in vitro experiments showed that recombinant CcmG can reduce CcmH and that an apocytochrome c mimick can be reduced by CcmH (65).
However, there is still no in vivo evidence that CcmG is the reductant of CcmH and that apocytochrome c is reduced by the activity of CcmH. The detection of mixed disulfide between CcdA/DsbD, CcmG, CcmH, and apocytochrome c should indicate which redox enzyme controls the electron transfer from CcdA/DsbD to apocytochrome c and distinguish between the two proposed models of CcmH action. A clue for a distinct role of CcmH came from the finding that unlike ccdA-null or ccmG-null, a ccmH-null mutant cannot be compensated by genetic inactivation of the disulfide bond-forming pathway (101). This suggests that the role of CcmH is not limited to the reduction of oxidized apocytochrome c. This function was first inferred from the fact that ccmH mutants can be compensated by reduced thiols similarly to ccmG or ccdA/dsbD mutants (35, 80, 82). It is now apparent that the heme ligation reaction is mechanistically very different when catalyzed by System I or System II machinery. One possibility is that CcmH is recruited to ensure stereospecificity of the heme attachment reaction by the CcmF heme lyase. Interestingly, in Archaea and some bacteria, the absence of CcmH appears to correlate with a variant of CcmE containing a cysteine instead of a conserved histidine as the heme binding residue (5). The significance of this is obscure but suggests that the redox role of CcmH is also linked to the activity of heme chaperone CcmE.
The involvement of redox chemistry in mitochondrial System I and System III assembly pathways
The question of redox chemistry in the context of mitochondrial cytochrome c assembly received little attention because of the accepted dogma that the IMS was a “naturally” reducing compartment due to the permeability of the outer membrane to cytosolic reducing components such as glutathione and pyridine nucleotide (12). However, the use of redox sensor showed that redox homeostasis in the IMS is independent from that of the cytosol (49). Moreover, the fact that thiol–disulfide chemistry is a catalyzed process has gained appreciation with the discovery that Mia40p and Erv1p drive the oxidative folding of a subset of IMS resident proteins (90). The Mia40 import pathway specifically interacts with small IMS proteins that possess a twin CX3C and CX9C motif but also other cysteine arrangement such as Erv1p (40, 63, 97). Apocytochrome c is not dependent upon Mia40p for its import (at least in yeast mitochondria (16)). However, it is possible that mitochondrial apocytochrome c/c 1 become a target of Mia40 or Erv1 in conditions where they can no longer be assembled (Fig. 5), similarly to bacterial apocytochromes c that are believed to become disulfide bonded when they are not maintained reduced by the redox assembly components (24, 33, 101).
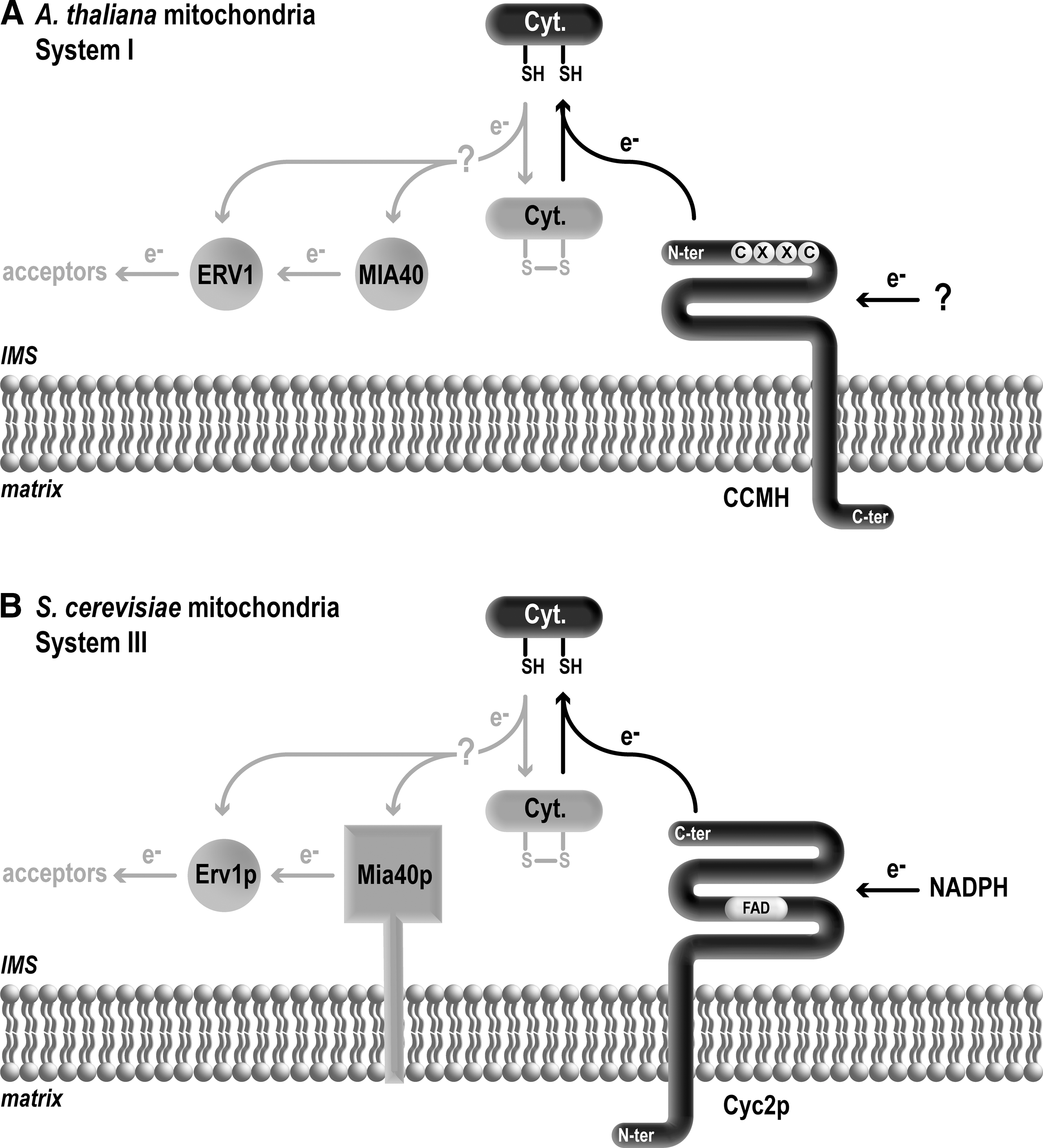
System I
Plant mitochondria do not possess the periplasmic Dsb oxidizing system or the DsbD/CcdA protein, which is key in the transfer of reducing equivalents through the membrane. In addition, no ortholog of the thioredoxin-like gene ccmG could be identified in the plant genome. This observation is striking when considering that most of the CCM proteins involved in heme delivery are conserved in plant mitochondria. CCMH is the only CCM protein present in plant mitochondria, which has been proposed to fulfill a thiol–disulfide oxidoreductase function (Fig. 5A). The topology of Arabidopsis thaliana CCMH (AtCCMH) is similar to that of its bacterial counterpart: AtCCMH is an integral membrane protein and the CXXC motif located in the IMS-facing N-terminal domain (64). In A. thaliana, CCMH is an essential gene required for embryonic development. Despite sequence divergence of the redox active site, VRCTECG motif in plant mitochondria versus LRCXXCQ in bacteria, in vitro experiments revealed that AtCCMH has kept the ability to function as a thiol–disulfide oxidoreductase like its bacterial counterpart (65). AtCCMH can reduce a peptide containing the apocytochrome heme binding motif with an intradisulfide bond (64). Yeast two hybrid experiments revealed that the IMS domain of AtCCMH interacts with apocytochrome c (64). In Arabidopsis, the CcmF ortholog is encoded by three mitochondrial genes ccmFN1
, ccmFN2
, and ccmFC
. Domains of mitochondrial CcmF
In one model, bacterial CcmH is proposed to form a mixed disulfide with apocytochrome, which is resolved by the specialized CcmG thioredoxin-like protein, releasing reduced apocytochrome (75). If such a model is valid, one must postulate the operation of another redox protein because no CcmG ortholog is present in System I mitochondria. In plants, it appears that a large number of redoxins with diverse catalytic and regeneration mechanisms have evolved in order to cope with environmental stresses (83). Some of them may have taken over CcmG specialized function.
System III
With CCHL as the only prototypical component, System III appears unexpectedly simple. The “simplicity” of System III versus the complexity of Systems I and II is puzzling and prompts the following question: If all cytochromes c are united by common biochemical requirements for their assembly, irrespective of the membrane system under consideration, what is the mechanism controlling the maintenance of the apocytochrome c/c
1 (and heme) substrate(s) in a reduced state prior to the heme ligation reaction? Cyc2p, the only redox System III component was first isolated by the Sherman group, but its function had remained obscure since a cyc2-null mutant is not drastically affected for c-type cytochrome assembly (reviewed in (45)). The molecule was re-isolated as a partner of CCHL through a multicopy genetic screen and further characterized (13). Cyc2p contains a FAD co-factor noncovalently bound and is located at the mitochondrial inner membrane with the FAD binding fold exposed to the IMS (14). An indication of the redox activity of Cyc2p in vitro comes from the finding that this protein carries a NADH/NADPH-dependent reductase activity (14). A clue to Cyc2p activity in vivo came from the observation that a cyc2-null mutation results in a synthetic respiratory deficient phenotype with loss of holocytochrome c
1 assembly when combined with a mutation that maps to the apocytochrome c
1 encoding gene (14). The mutation results in a CA
Delivery of Reduced Heme
Heme delivery to cytochrome c assembly pathways requires transport through a membrane
The last step of heme synthesis is catalyzed by the ferrochelatase that inserts Fe2+ into protoporphyrin IX to form protoheme (Fig. 1). In prokaryotes, ferrochelatase is located on the cytoplasmic side of the membrane and heme must be transported across the cytoplasmic membrane to be attached to apocytochrome (70, 100). In eukaryotes, ferrochelatase is found in plastid or in mitochondria. Plastid ferrochelatase is reported to be associated with both envelope and thylakoid membranes, but the majority of enzyme activity is found in thylakoids (95). The ferrochelatase is believed to be located on the stromal side. Hence, there is a need for heme to cross the thylakoid membrane to reach the lumen side of the membrane where assembly through system II takes place. In mitochondria, two different situations are encountered depending whether the organelle compartment is the site of heme synthesis. In most animals and fungi, mitochondria are the only source of heme for the entire cell. The mitochondrial ferrochelatase is located on the matrix side of the inner mitochondrial membrane and heme export to the IMS for c-type cytochrome maturation and to the cytosol for cellular hemoproteins assembly is obligatory (3, 47). In photosynthetic eukaryotes, it is believed that mitochondrial heme originates from plastid (45, 96). Such an extramitochondrial source of heme is also required for cytochrome biogenesis in some organisms that are natural heme auxotrophs such as the nematode Caenorhabditis elegans (72). The delivery of heme originating from outside the mitochondria to the mitochondrial IMS could follow different routes involving at least transport through the outer mitochondrial membrane (45). It should be noted that there is no correlation between the presumed origin of heme and the cytochrome c maturation system used in mitochondria (I or III).
In the context of system III, heme transport across mitochondrial membranes is not understood, while for system I and II, proteins involved in transport, binding and presentation of heme have been identified (30, 37, 42, 98). However, Systems I and II components involved in heme delivery/handling appear to be specific to the cytochrome c maturation pathway. It is possible that in System III, the provision of heme to the IMS is depending upon a general heme exporter that has escaped genetic identification in saturating screens for mutants deficient in mitochondrial cytochromes c (89). Recently, the functions of the Ccm and Ccs proteins in heme provision to the p-side of the membrane have been deciphered and new roles have emerged (59). In System I, a heme iron redox control has been defined and a new function for CcmF protein as a quinol:heme oxido reductase has been unveiled.
System I: the determinant role of the heme chaperone CcmE and the operation of a heme iron redox system
In System I, CcmABCDE proteins are implicated in heme delivery to the heme lyase unit composed of CcmFH. CcmE appears to occupy a key and central position in this heme delivery branch. CcmE is a periplasmic heme chaperone that binds heme covalently on a single histidine residue (87). A covalent adduct is formed between one vinyl of heme and a conserved histidine on CcmE. CcmE first interacts with CcmC to capture heme and with CcmF to transfer it to apocytochrome for covalent ligation (76, 77). In E. coli, overexpression of CcmC is necessary and sufficient to promote covalent heme incorporation into CcmE, while CcmD favors the formation of holo-CcmE (86). CcmC is an integral membrane protein that contains the hydrophobic WWD domain in its second periplasmic loop (Fig. 3). The ability of CcmC to bind heme has been shown for the first time by affinity chromatography on hemin agarose (77). Recent advance from the Kranz lab has uncovered the role of axial ligation in maintaining the oxidation state of heme iron in System I (78). The expression of a subset of Ccm proteins allowed the authors to trap different intermediate complexes with bound heme. Biochemical and spectral analyses of these complexes have led to the following sequence of events (Fig. 6). Heme is transferred through CcmC and can be detected in a CcmCDE complex before the covalent heme attachment onto CcmE. The external heme binding domain of CcmC is composed of the periplasmic WWD domain proposed to provide a hydrophobic platform and two conserved histidines located in the adjacent periplasmic loops 1 and 3, which provide the fifth and sixth axial ligand to heme iron (Fig. 6). When holoCcmE is formed, heme is still coordinated by CcmC histidines. The change in axial ligand only occurs when holoCcmE is released from CcmC by the action of the ATPase activity of CcmAB (17, 37). In free holoCcmE, heme is ligated by CcmE Tyr as already proposed from structural and spectroscopic data (31, 102). The work by Richard–Fogal et al. revealed that, although heme is transferred to CcmC periplasmic heme binding site in a reduced form, all heme detected on holoCcmE or holoCcmE containing complexes is oxidized (78). This observation implies that the covalent ligation of heme to the His130 of CcmE is somehow accompanied by heme oxidation, yet the oxidant for ferric heme formation is not identified. In accord with this finding is the fact that apo-CcmE has a higher affinity to ferric heme when covalent attachment of heme to CcmE is achieved in vitro (23). The importance of the iron state for the chemistry of heme adduct on CcmE has been reviewed in (59).
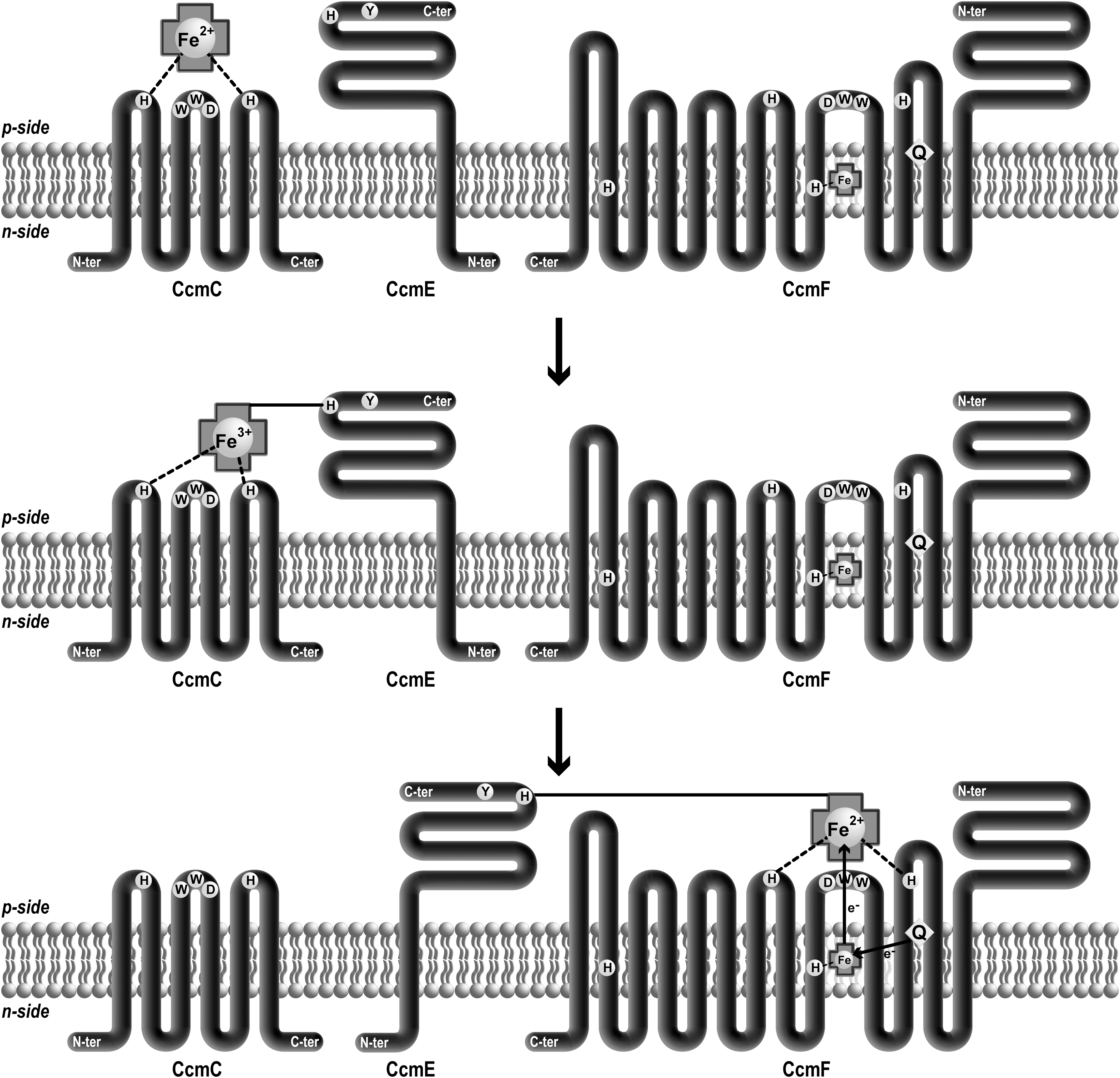
As described in the first section, reduced heme is a prerequisite for the heme ligation reaction. Therefore, a holoCcmE reductase function is needed to complete holocytochrome c formation. Similarly to CcmC, CcmF possesses a periplasmic heme-binding site that provides axial ligands to holoCcmE heme. CcmF also contains a B heme ligated by at least one histidine located in TM5 and possibly a second one in TM12 (78) (Fig. 6). The structural heme of CcmF is reduced in vitro by quinol. This led to the proposal that CcmF acts as a quinol:heme oxidoreductase that relays electrons from quinol to holoCcmE heme through the B heme. However, there is still no direct proof that the B heme in CcmF reduces ferriheme prior to its ligation onto apocytochrome c.
Most of the components of the heme delivery pathway (i.e., CcmABCEF) have counterparts in plant mitochondria (41). The most hydrophobic proteins (CcmB, -C, -FN1, -FN2, -FC) are still encoded by the mitochondrial genome while the peripheral CCMA and the monotopic CCME are nuclear encoded (41). The topology and motifs depicted in Figure 3 are conserved in bacterial and mitochondrial proteins (64, 73, 74, 91). Since CcmF is split into three polypeptides, the potential histidine ligands of heme in holoCCME and those of the structural heme are located on two different polypeptides. The three CcmF polypeptides are present in a high molecular weight complex including the CCMH protein (64, 73). Yeast two-hybrid experiments suggest that CcmFN2 interacts with CcmFN1 and CcmFC, while CCMH interacts with CcmFN1 and CcmFN2. Both apocytochromes c and c 1 could also interact with CCMH, CcmFN1 and CcmFN2. The association of the four CCM proteins with CcmE, as a possible heme lyase or scaffold for heme and apocytochrome assembly seems very likely in mitochondria, similarly to the bacterial situation.
System II: Heme channeling via histidine ligands protects iron from oxidation
In bacteria, two out of the four proteins involved in System II, CcsA and CcsB, have been identified as the minimal set required for heme transport and ligation (33, 37) (Table 1). In Helicobacter hepaticus, the two proteins are fused into a single CcsBA protein whose topology has been determined (38). CcsBA has 10 transmembrane domains, the WWD motif flanked by two conserved histidines present in adjacent loops in the periplasm, and two other histidines located in transmembrane segments closer to the cytosolic face. If the overall topologies of B. subtilis ResB (1), M. leprae and C. reinhardtii CcsA (43, 46) differ from H. hepaticus CcsB and CcsA corresponding domains, the location of WWD and histidines that are essential for c-type cytochrome maturation appears to be conserved in all the orthologs (Fig. 3). Frawley and Kranz have recently described a heme transfer pathway in System II using the naturally fused CcsBA protein (38). When expressed in E. coli, CcsBA binds one reduced heme. Mutation of the conserved histidines in CcsBA revealed different roles for the two pairs of residues (Fig. 7). When either of the periplasmic histidines is mutated, CcsBA still binds heme but displays a change in its absorption spectra. This experiment underscores the role of the two periplasmic histidines as axial ligands of heme. Interestingly, heme associated to the CcsBA histidine mutants is found under the oxidized form. This led to the proposal that the axial ligands protect iron against oxidation. An alternative interpretation is that mutation of the periplasmic histidines induces a change in a heme binding interface resulting in heme exposure to air oxidation.
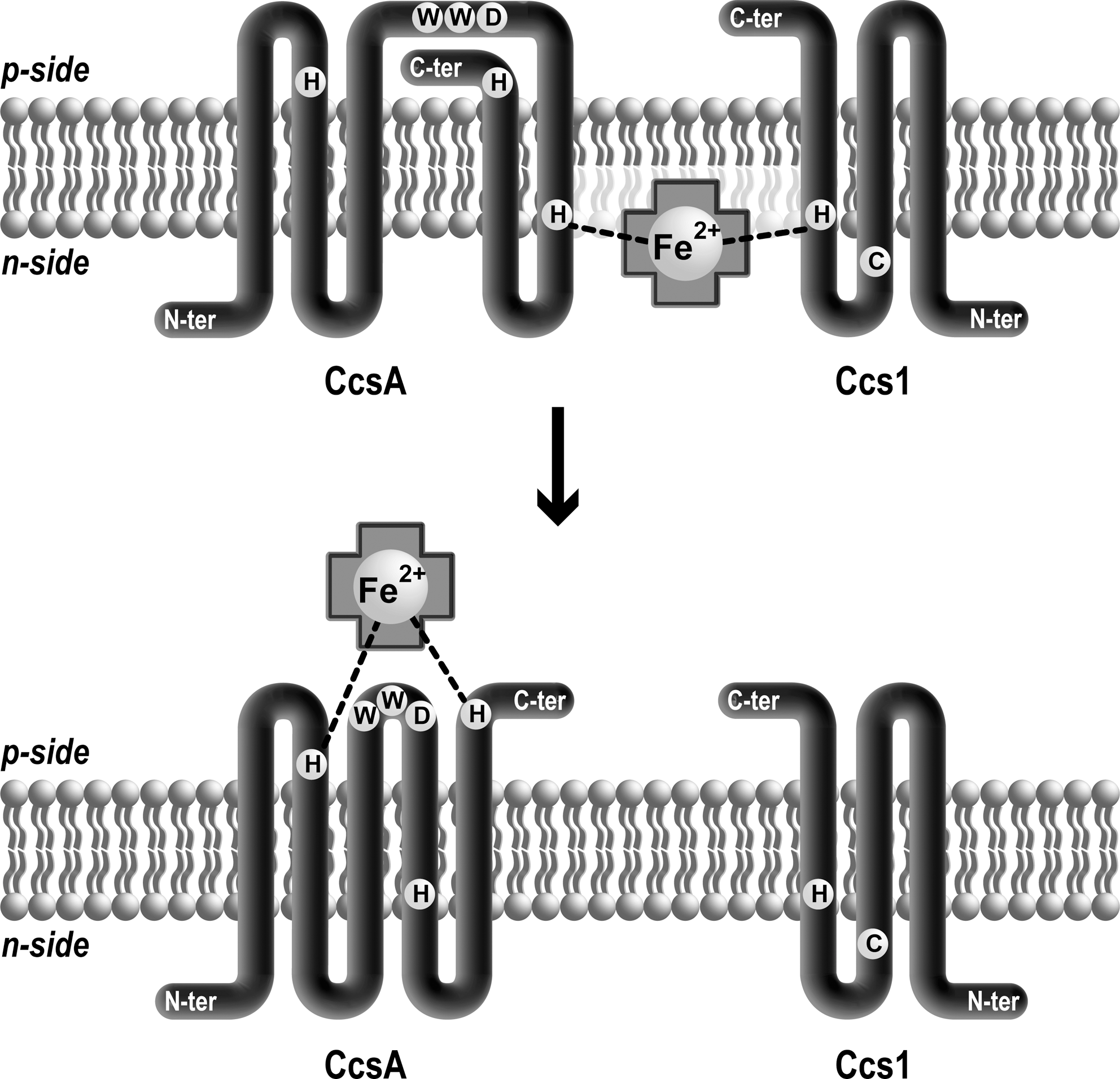
Conversely, when either transmembrane histidine on the cytosolic side is mutated, a reduction of heme binding to CcsBA is observed. In one histidine mutant, the spectral characteristics of CcsBA can be measured and are similar to those of wild type, although some CcsBA bound heme appears to be oxidized. The transmembrane histidines have been proposed to participate in channeling heme to the external heme-binding site and to also play a protective role against heme oxidation (Fig. 7). Interestingly, provision of imidazole restores the function of the TM histidines mutants for cytochrome c production, but this complementation is less efficient under aerobic growth conditions, an observation that was attributed to a possible heme oxidation. A general protective role of histidine axial ligands of heme iron against oxidation has been proposed (38, 62). H. hepaticus CcsBA assures both heme transport and cytochrome c assembly without the help of a heme chaperone like CcmE. The heme attachment reaction in System II appears to be mechanistically simpler than in System I.
This view of a CcsB/CcsA-mediated transport/ligation via the histidine ligands might be oversimplified based on a recent study using B. subtilis ResB and ResC that are orthologous to CcsB and CcsA, respectively. Expression of ResB and ResC in E.coli does not complement cytochrome c maturation, unlike CcsBA (1). Surprisingly, ResB accumulates with a heme covalently attached to a conserved cysteine residue located in the second transmembrane close to the cytosolic side of the membrane (1). This cysteine is not essential for cytochrome c assembly function, as previously shown for Ccs1, the C. reinhardtii ResB ortholog (26). The biological significance of this finding is unclear but it is possible that the cysteine participates in capturing heme on the cytosolic side and becomes reactive to this molecule in conditions where the ResB cannot function as a heme channel.
System III: Role of Cyc2p
The redox role of Cyc2p in cytochrome c maturation is supported by the fact that a recombinant form of the protein is redox active in an in vitro assay (14). However, there is still no direct evidence that heme and/or apocytochrome c sulfhydryls are direct relevant targets of action in vivo and there are arguments in support of both hypotheses (14). The sequence similarity of Cyc2p to NADH-cytochrome b 5 reductase led to the hypothesis that heme, the substrate of the heme ligation reaction could be reduced by the activity of Cyc2p (13, 14). Cytochrome b 5 reductase (EC 1.6.2.2), is a family of ER enzymes that catalyzes the reduction of cytochrome b 5, a b-type cytochrome using reduced pyridine nucleotide as the physiological electron donor. The sequence relationship of Cyc2p to this family of enzymes suggests that a b-type cytochrome could be a target of Cyc2p reducing activity. It is conceivable that a b-type cytochrome is formed as an intermediate of the heme ligation reaction similar to what has been observed in bacterial System I. Another possibility is that CCHL, the mitochondrial heme lyase contains, similarly to CcmF (78), a B-type heme that is reduced by Cyc2p. The presence of conserved histidines in CCHL is compatible with such a scenario but this awaits spectroscopic analysis on recombinant CCHL. That heme is reduced by a flavin-linked NADPH dependent electron transfer is compatible with the observation that both redox molecules are required for the in organello reconstitution of holocytochrome c assembly (69).
Conclusion
The discovery that two distinct c-type cytochromes assembly machineries, System I and System III, operate in mitochondria has raised a number of exciting questions. The realization that covalent linkage of heme to mitochondrial apocytochromes c is achieved by assembly components that differ in number and identity (eight in System I versus three in System III) has challenged our view that the chemistry of the reaction is the same. Studies of bacterial cytochrome c maturation and in organello/in vitro reconstitution of holocytochrome c assembly have established the need of reducing conditions in the heme lyase reaction. That both heme and apocytochrome c sulfhydryls have to be reduced for the heme attachment reaction to proceed in vivo is now no longer questioned. However, the identity of the specific components controlling redox chemistry in the context of mitochondrial cytochrome c maturation is still not established.
Reduction of apocytochrome sulfhydryls
Thioredox chemistry was historically defined, perhaps naively as a sole need to maintain apocytochrome c under the reduced dithiol form prior to the heme ligation reaction. A comparison of thioredox components involved in System I and System II now indicates that this perception needs to be expanded. In System I, a role for CcmH as a redox factor controlling a yet-to-be-defined step in the heme lyase reaction is now beginning to emerge. The exact activity of CcmH is still enigmatic but a “simple” function in converting apocytochrome c CXXCH disulfide to dithiol seems unlikely. Several arguments indicate that the activity of CcmH is dependent upon the heme lyase CcmF and heme chaperone CcmE. It is possible to envision that the CcmE-dependence in relaying heme to the CcmF lyase dictates a specific redox requirement to ensure stereospecificity of the heme attachment reaction. CcmH is likely to have an essential role as it is the only redox component that appears to be conserved in System I mitochondria. There is no transmembrane thio-reducing pathway in mitochondria, and one has to envision that “simple” reduction of the CXXCH cysteines is under the control of unrecognized components, possibly involving Cyc2p in System III. Alternatively, the reduction of CXXCH cysteines might no longer be needed because apocytochrome is rapidly trapped by the assembly machinery and thereby protected from oxidation. Conceivably, CCHL in system III or CcmF/H for System I would exert this protection on the CXXCH cysteines and maintain the apocytochrome substrate in the competent conformation for heme ligation to occur.
Reduction of the heme cofactor
Functional dissection of the heme relay subpathway in bacterial System I and System II has considerably advanced our view on this question. In System I, the involvement of a direct heme reduction mechanism was deduced from the discovery that heme is oxidized when bound to the CcmE chaperone. Because CcmE defines the last step of heme transfer before ligation onto apocytochrome c, a mechanism for heme reduction has to be postulated. There is support but yet no direct proof that heme reduction is achieved by an electron transfer reaction from quinone via the CcmF heme lyase. Since System I mitochondria have kept a number of the heme handling CCM proteins from their bacterial ancestor, it is very likely that the mechanism of heme reduction in System I mitochondria and bacteria is similar. Investigation in the plant experimental system is required to corroborate this view and it is conceivable that some differences might be revealed between System I mitochondria and bacteria. In bacterial System II, it appears that heme is not enzymatically reduced but protected from oxidation, presumably from its site of synthesis (on ferrochelatase) to its site of assembly onto apocytochrome c. It is not known if System III mitochondria require an active mechanism for heme reduction similarly to bacterial System I. The need for a pyridine nucleotide and flavin molecules to reduce heme for holocytochrome c reconstitution in the in organello reaction is indicative of such an active mechanism (68, 69). The discovery that Cyc2p, a NADPH-dependent flavoprotein is required for cytochrome c assembly suggests that this component could function as a heme reductase. However, this proposal still awaits experimental testing as there are also indications that the protein could display disulfide reductase activity. The fact that Cyc2p appears to be restricted to fungal mitochondria raises the question as to the universality of such a mechanism. Other factors must operate in System III mitochondria that have not retained Cyc2p but their molecular identity is currently unknown.
A cytochrome c centric view?
The reason why there is such a diversity in cytochrome c maturation pathways has no obvious and straightforward answer. One attractive hypothesis is that the cytochrome c assembly machineries have evolved with additional functions that remain to be deciphered. In some System I bacteria, there are several reports that the heme handling Ccm proteins participate in cellular processes that are distinct from the cytochrome c maturation (18). These additional functions were postulated on the basis that mutations in the corresponding Ccm genes result in new phenotypes such as disturbance of siderophore metabolism and metal homeostasis that cannot be attributed to loss of cytochrome c production. Considering that the functional repertoire of c-type cytochromes keeps expanding, it is not surprising to envision that their dedicated assembly components could also be recruited for functions beside the well-documented activity in co-factor attachment.
Footnotes
Acknowledgments
This work is supported by a Muscular Dystrophy Association grant (4727), National Science Foundation grant (MCB-0920062) to PPH. VC is supported by an American Heart Association post-doctoral fellowship. GB and EHM are supported by CNRS.
Abbreviations Used
*
By opposition, the negative side (n-side) of the membrane is the mitochondrial matrix, the plastid stroma and the bacterial cytoplasm.