
Abstract
Peroxiredoxin 5 (PRDX5) was the last member to be identified among the six mammalian peroxiredoxins. It is also the unique atypical 2-Cys peroxiredoxin in mammals. Like the other five members, PRDX5 is widely expressed in tissues but differs by its surprisingly large subcellular distribution. In human cells, it has been shown that PRDX5 can be addressed to mitochondria, peroxisomes, the cytosol, and the nucleus. PRDX5 is a peroxidase that can use cytosolic or mitochondrial thioredoxins to reduce alkyl hydroperoxides or peroxynitrite with high rate constants in the 106 to 107 M−1s−1 range, whereas its reaction with hydrogen peroxide is more modest, in the 105 M−1s−1 range. PRDX5 crystal structures confirmed the proposed enzymatic mechanisms based on biochemical data but revealed also some specific unexpected structural features. So far, PRDX5 has been viewed mainly as a cytoprotective antioxidant enzyme acting against endogenous or exogenous peroxide attacks rather than as a redox sensor. Accordingly, overexpression of the enzyme in different subcellular compartments protects cells against death caused by nitro-oxidative stresses, whereas gene silencing makes them more vulnerable. Thus, more than 10 years after its molecular cloning, mammalian PRDX5 appears to be a unique peroxiredoxin exhibiting specific functional and structural features. Antioxid. Redox Signal. 15, 817–829.
Introduction
Peroxiredoxin 5 (PRDX5) (Fig. 1), also named PrxV, ACR1, PMP20, or AOEB166, is the only mammalian atypical 2-Cys peroxiredoxin (41, 80, 99). Partial amino acid sequences of PRDX5 were initially reported as sequences of an unknown liver or lung protein (29, 92). In 1999, three different articles reported the identification and the molecular cloning of a novel mammalian protein named ACR1 (44), PMP20 (99), or AOEB166 (41). Human ACR1 (Alu complex in gel retardation assays 1) was described as a DNA-binding protein potentially implicated in the repression of RNA-polymerase-III–driven transcription of the Alu-family retrotransposons (44). Human and murine PMP20 (peroxisomal membrane protein 20) were characterized as homologues of the previously identified yeast PMP20, a peroxisomal antioxidant enzyme (99). Moreover, human PMP20 was shown to be able to reduce hydrogen peroxide by its thiol-peroxidase activity, and it was proposed that the enzyme may have antioxidant protective functions in peroxisomes but also in the cytosol (99). Finally, human and rat AOEB166 (antioxidant enzyme B166) was identified as a novel mammalian peroxisomal and mitochondrial peroxidase representing the prototype of a new peroxiredoxin subfamily (41). At the beginning of 2000, several articles on PRDX5 were published (52, 80, 103). Among them, the article by Sue Goo Rhee and his collaborators (80) reported a more complete biochemical characterization of human PRDX5, showing that the enzyme is indeed a thioredoxin peroxidase that can be addressed intracellularly to peroxisomes, to the cytosol, and potentially to mitochondria. It was also proposed to classify human PRDX5 among atypical 2-Cys peroxiredoxins, based on its unconventional enzymatic mechanism using two catalytic Cys (80).

According to its amino acid sequence, PRDX5 is the most divergent isoform among mammalian peroxiredoxins (53, 72). Mature human PRDX5 (Fig. 2) shares only 28% to 30% sequence identity with human typical 2-Cys and 1-Cys peroxiredoxins (53). Interestingly, PRDX5 exhibits also a remarkably wide subcellular distribution compared with the other mammalian peroxiredoxins. These features, as well as other characteristics discussed later, suggest that this peroxiredoxin may possess specific functions in mammalian cells compared with the five other isoforms.
Gene Organization, Transcription, and Translation
The human PRDX5 gene is localized to chromosome 11q13 and is composed of six exons and five introns spanning approximately 3.7 kb (41, 45, 46, 61). Interestingly, the number of exons and introns as well as the size of the corresponding protein-coding exons appears to be highly conserved among chordates, although intron lengths varied considerably throughout evolution (61). It must also be pointed out that human PRDX5 and HSPC152/TRM112 genes are in close proximity on chromosome 11 and positioned in opposite directions (61). This locus appears to be conserved in mammals. In humans, only 689 bp separate the two first ATGs corresponding to the translation start codons of each gene, suggesting that PRDX5 and HSPC152/TRM112 may share regulating sequences (61).
In silico analysis of the upstream sequence of the human PRDX5 gene revealed features of the 5′-flanking region of a housekeeping gene. The 5′-flanking region of the human PRDX5 gene and its murine counterpart contain CpG islands, and they lack TATA- and CAAT-boxes (45, 46, 61). Moreover, multiple transcription start sites were reported for the human PRDX5 gene, also a characteristic of housekeeping genes (41, 61, 99). PRDX5 has been reported to be constitutively expressed in virtually all mammalian tissues but at different levels, depending on the cell type (41, 46, 80). It was also shown that transcriptional or translational upregulation may occur in mammalian cells in pathophysiologic situations or in response to different kinds of stresses (see later). Accordingly, the analysis of the 5′-flanking region of human PRDX5 revealed the presence of several putative response elements for transcription factors involved in the response of mammalian cells to oxidative stress, such as Nrf2 or NF-κB (45, 46, 61). However, their implication in PRDX5 regulation is still to be clearly demonstrated. Interestingly, Foxo3a, another transcription factor involved in a direct transcriptional regulation of antioxidant enzymes such as MnSOD/SOD2 or catalase, was also shown to interact with PGC-1α to regulate PRDX5 transcription in endothelial cells (64, 88). Moreover, it was reported that Ets1/2, known as transcription factors implicated in oxidative stress–inducible expression, were associated with HMGB1 to function as coactivators to upregulate PRDX5 expression in human cancer cells exposed to hydrogen peroxide or hypoxia (82). Finally, it was reported recently that a specific compensatory increase of PRDX5 appears in cells of PRDX1-/- mice as the consequence of a fine-tuning mechanism to control increase of cytoplasmic and nuclear ROS (reactive oxygen species) (27). It was suggested that PRDX5 expression may be under the transcriptional control of c-Myc and PRDX1 complexes, therefore regulating ROS homeostasis in situations in which the peroxidase activity of PRDX1 is compromised (27).
Upregulation of PRDX5 expression was first described during acute inflammation induced by lipopolysaccharide (LPS) in rat lung (41). More recently, it was shown that this dramatically increased expression of PRDX5 in the lung during airway inflammation was moderately induced by changes in PRDX5 expression in epithelial cells but rather caused by migrating neutrophils and monocytes recruited by LPS and expressing high levels of this peroxiredoxin (10, 48). Accordingly, PRDX5 expression is highly increased in vitro in primary macrophages exposed to LPS and IFN-γ (1, 19). Downstream of the LPS/TLR pathway, p38 and JNK were shown also to be major contributors to PRDX5 upregulation (1). Moreover, up- or downregulation of PRDX5 has been further reported in the literature in different pathophysiologic situations that may have functional consequences in term of antioxidant protection or redox-dependent signaling. Upregulation was reported in cells with mitochondrial dysfunction (35), in the lungs of mice in an asthmatic model (56), in degenerative human tendons (90), in osteoarthritic cartilage (91), in human tendon cells exposed to H2O2 (101), in ovarian carcinoma (38), in malignant mesothelioma (40), in breast carcinoma (37), in different thyroid pathologies (26, 70), during keratinocyte differentiation (79), in astrocytes in multiple sclerosis (31), in endothelial cells during shear stress (60), and in retina during hyperglycemia (49). Downregulation was described during aging in colon epithelial cells (55), during aging of peripheral nerves (57), in bone marrow macrophages during ionizing radiation exposure (11), in corneal endothelial dystrophy (36), in prostate after androgen deficiency (66, 84), in the nucleus accumbens in the brains of rats addicted to cocaine (18), in corticobasal neurodegeneration (12), in failing myocardium (6), in adrenocortical carcinoma (24), and in rat epithelial cells of airways exposed to cigarette-smoke extract (81).
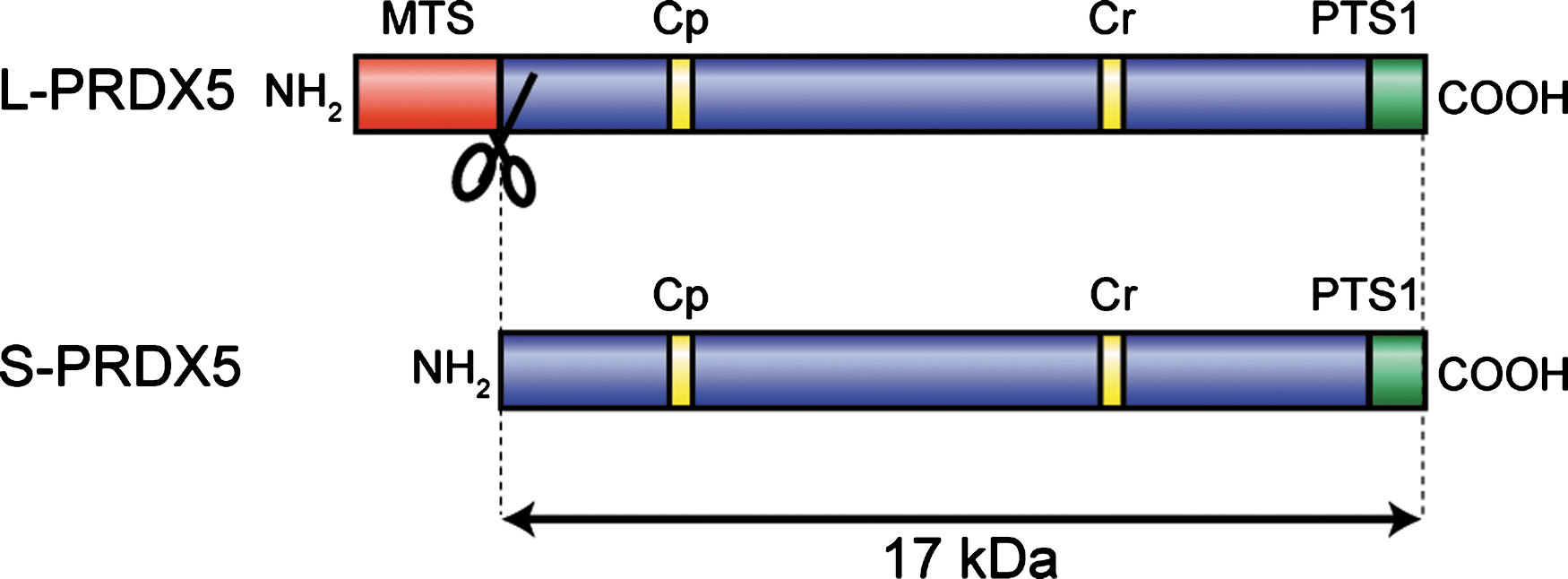
In an initial report on the characterization of PRDX5 (41), it was noted that human PRDX5 mRNAs contained two AUGs in the same reading frame that could be used as two potential translation-initiation sites. It was suggested that translation from the first AUG would result in the synthesis of a long 214-residue protein, whereas the use of the second AUG would result in the production of a short 162-residue polypeptide. Moreover, as reported in Nguyen-Nhu et al. (61), several transcription start sites were also identified in human from PRDX5 transcripts expressed in liver, lung, and brain. These transcription start sites are positioned upstream and downstream of the first translation-initiation site (61), suggesting that the use of alternative transcription start sites or the alternative use of the two translation initiation sites or both could be indeed used by human cells to produce a long form (L-PRDX5) or a short form (S-PRDX5) of the protein (Fig. 2). Interestingly, the additional 52 residues at the amino-terminal part of human L-PRDX5 exhibited features of a cleavable mitochondrial targeting sequence. This sequence was shown to be able to target a fusion of L-PRDX5 and green fluorescent protein into mitochondria (41). Also, it was noted that a potential peroxisomal targeting sequence (SQL) was present at the carboxy-terminus of both S-PRDX5 and L-PRDX5 (41, 99). Further examinations of PRDX5 in tissues or cell lines and subcellular fractionation revealed that the protein was detected at only one molecular size (17 kDa), which was identical to that of the short form (80). The first amino acid downstream of the predicted cleavage site for the mitochondrial presequence of L-PRDX5 corresponds exactly to the first amino acid of S-PRDX5 after the cleavage of its first methionine. It corresponds also to the first amino acid initially determined by amino acid sequencing of AOEB166 spot in two-dimensional electrophoresis analysis of human bronchoalveolar lavage fluid (41, 92). These observations and the identification of a functional mitochondrial presequence in L-PRDX5 suggest that L-PRDX5 is rapidly imported into mitochondria as a precursor protein where the mitochondrial presequence is cleaved, converting L-PRDX5 to a mature form, indistinguishable from S-PRDX5. Accordingly, detection by immunoblotting of PRDX5 in cells transfected with constructs coding for human L-PRDX5 and human S-PRDX5 revealed only one band corresponding to a protein of 17 kDa (4).
Alternative splicing variants expressed in human, baboon, and African green monkey have been also reported (61). These alternative splicing variants were not detected in mouse, rat, or pig tissues (61). Although it was noted that coding sequences are in frame with the two AUGs in the primate mRNA variants, no translation products were so far detected with Western blotting either in human tissues (80, 90, 91) or in human cell lines (80). However, it was shown recently that among these alternative splicing variants in humans, two isoforms showed widespread expression in neoplastic and normal tissues but were unstable at the protein level (78). The peptides were detectable only after inhibition of proteasomal degradation and were shown to be part of the immune-mediated stress response implicated in the detection and elimination of damaged normal or neoplastic cells (78).
Subcellular Localization
PRDX5 is widely expressed in mammalian tissues, and similar to other peroxiredoxins, the amounts of PRDX5 were estimated at between 0.2 and 1.3 μg/mg of soluble proteins in rat tissues (80). However, contrary to the other members of the family, PRDX5 exhibits a surprisingly wide intracellular localization. PRDX5 can be localized to mitochondria, peroxisomes, the cytosol, and the nucleus in mammalian cells. It was previously noted that L-PRDX5 possesses an N-terminal mitochondrial targeting sequence that is able to address a fusion protein of L-PRDX5 and green fluorescent protein to mitochondria (41) (Fig. 2). The mitochondrial localization of PRDX5 was confirmed by subcellular fractionation (15, 46), in situ immunolocalization (3, 4, 40, 54, 63), as well as in mouse mitochondrial proteome analysis (65). The L-PRDX5 also exhibits a C-terminal peroxisomal targeting sequence 1 (PTS1) SQL tripeptide, but it appeared that when both mitochondrial and peroxisomal targeting sequences coexist in the L-PRDX5, the enzyme is addressed preferentially to mitochondria (3, 4). S-PRDX5 possesses only the C-terminal SQL peroxisomal targeting sequence. The peroxisomal localization of PRDX5 was confirmed in peroxisomal fractions prepared from guinea pig liver or HeLa cells (80), by in situ immunodetection, and in peroxisomal fractions of HeLa cells expressing HA-tagged S-PRDX5 (99), as well as in human peroxisomal proteome analysis (75). Moreover, it has been shown that PRDX5 is also localized in the cytosol (4, 40, 80, 99) and, in several situations, in the nucleus of mammalian cells (26, 40, 44, 63, 70). It was proposed that peroxisomal, cytosolic, and nuclear PRDX5 corresponds to the S-PRDX5 form, based on the fact that C-terminal SQL in PRDX5 appears as a weak peroxisomal targeting sequence and that in cells overexpressing S-PRDX5, the enzyme may be localized to these three subcellular compartments (4, 46, 61, 99). It is clear that further work will be necessary to elucidate the mechanisms controlling the complex PRDX5 subcellular addressing.
Crystal Structure
The reduced form
The first available crystal structure of the atypical 2-Cys human PRDX5 concerned its reduced form, in which the side chains of all Cys residues, the peroxidatic (Cp) Cys47, the resolving (Cr) Cys151, and the additional Cys72 were characterized by the presence of a sulfhydryl group (16, 17). The overall structure of the reduced form is presented in Fig. 3 as a topologic diagram and as a ribbon diagram in more or less similar orientations. This structure is characterized by the presence of a thioredoxin fold composed of a four-stranded β-sheet flanked by three α-helices. In addition to this thioredoxin fold, one can observe, in the N-terminal part, a short two-stranded β-sheet followed by a short α-helix. Furthermore, an additional β-strand (β5) is associated with the thioredoxin β-sheet to form a five-stranded β-sheet. Two additional α helices (α3 and α5) are inserted in the sequence. The presence of this last α5 helix is unique, as it is not observed in other peroxiredoxin structures, and it replaces the usual loop joining α4 to β6 in the thioredoxin fold. The complete structure is composed of one single domain. This situation is different from what is observed in 1-Cys and in typical 2-Cys peroxiredoxins, in which a second domain is usually present in the C-terminal part of the sequence.

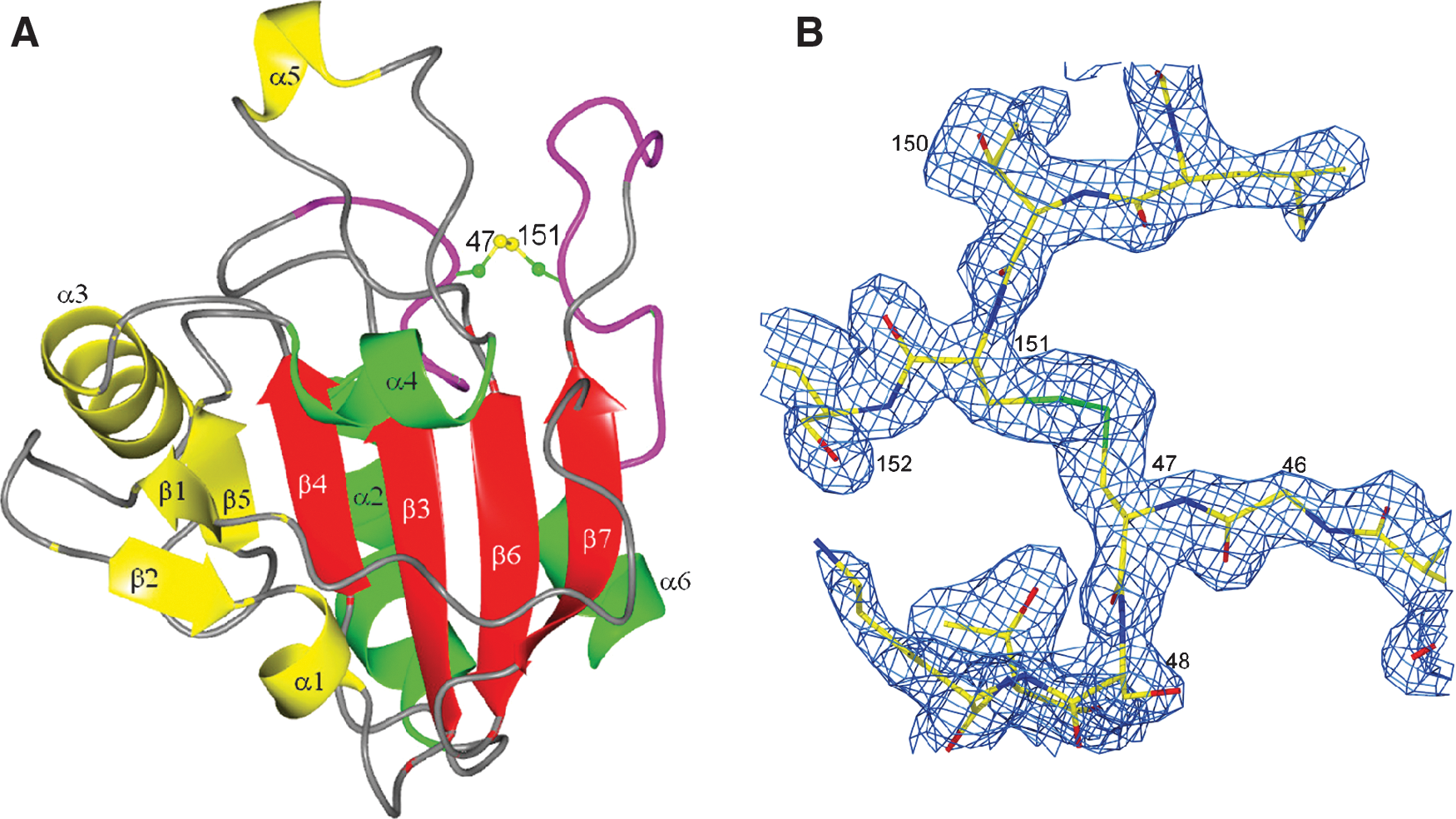
The Cp residue Cys47 of PRDX5 corresponds to a highly conserved Cys residue in all peroxiredoxins. In the reduced form, this residue is located in the N-terminal part of the α2 helix, which is kinked at the level of Ala59. The direct surrounding of the Sγ atom of Cys47 involves Arg127, Thr44, Val39, and a benzoate ion. Arg127 is probably responsible for the positively charged active-site pocket in which Cys47 lies (Fig. 4). As shown in Fig. 5, the Nη1 atom of Arg127 is at a distance of 3.3 Å from Cys47 Sγ. The threonine residue, Thr44, forms a hydrogen bond between its oxygen atom Oγ1 and Cys47 Sγ at a distance of 3.0 Å. The presence of a benzoate ion in the vicinity of the active site was completely unexpected. One of the oxygen atoms is in direct contact with Cys47 Sγ. The aromatic part of the benzoate is in contact with hydrophobic residues belonging to the α5 helix, which is present only in PRDX5. The presence of this ion was observed in all the crystal structures of the reduced forms of human PRDX5, as well as in the C47S mutant of this enzyme (23). It has disappeared in the oxidized forms. The origin and the exact role of this ion remain unknown, but it was speculated by Karplus and Hall (39) that it could bind as a substrate analogue, with the two carboxylate oxygens mimicking the two oxygen atoms of a peroxide substrate.


The other Cys residue implicated in the proposed mechanism of action of PRDX5 (80), the Cr residue Cys151, is located in the loop between β7 and α6, and its side chain is exposed to the solvent. The proposed mechanism involves the formation of an intramolecular disulfide bond between these two Cys residues. However, the distance observed between the two Sγ atoms is 13.8 Å and the formation of the expected disulfide bond will thus not be possible without a large conformational rearrangement, as will be explained later in the description of the oxidized form of human PRDX5. This large conformational rearrangement is confirmed by the fact that freshly grown PRDX5 crystals are destroyed immediately in presence of hydrogen peroxide and survive only for a few days when they are simply exposed to air, whereas crystals of the C47S mutant in which the disulfide bond between Cys47 and Cys151 cannot occur can survive several weeks, even in presence of hydrogen peroxide. An additional Cys, Cys72, is found in the C-terminal part of the β4 strand of human PRDX5. It was shown (80) that this residue is not implicated in the mechanism, because its mutation does not modify the activity. It is thus surprising to find this Cys located close to the peroxidatic Cys, at the bottom of the active-site pocket.
The oxidized forms
From a historic point of view, the first crystal structure of the oxidized form of human PRDX5 showed unexpected intermolecular disulfide bonds between the Cp residue of one monomer and the Cr residue of another monomer and vice versa (22). The formation of this kind of dimer involved a domain swapping in which the α6 helix of one chain comes in contact with the other chain and regenerates in an intermolecular way exactly the same hydrophobic contacts with α2 and β6 that were observed in the reduced form in an intramolecular way. This movement associated with the unwinding of the N-terminal part of α2 helices brings Cys47 of one chain in close proximity to Cys151 of the other chain, allowing the formation of the disulfide bond. Because it is not clear whether this dimeric oxidized form is physiologically relevant, it is not further discussed. The crystal structure of the oxidized form with the expected intramolecular disulfide bond was described a few years later (83). In this structure, the overall fold is very similar to that of the reduced form previously described. The only differences occur in the regions of the Cp and Cr residues. The N-terminal part (residues 44 to 51) of the α2 helix containing the Cp residue Cys47 is completely unwound, allowing a movement of this residue in the direction of the Cr residue Cys151. An amplitude as large as 9.5 Å was estimated for this movement but clearly did not suffice for the formation of the disulfide bridge. It was also necessary to displace the region containing the Cr residue Cys151 in the direction of Cys47. This second movement involved residues 145 to 156 and thus included the N-terminal part of the α6 helix. The amplitude of the movement of the Cr residue is about 4 Å. The combination of these two movements finally allows the formation of the intramolecular disulfide bond illustrated in Fig. 6. This disulfide bond appears at the surface of the molecule and is thus accessible for reduction by thioredoxin. The benzoate ion has disappeared, as well as the contacts of Cys47 with Thr44 and Arg127. The large flexibility of the two regions (44 to 51 and 145 to 156) involved in the formation of this disulfide bond can be estimated from a comparison of the main-chain conformational angles φ and ψ. This is illustrated by a multiple Ramachandran plot (Fig. 7), in which an arrow joins the locations of a residue in the reduced and the oxidized forms. As expected, the origin of many arrows appears in the α−helix region. It can also be seen that some angle modifications are close to 180 degrees, the maximum possible value.

Oligomerization
In typical 2-Cys and 1-Cys peroxiredoxins, the molecules are associated as dimers, known as type B dimers, in which two β7 strands of two molecules are linked in an antiparallel mode to form a 10-stranded β-sheet. This kind of dimer was never observed for human PRDX5 in either the reduced or the oxidized form. As can be seen in Figs. 3B and 6A, the conformation of the β7-α6 loop presents a bump close to the C-terminal part of β7, which does not allow the bringing of the β7 strands of two molecules together for the formation of the large β-sheet. However, PRDX5 molecules are also associated as dimers, known as type A dimers, illustrated in Fig. 8, in which the α3 helix of one monomer comes in close contact with the α5 helix of the other one. These contacts also involve residues in the β3-α2 loop and in the β5-α4 loop. Most contacts are hydrophobic, except an intermolecular salt bridge Arg124 to Asp77. These dimers exist irrespective of the redox state, and the buried surface is approximately 730 Å2 per monomer. This kind of type A interaction also is observed during the formation of toroid decamers in reduced or overoxidized forms of typical 2-Cys peroxiredoxins by the association of five type B dimers (98). Of course, PRDX5 is unable to form this kind of decamer, because type B dimers do not exist.

Enzymatic Mechanism and Kinetics
Mammalian PRDX5 was the first characterized peroxiredoxin belonging to the atypical 2-Cys subfamily (80). Mutants of human PRDX5 in which Cys47 (Cp), Cys72, and Cys151 (Cr) were replaced by a serine revealed that oxidized Cys47 reacts with Cys151 to form an intramolecular disulfide bridge (Fig. 9), whereas in typical 2-Cys peroxiredoxins, the resolving mechanism leads to the formation of an intermolecular disulfide linkage between two monomers (80). The existence of an intramolecular disulfide bridge between Cp and Cr was confirmed in oxidized crystal forms (83). The reduction of the disulfide formed by the oxidation of PRDX5 was shown to be performed by cytosolic or mitochondrial thioredoxin but not by glutaredoxin or glutathione (20, 80, 87, 103). Interestingly, cyclophilin A was also reported as potential reductant for PRDX5 (51), but the physiologic relevance is unknown. It must be also noted that a PRDX5 reductant in peroxisomes remains to be identified with regard to the potential peroxidase activity of PRDX5 in this organelle.
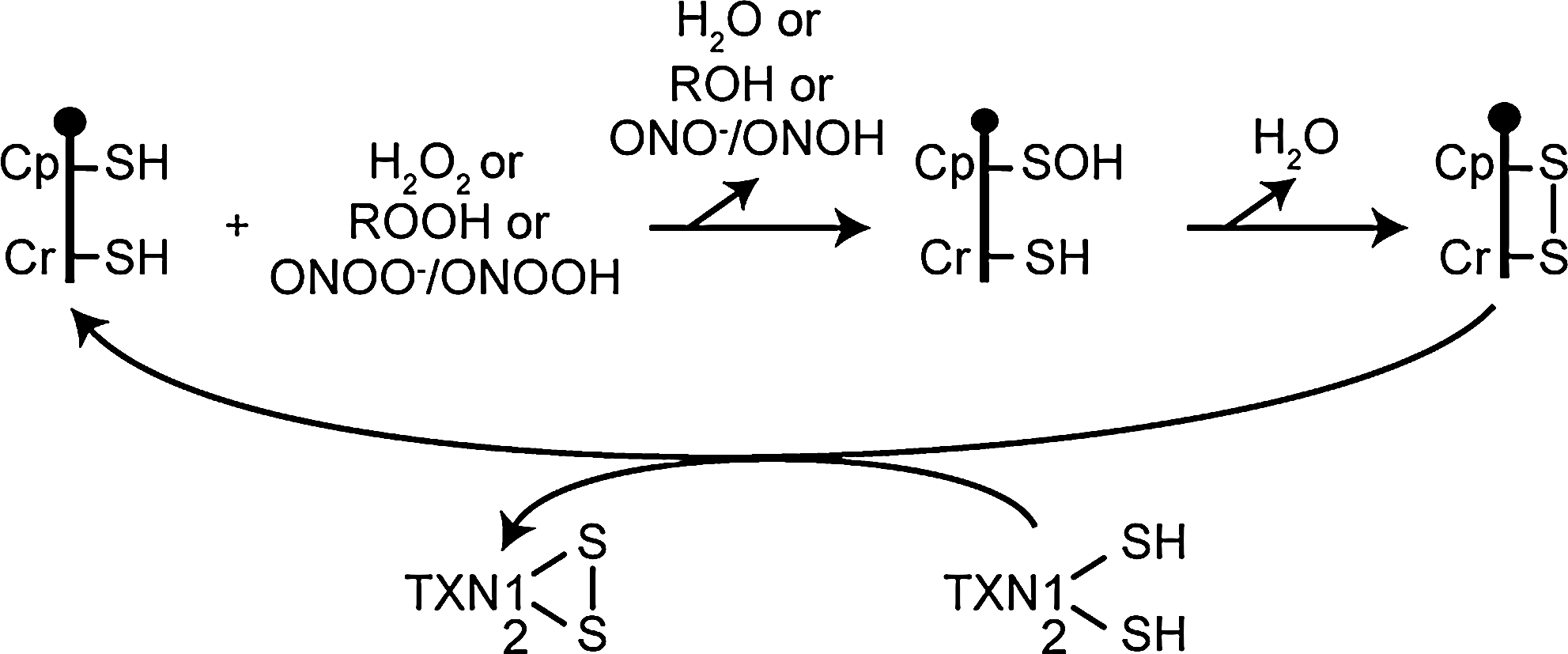
Prokaryotic and eukaryotic peroxiredoxins catalyze the reduction of hydrogen peroxide, organic peroxides, and peroxynitrite with different efficiencies (87). Human PRDX5 was the first mammalian peroxiredoxin shown to react with high rate constants with peroxynitrite in the 107 M−1s−1 range (20, 87). Human PRDX5 reacts with hydrogen peroxide more slowly (∼105 M−1s−1) than with organic peroxides (∼106 M−1s−1) or peroxynitrite. This suggests that, in subcellular compartments where PRDX5 coexists with other peroxiredoxins (cytosol, nucleus, mitochondria), the mammalian atypical 2-Cys peroxiredoxin could be involved more specifically in the reduction of peroxynitrite or lipid peroxides instead of hydrogen peroxide (14, 87).
Peroxiredoxin enzymatic activities have been shown to be regulated by posttranslational modifications. Mammalian typical 2-Cys peroxiredoxins, for example, are prone to overoxidation of their Cp in presence of an excess of hydrogen peroxide, which results in a transient (sulfinic acid) or permanent (sulfonic acid) inactivation of the enzyme (94). The reduction of the sulfinic acid can be catalyzed by sulfiredoxin (9). It has been proposed that mammalian typical 2-Cys peroxiredoxins are more susceptible to hyperoxidation than are peroxiredoxins lacking a C-terminal YF motif that interacts with a GGLG motif in a loop neighboring the Cp. These motifs and other structural features would participate in the kinetic pause in disulfide bond formation, leading to overoxidation of Cp (97). These domains are lacking in prokaryotic peroxiredoxins but also in PRDX5 and PRDX6 in mammals. It appeared that mammalian PRDX5 is more resistant to the sulfinylation by hydrogen peroxide than are typical 2-Cys peroxiredoxins (95). Accordingly, it was also shown that sulfiredoxin does not interact with PRDX5 (94). Although the activity of mammalian typical 2-Cys peroxiredoxins may be modulated by different posttranslational modifications (72), little is known about such possible modifications of PRDX5. Interestingly, PRDX5 may undergo glutathionylation in rat hepatocytes exposed to oxidative stress (25), but the functional significance of such posttranslational modification is still to be investigated.
Functions
Cytoprotective antioxidant function of PRDX5 was first investigated after human PRDX5 was characterized as a peroxidase but also because of the presence of this enzyme in mitochondria and peroxisomes, two organelles that are major intracellular sources of ROS/RNS (reactive nitrogen species). Cellular peroxidase activity of human PRDX5 was first investigated in NIH3T3 and HeLa cells by overexpressing human PRDX5 that inhibited hydrogen peroxide accumulation induced by tumor necrosis factor (TNF)-α and PDGF (80) or p53 (103). Moreover, it was also reported that human PRDX5 expressed in mitochondria or in the cytosol of Saccharomyces cerevisiae protected yeast cells from oxidative stress induced by paraquat (85). Overexpression of human PRDX5 in the cytosol, in mitochondria, in the nucleus of CHO or HT-22 cells (4, 104), or in the cytosol of tendon cells (101) was shown also to reduce significantly cell death caused by exogenous peroxide exposure. Interestingly, exogenous administration of recombinant human PRDX5 was demonstrated to provide protection against ibotenate-induced excitotoxic stress in mouse brain (69). Also, PRDX5-expression modulation in mitochondria or in the nucleus of cell lines exposed to prooxidant drugs or peroxides revealed that the enzyme may confer protection against mitochondrial or nuclear DNA damage (3, 4, 45). Moreover, cells with reduced expression of PRDX5 were shown to be more prone to oxidative damage and apoptosis induced by etoposide, doxorubicin (Adriamycin), hydrogen peroxide, and the mitochondrial toxin MPP+ (2, 15, 43, 81). In Xenopus embryos, heterologous expression of human PRDX5 protects cells against alcohol-induced ROS/RNS production (67, 68). Finally, orthologous PRDX5 in Drosophila melanogaster, that is expressed in different cellular compartments among which mitochondria, the nucleus and the cytosol, was demonstrated not only to protect against oxidative stress and apoptosis but also to promote longevity of the fly (71). It must be noted that yeast Pmp20, a 1-Cys peroxiredoxin homologous to human PRDX5 and localized in peroxisomes of methylotrophic Candida boidinii, Hansenula polymorpha, or Pichia pastoris, has been reported to play a major antioxidant cytoprotective role, emphasizing the functional importance of peroxiredoxins in these organelles (5, 32, 100).
It is now recognized that hydrogen peroxide produced in mammalian cells in response to different stimuli may serve as signaling molecule (73). Although PRDX5 was initially shown to modulate peroxide levels produced in cells after stimulation by PDGF, TNF-α, or p53 (80, 103), few data are available on a possible role of mammalian PRDX5 as a redox sensor. Moreover, it must be noted that mammalian PRDX5 seems to lack important features to be a good candidate as modulator of peroxide-dependent signal transduction. As discussed earlier, contrary to mammalian typical 2-Cys peroxiredoxins, PRDX5 appears to be more resistant to overoxidation by hydrogen peroxide (95), and, so far, PRDX5 is not recognized as a good substrate for kinases, such as cyclin-dependent kinases regulating the peroxidase activity of typical 2-Cys peroxiredoxins (8, 96). However, we cannot totally exclude that mammalian PRDX5 may act in redox signaling mechanisms as a peroxidase-based protein-oxidation relay (28) by using a mechanism similar to the one identified in yeast between peroxidase Orp1 and transcription factor Yap1 (86). It is tempting to hypothesize that the translocation of PRDX5 into the nucleus, as described for example in thyrocytes under pathophysiologic oxidative stresses (26, 70), may be involved in redox-dependent transcription regulation. Further studies will be necessary to investigate this possible function.
The existence of different enzymes able to decompose peroxides in the same subcellular compartment, such as the mitochondrion, the peroxisome, or the cytosol, obviously raises questions about the redundancy and thus the specific function of these antioxidant enzymes. In mitochondria, PRDX5 coexists with other peroxidases, namely PRDX3, GPX1, and GPX4 (14). Mitochondrial localization of PRDX5 seems to be quite well conserved throughout evolution, as predicted mitochondrial targeting sequences have been also identified in orthologous PRDX5 in invertebrates (71, 102) (see also Fig. 1). A very informative kinetic hierarchy of mitochondrial reductases of hydrogen peroxide was proposed recently, based on known rate constants and enzyme abundance (14). Obviously, the hierarchy must be adapted to cell types, as this ranking depends on abundance of each peroxidase and the availability of reducing equivalents for peroxidase recycling. In this analysis, mitochondrial PRDX5 would play a minor role in the reduction of hydrogen peroxide compared with PRDX3 or GPX1. However, in this organelle, PRDX5 high rate constants with alkyl hydroperoxides and peroxynitrite could confer to this peroxidase a more-specific function in peroxide reduction (15, 87). Moreover, one should also take into account a possible heterogeneous distribution of peroxidases inside mitochondria and consider also different susceptibilities to inactivation. It is clear that such speculations will need experimental validation.
In the peroxisomal matrix, PRDX5 may assist catalase in peroxide reduction. Catalase is a very efficient hydrogen peroxide reductase, whereas PRDX5 is an efficient alkyl hydroperoxide and peroxynitrite reductase (20, 87). In methylotrophic yeasts, Pmp20 peroxiredoxin appeared, as emphasized earlier, to play a major role as protective antioxidant enzyme when yeast cells grow with methanol that is metabolized by peroxisomal enzymes (5, 32, 100). Studies investigating the effects of human S-PRDX5 overexpression were not able to target the protein only to this organelle. Overexpression of the short form of human PRDX5 addresses the protein to the cytosol, the peroxisomes, and the nucleus (4, 46, 99, 101). Moreover, even if PRDX5 acts as a peroxidase in the peroxisomes, a reductant is still to be identified as, to our knowledge, no thioredoxin system has been described in peroxisomes.
In the cytosol of mammalian cells, PRDX5 may coexist with PRDX1, PRDX2, and PRDX6 (72). Thus, the specific role of PRDX5 in this subcellular compartment is not clear. However, according to enzyme kinetics (87) and on accumulating data reporting that PRDX1 and PRDX2 are involved in hydrogen peroxide signaling in mammalian cells (73, 96), cytosolic PRDX5 as well as PRDX6 should rather be considered cytoprotective antioxidant enzymes (2, 4, 58) (see also the article by A. Fisher in this Forum), although this view may be not generalized to all mammalian cell types.
Finally, the function of PRDX5 in the nucleus is still a matter of debate. Initially, PRDX5 was described as a DNA-binding protein able to repress transcription of retrotransposons in human cells (44) before its peroxidase activity was established. Also, consistent with a role as a protective antioxidant enzyme, overexpression of PRDX5 in the nucleus protects DNA from oxidative damage caused by peroxides (4), and silencing the gene stimulates formation of etoposide-induced double-strand DNA breaks (47). It is important to note that in yeast, peroxiredoxins appeared also to play a major role against oxidative attacks leading to DNA damage and genome instability (7, 33, 34, 93). However, PRDX5 is not expressed in the nucleus at high levels in all cell types in mammals (4). Interestingly, translocation into the nucleus under pathophysiologic situations, for example in thyrocytes (26, 70), suggests that PRDX5 may have a function in the nucleus either as a DNA-protective enzyme or as a redox-dependent signaling molecule acting possibly as a peroxidase-based protein oxidation relay (28).
An unexpected role of PRDX5 in immunogenicity in human metastatic melanoma appeared more recently with the identification of an antigen generated by a point mutation in PRDX5 in human melanoma lacking the wild-type allele (77). It was suggested that this tumor antigen could be an immunogenic target for T cell–specific immunotherapy (76). Moreover, as mentioned earlier, the alternative splicing variants of PRDX5 expressed in human cells appeared (61) to be involved in immune regulation (78). Through the generation of short antigenic peptides encoded by these spliced PRDX5 isoforms, it was proposed that an immune-mediated response to stress, including oxidative stress, may contribute to the detection and elimination of neoplastic cells or even damaged normal cells (78).
Conclusions and Perspectives
To conclude, PRDX5 is the last-discovered peroxiredoxin in mammals and the only mammalian atypical 2-Cys peroxiredoxin. In humans, PRDX5 exhibits a remarkably wide subcellular distribution with a localization in the cytosol, in mitochondria, in peroxisomes, and in the nucleus. PRDX5 is a peroxidase that uses cytosolic thioredoxin 1 or mitochondrial thioredoxin 2 as physiologic reductants to reduce alkyl hydroperoxides or peroxynitrite with high rate constants, whereas the PRDX5 reaction with hydrogen peroxide is slower. Although PRDX5 was shown to be constitutively and ubiquitously expressed in virtually all mammalian tissues, up- and downregulation have been reported in different pathophysiologic situations. Moreover, PRDX5 overexpression protects cells against nitrooxidative stress, whereas a decreased expression makes cells more vulnerable. These observations as well as biochemical and structural features suggest that in mammalian cells, PRDX5 may act mainly as a cytoprotective antioxidant enzyme like prokaryotic peroxiredoxins, whereas typical 2-Cys peroxiredoxins would rather play the role of redox sensors in hydrogen peroxide-dependent intracellular signaling. It is clear that further studies will be required to gain insights into the specific functions of PRDX5 in subcellular compartments to which other peroxidases are also targeted. Finally, although knockout mice exist for the five other peroxiredoxins, so far, PRDX5 knockout mice or transgenic mice overexpressing PRDX5 are lacking but would be necessary to understand better the functions of the unique mammalian atypical 2-Cys peroxiredoxin.
Footnotes
Acknowledgments
The research of the authors is financially supported by the Fonds National de la Recherche Scientifique (FNRS), by the Communauté française de Belgique-Actions de Recherche Concertées (ARC), and by the DIANE research program of the Walloon Region.