
Abstract
The major histocompatibility complex class I molecule human leukocyte antigen (HLA)-B27 is strongly associated with a group of inflammatory arthritic disorders known as the spondyloarthropathies. Many autoimmune diseases exhibit associations with major histocompatibility complex molecules encoded within the class II locus with defined immune responses either mediated by T or B-lymphocytes. Despite the association being known for over 30 years, no defined immune response and target autoantigens have been characterized for the spondyloarthropathies. Thus, the mechanism and role of HLA-B27 in disease pathogenesis remains undetermined. One hypothesis that has recently received much attention has focused around the enhanced propensity for HLA-B27 to misfold and the increased tendency of the heavy chain to dimerize. The misfolding of HLA-B27 has been associated with its redox status and this is postulated to be involved in disease development. Here we discuss the impact of the redox status on HLA-B27 biosynthesis and function. Antioxid. Redox Signal. 15, 669–684.
Human Leukocyte Antigen-B27 Structure, Function, and Inflammatory Arthritis
Human leukocyte antigen (HLA)-B27 is a member of the HLA-B family of alleles encoded within the MHC locus. Like all other HLA class I alleles present in the human population, HLA-B27 assembles within the oxidizing environment of the endoplasmic reticulum (ER). MHC class I heavy chains can be structurally divided into three extracellular domains, named α1, α2, and α3, and a transmembrane and cytoplasmic domain. MHC class I heavy chains usually contain four conserved cysteines (C, cys), which form structurally important disulfide bonds. The conserved cys residues are within the α2 and α3 domains, forming disulfide bonds between C101-C164 and C203-C259, respectively (Fig. 1). However, most HLA class I molecules possess at least one additional unpaired cys residue (Table 1). Most HLA-A molecules possess an additional unpaired cys within the cytoplasmic domain at p329. Intriguingly, HLA-C can have up to an additional five unpaired cys residues located throughout the heavy chain, whereas HLA-G has an unpaired cys at p42 within the α1 extracellular domain (Table 1). HLA-B alleles usually have two additional cys, either at positions 67 (extracellular) and 308 (transmembrane/cytoplasmic tail border) or 308 and 325 (cytoplasmic tail), but HLA-B27 is unusual in that it possesses all three of these unpaired cys (Table 1). It is these unpaired cys residues, especially C67, that have generated interest in the influence of the redox state and environment on the structure of HLA-B27. This article will discuss how both the structural and unpaired cysteine residues of HLA-B27 impact on its redox chemistry and the consequences for both folding and misfolding events.

Unpaired cysteines found within major histocomptability complex class I alleles A, B, C, and D are marked in bold.
Unpaired cysteines of HLA-B27 are shown with underline, bold, and italics.
HLA, human leukocyte antigen.
HLA-B27 exhibits a strong association with a group of inflammatory arthritic disorders known as the spondyloarthropathies (SpAs) (24, 129). The association is one of the strongest known between a gene encoded within the MHC locus and an autoimmune disorder, and remains one of the most unusual associations for several reasons. Most autoimmune diseases exhibit associations with MHC class II molecules, which present peptides to CD4+ T cells and exhibit immune responses either mediated by T or B-lymphocytes. No defined immune response and target autoantigens have been characterized for the SpAs. The related reactive arthritis, another of the SpAs, is triggered after a bacterial infection (23). In the case of reactive arthritis, a number of defined pathogenic triggers have been identified, but the immune response remains poorly characterized. Thus, despite the association being known for over 30 years, the mechanism and role of HLA-B27 in SpA pathogenesis remains undetermined.
One hypothesis that has received much attention recently has focussed around the enhanced propensity for HLA-B27 to misfold and the increased tendency of the heavy chain to dimerize. It has been proposed that HLA-B27 misfolding and possibly dimer formation induce a group of stress responses within the ER, leading to the induction or upregulation of proinflammatory cytokines (30, 31, 80, 132, 133, 149, 157). This review will discuss the misfolding characteristics of HLA-B27, the elements that can govern its redox state and highlight how these may be critical in disease development.
HLA-B27 Associations with Early Folding Chaperones
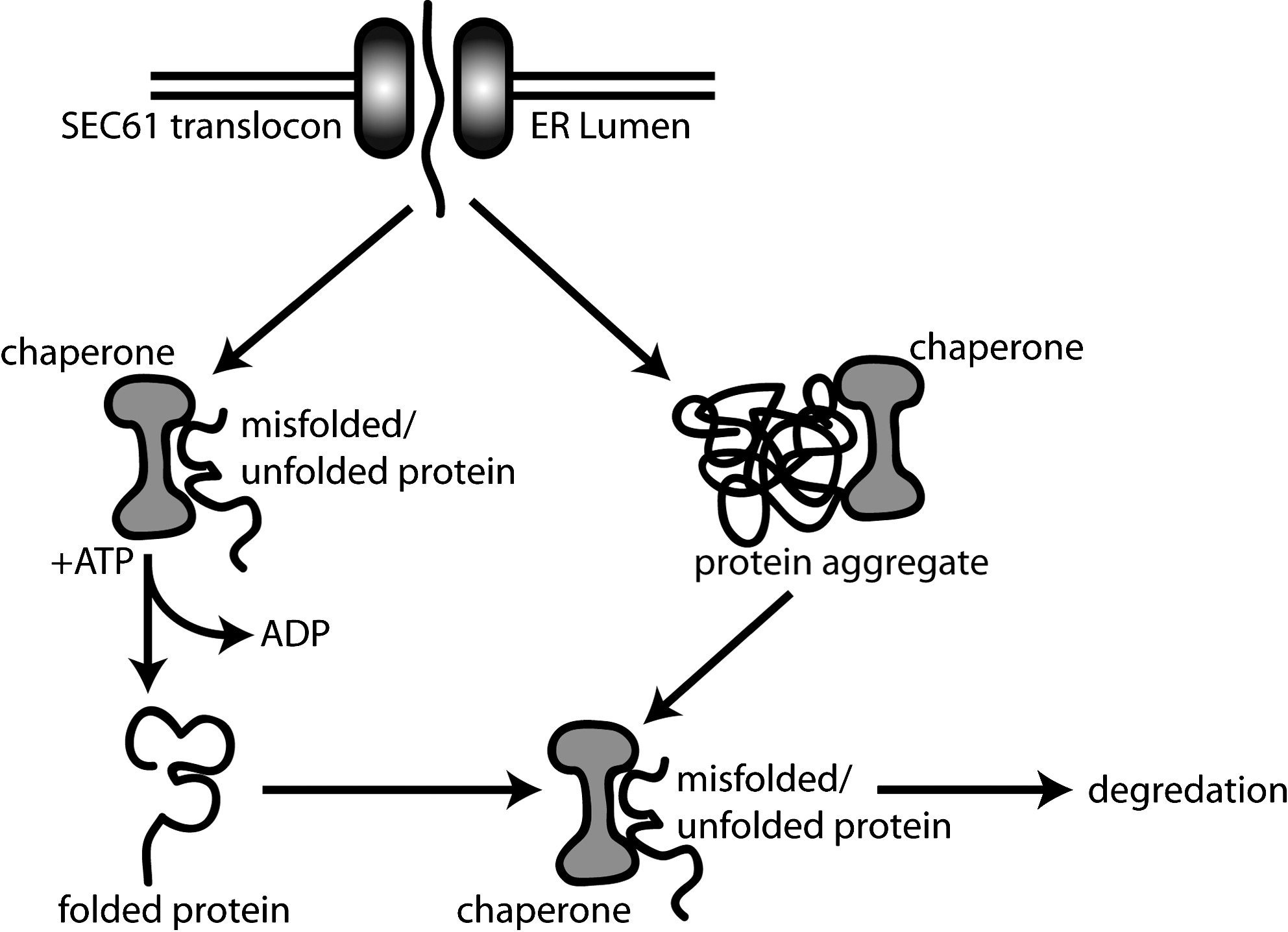
MHC class I folding is a closely regulated ER chaperone-mediated event. Chaperones recognize and bind non-native proteins and assist in their conformational processing aided by ATP hydrolysis and cochaperones. In addition, it is becoming apparent that chaperones can regulate the degradation of misfolded proteins (Fig. 2). MHC class I folding, as we currently understand it, occurs in two main phases: (i) an early folding stage, which predominantly involves the lectin chaperone calnexin (CNX), the immunoglobulin binding protein (BiP), and ERp57, a member of the protein disulfide isomerase (PDI) family of oxidoreductases, and (ii) a latter folding stage, involving the formation of an MHC class I-specific scaffold called the peptide loading complex (PLC) (10).
The CNX-ERp57 cycle
CNX is thought to normally associate with nascent glycoproteins via a monoglucosylated oligosaccharide composed of Glc1Man5–9GlcNAc2. CNX has an extended proline-rich domain (P domain) that recruits the oxidoreductase, ERp57 (109, 117). The positioning of the P domain with respect to the protein substrate enables ERp57 to interact within a protein folding cage (59, 121). It has been postulated that ERp57 dissociates rapidly from proteins that have misfolded but persists on those undergoing productive folding reactions. However, the opposite is thought to be true for CNX-protein interactions (45).
Monoglucosylated oligosaccharide units exist transiently on asparagine–X–serine oligosaccharide acceptor sites. In the case of HLA class I molecules, the oligosaccharide acceptor site is located at position 86–88 on an external loop of the α1 domain. Monoglucose subunits can be generated in two ways, first by the trimming of two glucose residues from the precursor N-linked oligosaccharide Glc3Man9GlcNAc2 by glucosidases I and II (41, 56, 82). Once a native conformation has been attained, the substrate is released from the CNX/ERp57 cycle by removal of the third mannose linked glucose via the activity of glucosidase II (56, 82).
The second pathway of generating monoglucosylated proteins is via UDP glucose:glycoprotein glucosyltransferase (UGGT), a protein folding sensor that can detect substrates that have not acquired a native conformation, leading to reglucosylation and further rounds of CNX binding and folding (82, 91). Recently, using UGGT mouse knockout cell lines, protein substrates entering the CNX/ERp57 cycle have been classified into three categories: (i) proteins that complete their folding in one binding event, (ii) proteins requiring multiple rounds of folding, and (iii) proteins that UGGT deletion delays release from CNX, presumably requiring some structural maturation before release from the folding cycle (134).
HLA-B27 and CNX
Newly synthesized MHC class I heavy chains associate with CNX (37, 58, 68) and can also bind in a lectin independent manner via secondary CNX-protein interactions (81, 156). MHC class I molecules represent an unusual group of protein substrates as they attain several different conformations during their assembly. As the final structure is determined by their bound peptides, which will often be of a different affinity to those bound early on in the folding process (85), the chaperone machinery must be capable of distinguishing the many different conformations that MHC molecules can attain either during the folding process and acquisition of peptide from those structures that indicate MHC class I molecules are terminally misfolded.
Early studies using mouse MHC class I molecules indicated that CNX enhanced folding, reduced aggregation, and protected the heavy chain from ER-associated degradation (ERAD), the process by which misfolded proteins in the ER are removed to the cytosol and degraded by the proteasome (68, 151). However, there does appear to be undefined differences in the activity of mouse and human CNX, as castanospermine, a glucosidase inhibitor that perturbs the formation of monoglucose subunits, has a more pronounced effect on mouse in comparison to human MHC class I molecules (151). This could be due to differences in glycan site usage, with human and mouse MHC class I molecules possessing one and three glycosylation sites, respectively. The reason for these species differences in glycan site usage remains speculative, but could reflect a greater dependence on CNX function for mouse MHC class I folding. Differences between mouse MHC class I alleles in their requirement for CNX also occur (151). Using human cell lines lacking CNX, it has been shown that cell surface expression of MHC class I molecules (123, 130), including HLA-B27, are unaffected (Guiliano, manuscript submitted).
HLA-B27 and ERp57
The demonstration that ERp57 was part of MHC class I assembly process led to the proposal that this oxidoreductase was responsible for MHC molecule disulfide bond formation (64, 87, 104). ERp57 is composed of the a, b, b′, and a′ domains along with an acidic C-terminus. The a and a′ domains contain the reactive CXXC motifs. ERp57 is predominantly found in a reduced state within the ER (8, 69), and thus its functions are limited to reduction and/or isomerization of disulfide bonds. Therefore, the direct association between MHC class I and ERp57 may serve a role other than the formation of conserved structural disulfide bonds. ERp57 knockout or RNA interference knockdown lines express MHC class I at their cell surface without apparent drastic changes to their redox states (49, 158). However, it was noted that the rate of formation of the mouse MHC class I α3 domain C203-C259 disulfide bond was reduced in the absence of ERp57 (158). MHC class I heavy chain oxidation can occur in the absence of CNX and calreticulin, a soluble paralog of CNX, which can also recruit ERp57. MHC class I heavy chain oxidation was catalyzed normally in the presence of an ERp57 R282A mutant, which has lost the ability to associate with these lectin chaperones. Thus ERp57 may potentially catalyze MHC class I heavy chain oxidation in the absence of CNX (and calreticulin) interactions (159). However, oxidation reactions catalyzed by other oxidoreductases in the absence of CNX and calreticulin cannot be fully discounted (72).
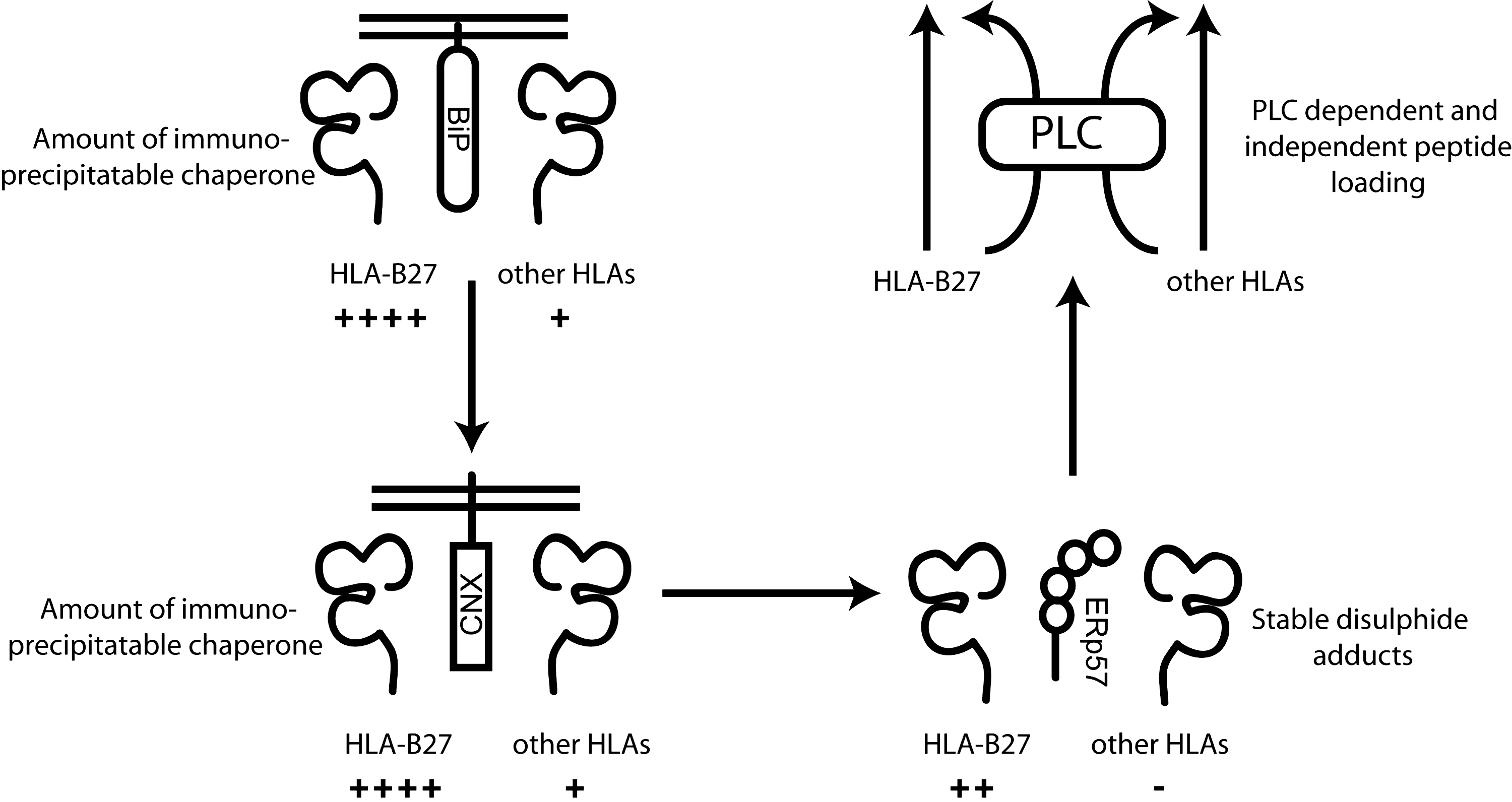
When compared to other HLA class I alleles, the interactions between HLA-B27 heavy chains, CNX, and ERp57 are unusually strong or long lived (7) (Fig. 3). We have shown that ERp57 can directly associate with HLA-B27, but is barely detectable with other MHC class I molecules such as HLA-A2 (7). ERp57 can form direct conjugates with HLA-A2 only if individual structural cys residues have been mutated, thus leaving unpaired free thiol groups free to associate with ERp57 (11). These experiments further demonstrated that the α2 domain C101 and C164 residues were important in determining the ERp57 association (11, 74). It is likely that ERp57 binds directly to HLA-B27, probably via C101 and/or C164, with these residues being exposed for prolonged periods, enabling the detection of these conjugates. However, the presence of C67 within the α1 domain raises the possibility that this additional cys residue could in fact stabilize or enhance the ERp57 interaction. Removal of C67 did not abrogate the ERp57 interaction (7) and does not seem to enhance ERp57 binding. This was further confirmed by replacing the valine (V) to cys at p67 within HLA-A2. Immunoprecipitation experiments failed to detect any HLA-A2.V67C-ERp57 conjugates at 37°C (7). Therefore, the ERp57-HLA-B27 heavy chain interactions probably occur through the breaking of non-native disulfide bonds and not via interactions with free cysteine residues.
HLA-B27 and BiP
MHC molecules can associate with BiP, a multifunctional soluble chaperone with roles in handling folding and misfolding proteins, and controlling ER homeostasis during conditions, leading to perturbation of protein folding. BiP contains an N-terminal ATPase domain and C-terminal peptide binding domain (110). The ATPase domain exists in an open conformation that binds ATP, which when hydrolyzed to ADP via the actions of ER-resident J-domain cochaperones (Erdj1-7), closes to a conformation with high substrate affinity (91). BiP interacts with newly synthesized proteins immediately and transiently (57), probably reflecting its role as a molecular rachet allowing protein translocation into the ER (96). However, the association with misfolding protein substrates is prolonged (135). How BiP partitions its functions associated with folding and misfolding after protein translocation is thought to depend on the activity and substrate specificity of the ERdj family of cochaperones (91). To date, ERdj3/6 are thought to be involved with folding, whereas ERdj4/5 greatly influence degradation of misfolded substrates via ERAD (40, 110, 150, 151).
The role of BiP in folding/misfolding is intimately involved with its role as the guardian of the unfolded protein response (UPR), which is activated by ER stress. BiP interacts with three key UPR effectors, the transcription factor ATF6, and the kinases IRE1 and PERK (61, 90). On activation of the UPR, BiP is upregulated, but also dissociates from ATF6, IRE1, and PERK, leading to their activation (16, 61). Interestingly, the BiP cochaperones ERdj3, 4, 5, and 6 are also induced by the UPR, which may reflect their roles in quality control either through the promotion of productive folding events or degradation of misfolding proteins (110).
BiP can be found in association with HLA-B27 more readily than other MHC class I heavy chains (Fig. 3). This association was noted to mimic the association between BiP and immunoglobulin; that is, slow folding immunoglobulin molecules exhibited a stronger and more prolonged association with BiP (7, 60). This was also true for HLA-B27, which was demonstrated to have a very slow maturation phenotype, which correlated with BiP association (7). This correlation can be traced back to one of the original descriptions of BiP associating with MHC class I molecules, where Nossner and Parham noted that HLA-B7, when expressed in the CIR cell line, exhibited slow maturation kinetics and BiP-HLA-B7 associations could readily be detected (107). Therefore, BiP predominantly associates with misfolding HLA molecules, probably at early stages of the assembly process as suggested for other misfolding substrates (135). The association with misfolding HLA-B27 could be part of BiP sensing such molecules as part of the UPR. If so, this interaction could be responsible for triggering the UPR, as has been detected in bone marrow-derived macrophages from HLA-B27 transgenic rats (148, 149).
Crosstalk exists between BiP and the CNX-ERp57 chaperone systems. For instance, proteins can associate sequentially with BiP and CNX (57). Both these chaperones can synergistically suppress the aggregation of nonglycosylated substrates in vitro (138). However, the relationship between MHC class I molecules that associate with BiP and those that associate with CNX remains undetermined. As HLA-B27 can associate with both BiP and CNX, it is possible there is a sequential association with these chaperones.
HLA-B27 and the PLC
Once β2m associates with the MHC class I heavy chain, CNX is displaced and replaced by a soluble lectin chaperone calreticulin, which has sequence and structural similarities to CNX (91). Calreticulin can also recruit ERp57 via a proline-rich P domain. Together with calreticulin and ERp57, the partially folded MHC class I molecules are tethered to the transporter associated with antigen processing (TAP) heterodimer via the MHC class I-specific accessory molecule tapasin, forming the PLC (Fig. 4). These components associate in a co-operative manner, which probably has the added advantage of increasing the affinity of the complex for MHC class I molecules. However, the co-operativity of PLC formation means that study of individual components is often somewhat problematic.
Tapasin the central scaffold of the PLC
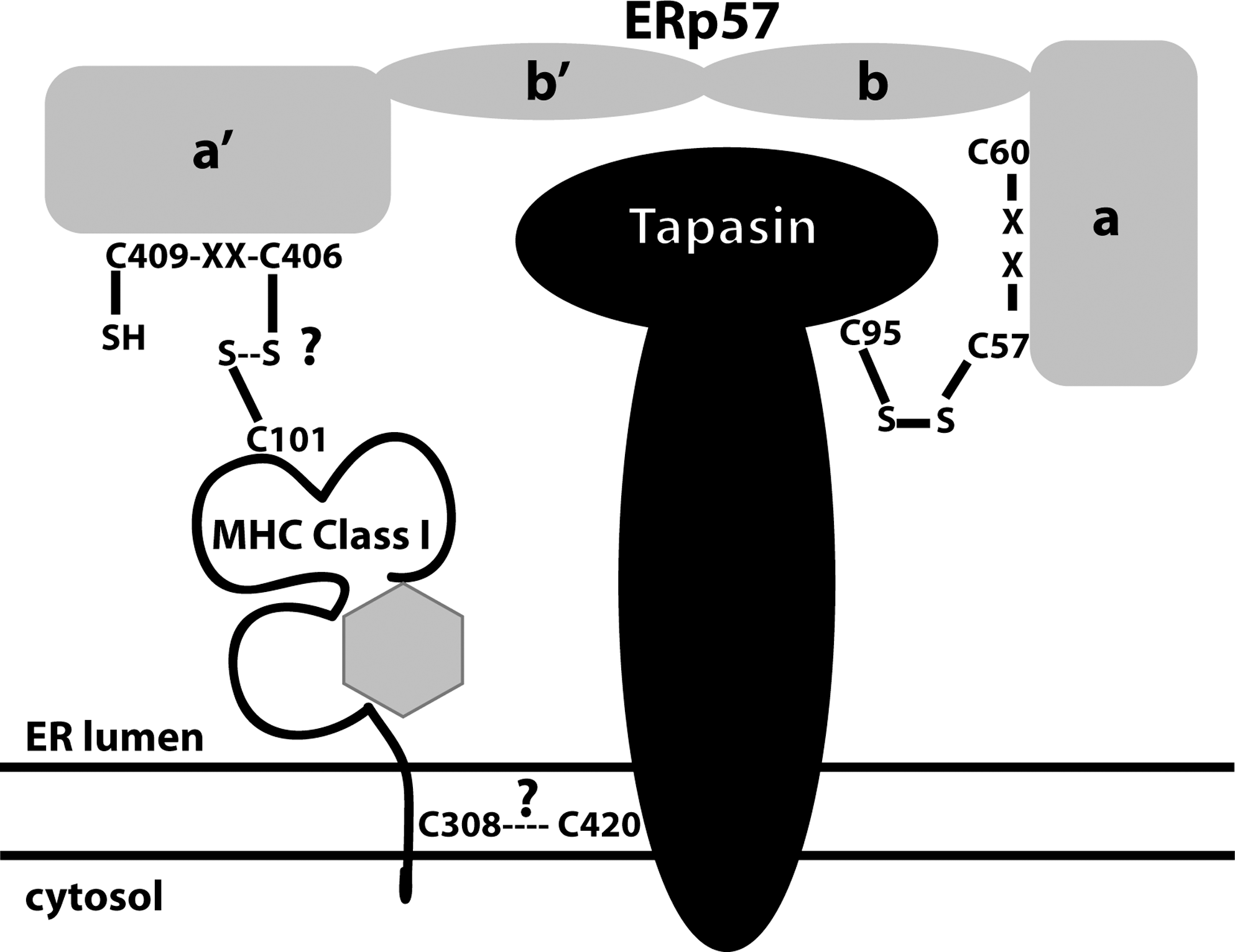
Tapasin is the central scaffold around which the PLC is composed and has several roles (122), including stabilization and enhancing peptide transport via the TAP heterodimer (84). Tapasin is predominantly found disulfide bonded to ERp57 via an unpaired cysteine at position 95, linked to C57 of the N-terminal CXXC redox active motif of ERp57 (8, 38) (Fig. 4). This association prevents ERp57 from undergoing a redox escape reaction, the normal method for resolving oxidoreductase-substrate interactions, thus stabilizing the tapasin-ERp57 interaction (115). It was proposed that the redox activity of ERp57 in conjunction with tapasin was involved in peptide loading and that the free C-terminal CXXC redox active motif of ERp57 participated in this step (Fig. 4). This was supported by the finding that HLA-B*4402 (a subtype of the HLA-B44 family of HLA-B class I alleles) had an altered redox state when it associated with a PLC containing a tapasin molecule lacking cys95 and was thus unable to bind to ERp57 (38). It was further suggested that the ERp57-tapasin conjugate inhibited ERp57 reduction of MHC class I molecules (74). However, on closer examination, very little of HLA-B*4402 exhibited an altered redox state in the PLC containing an ERp57 molecule with an inactive C-terminal CXXC motif and an ERp57-deficient binding tapasin molecule. It was concluded that MHC class I recruitment into the PLC, peptide loading, and editing were independent of the redox activity associated with ERp57. However, an intact tapasin-ERp57 conjugate was required for efficient MHC class I recruitment into the PLC (114).
Perhaps the most important function of tapasin and thus the PLC is that of peptide optimization, whereby relatively low affinity peptides are exchanged for optimal peptides (155). The tapasin-ERp57 conjugate could indeed be considered as a functional unit, as assay systems revealed that this conjugate was more effective in promoting peptide optimization than tapasin alone (153). This was further supported by the crystal structure of the tapasin-ERp57 conjugate, which showed extensive contacts between tapasin and the a and a′ domains of ERp57 (39). Peptide optimization is thought to occur via the ability of tapasin to monitor conformational changes occurring in the F pocket of the peptide binding groove via a series of hydrogen bonds (42). It is these residues within the F pocket that are thought to determine whether MHC class I molecules are tapasin dependent or independent. An important point to stress, however, is that MHC class I molecules that have been categorized as tapasin independent will tend to use tapasin if it is present (155).
HLA-B27 and tapasin
Whether HLA-B27 is tapasin dependent or independent remains debatable (19, 116) (Fig. 3). However, in the presence of tapasin, the HLA-B27 peptide cargo can be optimized (111, 112, 155). Our studies indicate that at steady state, HLA-B27 is in conformation(s) with exposed cys residues (46). How this relates to the conformation that enters the PLC remains undetermined. Interestingly, several subtypes of HLA-B27 exhibit polymorphisms within the F pocket, especially at p114 and 116, which are thought to determine tapasin dependency (111, 112, 155). Two HLA-B27 subtypes of interest are HLA-B*2706 and 09, which possess polymorphisms at p114 and 116, and do not exhibit an association with the SpAs. How these subtypes interact with the PLC has not been clearly demonstrated, but it is possible that these residues can change the association and kinetics of interaction between HLA-B27 and the PLC. An extensive analysis of HLA-B27 polymorphisms did not demonstrate a clear correlation between thermostability of the peptide cargo and interactions with tapasin. Interestingly, polymorphisms associated with the HLA-B*2706 and HLA-B*2709 subtype did exhibit lower thermostability but faster folding kinetics and enhanced tapasin interactions (47).
Studies from our laboratory indicate that the residues expressed at p114 and 116 can have a profound influence on the redox status of HLA-B27 and suggest that MHC binding peptides can influence the redox chemistry of MHC molecules (46). A role for peptides influencing the redox status was suggested where MHC class I molecules entering the PLC and had thus not acquired an optimal peptide cargo were in a partially reduced state (38). This latter point may be of importance as recent genetic analysis revealed that the endoplasmic reticulum associated aminopeptidase (ERAP1)/ARTS1 also exhibits a strong association with the SpAs (25, 27). ERAP1 has two reported functions; (i) cleavage of cytokine receptors and (ii) generation of optimal MHC class I-associated peptides from N-terminally extended sequences in the ER (18, 53, 54, 71, 131). How this genetic association translates into HLA-B27 activity remains to be resolved, but poor peptide loading could adversely affect the redox status of HLA-B27. We have recently observed increased ERAP1 levels in dendritic cells of some HLA-B27-positive ankylosing spondylitis patients, further raising the possibility of an altered peptide supply (Powis et al., submitted).
The conserved structural C101-C164 disulfide bond within the MHC class I peptide-binding groove is thought to be stabilized after acquisition of suitable peptide, possibly within the PLC (38). In the absence of tapasin, or a tapasin-ERp57 conjugate, the C101-C164 disulfide bond in the tapasin-dependent HLA-B*4402 allele was shown to be in a reduced state. In contrast, under similar conditions the tapasin-independent allele HLA-B*4405 was fully oxidized. Thus, the interaction between tapasin and ERp57 was postulated to protect the C101-C164 disulfide bond from ERp57-mediated reduction (74).
The suggestion that the C101-C164 disulfide bond could be stabilized by peptide binding within the PLC led to the idea that a potential trimolecular species might exist between MHC class I heavy chains and the ERp57-tapasin conjugate (Fig. 4). Using HLA-B27, such a species was detected and originally proposed to occur via C101 of the heavy chain and the a′ CXXC reactive motif of ERp57 (125) (Fig. 4). The same trimolecular species was again detected with HLA-B35 using an in vitro cell-free translation system (29). However, these latter observations suggested that MHC class I heavy chains interact with ERp57-tapasin via a disulfide bond between the MHC class I heavy chain transmembrane C308 residue and possibly C420 of tapasin (29). The latter observation has important consequences for the proposed model of tapasin optimization and the concept of tapasin dependency/independency of the PLC. Such a model would provide a function for C308, which is found predominantly in HLA-B alleles, but not in HLA-A alleles. This raises the question of how MHC alleles other than the B alleles interact with the PLC, if they lack C308? It is possible that HLA-B27 does indeed interact with tapasin via C308, as this was not formally tested. However, C308 can also undergo palmitoylation, a posttranslational lipid modification whereby a 16-carbon fatty acid palmitate is covalently conjugated to cys resides through a thioester linkage (52). In addition, C308 and C325 within the cytoplasmic domain were demonstrated to be important in egress from the ER and in recycling at the cell surface. Thus, if such a modification is predominant at C308, it may suggest a regulatory process for interactions with tapasin. It is clear, however, that some MHC class I molecules undergo palmitoylation, whereas others do not (73).
The redox state and the enhanced flexibility of the antigen binding groove of HLA-B*2705 (43, 44) may allow for a ternary complex to form with ERp57-tapasin via the α2 domain cys residues. Furthermore, the two most studied B alleles from which many of the ideas of tapasin activity have been formulated, HLA-B*4405 and 02, both encode a cys at position 308. Thus, the work of Chambers et al. raises the following question; how does peptide binding and optimization translate into a signal leading to the reduction of the heavy chain-tapasin disulfide bond? A disulfide bond between MHC class I heavy chains and tapasin would have to be reduced to release fully folded molecules from the ER to the cell surface. Conformational changes can be transmitted via protein transmembrane segments and this has been hypothesized for MHC class I molecules. Once MHC molecules are loaded with the appropriate peptide cargo, this may instigate release from tapasin. ATP hydrolysis by the TAP heterodimer is also thought to allow the release of folded MHC molecules from the TAP complex via conformational changes within the TAP heterodimer (1, 6, 75). Perhaps the combination of peptide binding and ATP hydrolysis are sufficient to allow reduction of the disulfide bond, but again this would depend on the lability of such a bond. C308 is at the ER membrane-cytosol boundary, and therefore the reducing environment of the cytoplasm could influence a direct interaction between MHC class I heavy chains and tapasin.
Calreticulin and the PLC
Calreticulin is an important member of the PLC (122), as its absence leads to rapid transit of MHC class I molecules from the ER to the cell surface. Although the PLC can form in the absence of calreticulin, MHC class I molecules were shown to have a low affinity peptide cargo (48). Calreticulin does not appear to depend on ERp57 binding and lectin interactions to become incorporated into the PLC (159). Calreticulin is not adequately recruited to the PLC in the absence of ERp57, and MHC class I molecules can also transit quickly through the secretory pathway with suboptimal peptides (49), which suggests an important quality control function for calreticulin.
Calreticulin does not prevent the export of suboptimally loaded MHC class I molecules, but can transit with these molecules to the Golgi apparatus and retrieve these molecules back to the ER via its own KDEL retrieval sequence (63). Whether the retrieved MHC molecules are then destined for degradation or further optimization remains to be determined. Such events would perhaps require alterations in the redox status of MHC class I molecules, either by reduction of the heavy chain in preparation for ERAD (145) or the reduction of the C101-C164 bond, to perhaps allow for more efficient peptide reloading. Thus, the redox status of MHC class I heavy chains could potentially influence the activity of calreticulin through the recruitment of ERp57.
PDI and the PLC
PDI, the archetypal member of the PDI family of oxidoreductases and one of the most abundant within the ER, can reduce, oxidize, and isomerize disulfide bonds via two CGHC reactive motifs. PDI was proposed to be a hub for MHC binding peptides and to allow both early and final oxidative folding events to take place (72, 113). Furthermore, PDI has been proposed to regulate release of MHC class I-peptide complexes from the ER by mediating the dissociation of the ERp57-tapasin conjugate (83). Support for a role of PDI in HLA-B27 assembly has been implicated by the detection of PDI in complexes containing HLA-B27 heavy chain, ERp57, and tapasin (125). We have failed to find PDI associated with HLA-B27 at the early folding stages (unpublished); however, PDI can be coimmunoprecipitated with partially unfolded HLA-B27 molecules (31).
Overall, the folding of MHC class I molecules can be subdivided into two distinct phases, which are either associated with distinct chaperones such as CNX and tapasin during the early and latter folding stages, respectively, or overlapping molecules such as the oxidoreductases ERp57 and PDI. Furthermore, peptide binding of MHC class I molecules can occur in two stages, which may reflect the two phases of assembly. The early folding process is governed by the binding of low affinity peptides, whereas the latter stage involves the optimization and acquisition of higher affinity peptides. Intriguingly, although full oxidation can occur before association with β2m, binding of β2m is thought to enhance the efficiency of disulfide bond formation (143). It remains uncertain and somewhat controversial whether the C101-C164 bond remains partially reduced/oxidized upon entry into the PLC. However, full oxidation of MHC class I heavy chains occur in two stages: first, the C203-C259 bond within the α3 domain followed by the C101-C164 bond within the α2 domain (142, 143). These two distinct stages of oxidation could well coincide with the early and latter stages of folding. With respect to HLA-B27, it remains undetermined how the unpaired cys residues impact on the oxidation of the heavy chain, especially C67. Our studies demonstrating that HLA-B27 has a slow maturation phenotype, exists in different oxidation states, and maintains strong associations with ERp57 could well reflect the process of resolving disulfide bonds, probably via a series of reducing and isomerizing reactions. Such potential redox reactions may contribute to the slow folding kinetics and enhanced propensity of HLA-B27 to misfold (7, 46).
HLA-B27 Heavy Chain Dimerization
One unusual biochemical characteristic of HLA-B27 directly related to its redox status and that has received much attention in recent years has been its enhanced ability to form heavy chain-dimers (2, 7, 36, 100) (Fig. 5). This has revived interest in the redox chemistry of HLA-B27 (13, 46, 99, 154). However, though dimerization is not a common event, both mouse and human MHC class I products have been shown to dimerize (20, 28, 35).

HLA-B27 dimerization was originally noted when purifying recombinant HLA-B27 heavy chains in vitro (2). These structures were subsequently detected in in vivo settings (7, 36, 100). With cell types delineating from the myelomonocytic lineage appearing to be important in disease development in the transgenic HLA-B27 rat model (21, 22), it was intriguing that immature dendritic cells expressing HLA-B27 did not have detectable dimers, but maturation lead to substantial dimer formation (126). HLA-B27 transgenic rats are perhaps the best animal model for the SpAs as they develop a spontaneous disease recapitulating many facets of the human disease, including both skeletal and gut disease (55, 141, 146), with dimer levels correlating with disease incidence in HLA-B27 transgenic rats (147). However, when misfolding was alleviated by the introduction of higher levels of human β2m, as determined by heavy chain-dimer formation, these animals still developed skeletal disease but did not develop the accompanying gut inflammation (146). Thus, observations in the rat model indicate that HLA-B27 misfolding and possibly the generation of heavy chain-dimers do play a role in SpA development with misfolding playing a more significant role in gut inflammation.
ER-resident dimerization
The precise mechanism of HLA-B27 heavy chain-dimerization remains undetermined. The unpaired C67 was originally proposed to participate in dimer formation, as mutating C67 prevented recombinant HLA-B27 heavy chains from dimerizing (2) (Fig. 5). However, our studies and those of others revealed that in the absence of C67, though HLA-B27 heavy chain-dimerization was lower, HLA-B27 maintained an ability to dimerize (7, 147) (Fig. 5). Transgenic rats expressing HLA-B27.C67A were found to still express dimers and develop inflammatory arthritic disease but with a lower prevalence (141, 146). Other HLA alleles also possess a C67, which include HLA-B14, B15, B38, B39, and B73 (77), and, interestingly, HLA-B14 exhibits an association with the SpAs and B39 with autoimmune-mediated diabetes (see Table 1) (101, 102, 105).
The formation of heavy chain-dimers in the absence of C67 suggested that either other unpaired cysteines at p308 and/or p325 or structurally conserved cys residues could participate in disulfide bonding. Our studies have revealed that the structurally conserved C164 plays a significant role in dimer formation (7) (Fig. 5). Why do cys residues other than C67 participate in disulfide bond formation? During disulfide bond formation, cys residues must be both exposed and reactive. As discussed previously, the disulfide bond between C101-C164 is thought to form after the C203-C259 disulfide bond and at a latter stage of the MHC class I folding process (38). Thus, C164 could remain exposed to the oxidizing environment of the ER for a prolonged period, allowing it to potentially form heavy chain-dimers. However, as this bond is thought to form following appropriate peptide loading, this would leave C67 exposed for prolonged periods, thus allowing it to also participate in ER-resident dimer formation. It was proposed that the positively charged lysine residue at p70 of HLA-B27 heavy chain contributed to the reactivity of C67 (154). However, mutagenesis studies revealed that several residues encompassing p65-71 and those that make up the B pocket also contribute to C67 participation in disulfide bond formation either by increasing its reactivity and/or its accessibility (7, 100).
The application of a rapid acidification and alkylation method, which has been used to monitor protein topology and redox status of oxidoreductases, revealed that HLA-B27 can exist in at least three different conformations in an in vivo setting (46, 118) (Fig. 5). To date our studies have yet to reveal similar conformations with other alleles. The cys residues of HLA-B27 are indeed exposed at steady state levels, which could give rise to multiple conformations and potential precursors to heavy chain-dimer formation. The X-ray crystallographic structure of HLA-B27 did not reveal any such altered conformations (93); however, these studies are biased toward crystallization of stable peptide-bound structures. Recent structural analysis has revealed that HLA-B*2705 can bind the same peptide in different orientations (65) because of the enhanced flexibility of its antigen binding groove (44). This inherent flexibility may allow the temporary binding of a larger repertoire of low affinity peptides or lead to enhanced propensity to relax, partially unfolding and thus lose peptide bound conformations.
Dimers within the ER have been detected that are reactive with different antibodies, namely, HC10 and W6/32 (36). These antibodies are specific for different conformations of MHC class I molecules. W6/32 can detect fully folded MHC class I molecules, whereas HC10 can detect partially folded/unfolded HLA-B and C alleles (137). It is possible that the W6/32 reactive heavy chain-dimers could represent cell surface dimers (Fig. 5). However, these observations raise the possibility that there are several populations of heavy chain-dimers within the ER (Fig. 5).
Expression of HLA-B27 in cell lines deficient for components of the PLC have resulted in diminished or enhanced dimer formation. In the absence of TAP, heavy chain-dimers are not so abundant. This may reflect that populations of dimers are either peptide dependent or the lack of peptide could allow for more efficient degradation. However, in the absence of tapasin, heavy chain-dimerization is enhanced. This may be due to tapasin or ERp57 reactivity imparting some quality control process that would otherwise eliminate dimers. Furthermore, the absence of β2m also allowed for enhanced dimerization, but only of the unfolded population as determined by HC10 reactivity (31).
Dimerization is therefore not a simple action of two heavy chains undergoing redox coupling, but could be regulated by chaperones, especially oxidoreductases or the overall oxidizing environment of the ER. HLA-B27 is strongly associated with the oxidoreductase ERp57 (Fig. 3), but other oxidoreductases could participate in dimer formation such as PDI. The heterogenous populations of heavy chain-dimers observed may arise through distinct inter- and intramolecular disulfide bonds between different cys residues, varying numbers of intermolecular disulfide bonds or dimers forming between HLA-B27 heavy chains that have adopted different conformations (Fig. 5). The latter possibility is supported by the detection of dimers by the W6/32 antibody (36); thus, HLA-B27 dimers can occur between not only partially unfolded heavy chains but also heavy chains that have adopted a more fully folded conformation.
Cell surface heavy chain dimers
HLA-B27 dimers have been detected at the cell surface (7, 17, 76) (Fig. 5). Heavy chain-dimers that form within the ER do not appear to be directly related to those formed at the cell surface. ER-resident dimers have not been shown to egress from the ER to the cell surface via the secretory pathway. Instead, heavy chain-dimer formation at the cell surface is dependent on endocytic trafficking and C67 (17) (Fig. 5). The acidic environment through which cell surface molecules recycle could dissociate or instigate unfolding events, exposing C67, which may become more reactive due to the acidic environment, thus enhancing dimerization. Alternatively, the potential protonation of free thiols in an acidic environment could trap heavy chain–heavy chain structures that would otherwise fall apart or become targeted to the lysosomal machinery for degradation (Fig. 5). However, methodology employed in detecting these structures may influence their formation, as techniques such as cell surface iodination is highly oxidizing and the use of different alkylating agents can affect levels of cell surface dimers detected (7). The recycling of HLA-B27 through the endocytic pathway could expose partially unfolded HLA-B27 to oxidoreductases at the cell surface, which could subsequently catalyze the dimerization process. Oxidoreductases such as PDI, ERp57, ERp44, and thioredoxin can be detected at the cell surface, especially following cellular activation or oxidative stress (62, 67, 127).
One intriguing aspect of HLA-B27 cell surface expression is the detection of partially folded/misfolded molecules by the HC10 antibody. It remains undetermined whether the HC10 reactive heavy chain population; (i) has undergone redox changes, (ii) contains precursors for heavy chain-dimer formation, and (iii) contains actual cell surface dimers. These partially folded/unfolded cell surface HLA-B27 molecules can be detected in association with BiP (7). However, such cell surface chaperone associations may not be totally unusual, as partially folded/unfolded MHC molecules can be found at the cell surface of activated cells in association with ER-resident chaperones (127). Other unusual HLA-B27 cell surface structures have been reported. For instance, a population of HLA-B27 heavy chain molecules bound to extended peptides in the absence of β2m can be detected by the MARB4 monoclonal antibody (95). Whether these latter structures are dependent on redox changes and/or status remains to be determined.
The presence of cell surface dimers and aberrant HLA-B27 cell surface molecule expression has important implications for immune recognition and potentially for the development of the SpAs. HLA-B27 dimeric structures have been detected on the cell surface of peripheral blood mononuclear cells (76). It has been postulated that changes in redox status of HLA-B27 in SpA patients can influence antigen recognition by the T cell receptor of cytotoxic CD8+ T cells (12). However, in the transgenic rat model CD8+ cytotoxic T cells are believed to play a minimal role in disease onset (140).
HLA-B27 is not only recognized by the cognate T cell receptor of cytotoxic CD8+ T-lymphocytes but by other immunomodulatory receptors (3). MHC class I molecules are ligands for NK cells, which are sensitive to MHC class I levels. NK cells can express killer cell immunoglobulin-like receptors (KIR) and leukocyte immunoglobulin-like transcripts (ILT). KIRs can also be found on T and NKT cells (78, 79) and ILTs can be found not only on T and NK cells, but also B cells, monocytes, macrophages, and dendritic cells (32). KIRs are polymorphic and can recognize different MHC class I molecules. HLA-B27 appears to be predominantly recognized by KIR3DL1. ILTs have a broader specificity, with ILT4 and ILT2 recognizing most HLA-B alleles (32, 33). Using an elegant HLA-B27 tetrameric molecule that detects heavy chain-dimer reactive ligands, Kollnberger et al. were able to demonstrate that heavy chain-dimers could be recognized by KIR3DL1 and 2, whereas these structures were also recognized by ILT4 but not ILT2 (76). Such interactions could have important implications for immune cell function and thus disease. The ILT4 interaction is influenced by specific peptides bound by HLA-B27; interestingly, the transmembrane and cytosolic unpaired cys residues were shown to impact on the recognition of HLA-B7 by leukocyte inhibitory receptor 1 (52). Therefore, B alleles may adopt unusual structures required for recognition by ILT/leukocyte inhibitory receptors. An intriguing and novel HIV escape mechanism involves mutations in an HLA-B27 gag epitope, which seems to be directed toward the interaction of the HLA-B27-gag peptide complex with ILT4 (86). This altered interaction is believed to lead to poor maturation of myelomonocytic cells and development of a more tolerogenic phenotype. Thus, the effect of dimeric structures on the immunobiology of myelomonocytic cells may have interesting implications for the SpAs.
Distinct Roles for Unpaired Cysteine Residues
The additional unpaired cys residues found within HLA-B27 and indeed other HLA molecules raise the question why do MHC class I molecules possess these residues? Do these extra cys participate in specific functions?
We have already discussed a possible function for C308 in PLC formation (29) (Fig. 4) and the potential impact that C308 and C325 may have on immune recognition (52). C67 was noted to confer stability to cell surface HLA-B27 molecules (4); thus, these additional cys residues may participate in the actual folding of HLA-B27 and other HLA-B alleles. Such a scenario was supported by the demonstration that C308 and C325 can participate in the folding of HLA-B7, with C308 acting as a palmitoylation site (52). However, the underlying basis for this phenomenon remains unresolved, especially when compared to other HLA class I molecules that do not possess these residues.
Recently, C325 of HLA-B27 was demonstrated to be involved in heavy chain-dimer formation, but in an entirely novel context (89) (Fig. 5). Dimers were found within exosomes, small vesicles that form by the inward budding of endosomes to generate multivesicular bodies, which can fuse with the plasma membrane releasing these internal vesicles/exosomes (70, 120). Interestingly heavy chain-dimers were not only composed of HLA-B27 homodimers but could also be found as HLA-B27-HLA-A2 heterodimers. HLA class I dimers could therefore be heterologous in nature. HLA-A2 can also be found as part of the haplotype that is associated with the SpAs, which may indicate a specific role for heterodimers in some aspects of disease (139).
From the above studies it is clear that dimer formation can occur within different cellular compartments (Fig. 5), with the cys residues involved differing depending on the compartment and probably exposure to the different redox environments. The dimers detected within exosomes were composed of fully folded molecules and their formation mediated via the cytoplasmic domain C325. This would appear to make biological sense, as C325 would be exposed to the more oxidizing environment within the exosome, due to the depletion of reduced glutathione (Fig. 5). Thus, different environments can have a profound effect on which cys residues participate in dimer formation (Fig. 5).
Not all subtypes of HLA-B27 are associated with the SpAs, and this is especially true for HLA-B*2706 and 09 (119). It has been postulated that different subtypes, such as HLA-B*2705 and 04, have differing redox activities (124). Our studies revealed that the different subtypes exhibited differential accessibility to dithiothreitol, revealing that these subtypes can exist in different redox states (46). Furthermore, the 114–116 region has been demonstrated to influence heavy chain-dimerization (19). As the predominant polymorphisms of HLA-B*2706 and 09 are found within the 114–116 region, which constitutes part of the F pocket, this again implicates peptide binding as the ultimate determining factor in redox status and dimerization.
The Influence of Redox on ERAD
Misfolded proteins within the ER are targeted, removed, and degraded within the cytosol by the cytosolic 26S proteasome in a series of regulated events collectively known as ERAD (98, 152). MHC class I molecules seem to be targeted for degradation via two distinct pathways of ERAD. This has been best illustrated by the activity of two immunoevasion proteins encoded by the human cytomegalovirus, US2 and US11. These two proteins seem to select different MHC class I substrates and thus identify two distinct ER degradation pathways (9).
The pathway used by US11 identified the Derlin-1-SEL1L complex comprising of derlin-1, which is a candidate translocation channel, the ER membrane integrated E3 ubiquitin ligase HRD1, and SEL1L. It also includes ER-resident targeting factors, which include ER degradation-enhancing α-mannosidase-like lectins (EDEMs) and lectins containing mannose-6-phosphate receptor-like domains OS9 and XTP3-B, valosin-containing protein-interacting membrane protein, homoCys-responsive ER-resident protein, and the AAA ATPase p97, which provides the driving force for extraction of misfolded polyubiquitinated proteins. Interestingly, this pathway was shown to be dependent on the redox status of MHC class I heavy chains, with their reduction a prerequisite before dislocation and degradation (145). Prior reduction of ERAD-targeted MHC class I heavy chains could well be mediated by the recently described transmembrane thioredoxin-related protein, which is thought to act in concert with CNX (97). The degradation of misfolded MHC class I heavy chains would be particularly sensitive to the Derlin-1-SEL1L degradation pathway. Misfolded protein substrates associated with CNX seem to rely on the Derlin-1-SEL1L degradation complex, with EDEM playing a vital role in handing over substrates from CNX to this complex for degradation (103, 108). EDEM was initially described as a gene that was activated by the UPR and was thought to possess mannosidase activity. Recently, it was demonstrated that EDEM can bind proteins independently of the sugar moiety and form a key interaction with SEL1, which modulates ubiquitination of protein substrates via the E3 ubiquitin ligase HRD1 (34). Recently, HRD1, together with the E2 ubiquitin-conjugating enzyme UBE2J1, was shown to ubiquitinate and dislocate misfolded MHC class I heavy chains from the ER. Interestingly, the absence of HRD1 resulted in the accumulation of HLA-B27 dimers (26).
The US2 protein exploits a different ERAD pathway that is less well characterized. US2 can target more folded MHC molecules (51, 92, 144) as suggested by the crystal structure of US2-HLA-β2m complex (50). The degradation complex is Derlin-1 independent and requires the activity of the signal peptide peptidase, which is a presenilin-like aspartyl protease capable of cleaving proteins within membranes (88). Using a library of siRNAs targeting ubiquitin E3 ligases, the E3 ligase translocation in renal carcinoma, chromosome 8 gene was identified to be part of this pathway. Furthermore, it was found to associate with both MHC class I heavy chain and signal peptide peptidase (136).
One of the key biological anomalies of HLA-B27 is the prolonged half-life of the immature heavy chain, which does not appear to be degraded efficiently. The existence of different HLA-B27 conformers with different redox states could influence their degradation rates. The susceptibility of these different conformations to ERAD has yet to be determined, but these populations could contribute to the enhanced detection of immature/misfolded protein. Furthermore, disulfide bonded heavy chain-dimeric structures could be less susceptible to degradation, especially if these structures are insensitive to reductases.
Two interesting observations may point to how aberrant heavy chain-dimeric structures may be reduced before degradation. We have noted that ERp57 predominantly associates with HLA-B27 heavy chains (Fig. 3), possibly reflecting the process of isomerization and reduction that is required before degradation (5, 7). This is supported by observations that ERp57 can preferentially reduce partially folded and inappropriately peptide-loaded MHC class I molecules (5). The prolonged association between ERp57 and HLA-B27 heavy chain could reflect that HLA-B27 can exist in several different redox states that maybe difficult to resolve.
In addition, the prolonged interaction between BiP and HLA-B27 may be required for its degradation (7) (Fig. 3). One of the BiP cochaperones ERdj5 has reductase activity, which participates in the degradation of protein substrates (150). ERdj5 works in concert with BiP and EDEM and has a low standard reduction potential of −219 mV, which indicates that its major role is reduction of client proteins (14, 150) potentially acting as an intersection with the Derlin1/p97 pathway. Interestingly, we have found that BiP exhibits a prolonged interaction with both HLA-B27 monomers and heavy chain-dimers, indicating that this pathway may be important in their degradation (unpublished observations). Whether ERdj5 is part of the complex remains to be determined. However, this prolonged association would mirror other complexes identified containing BiP and misfolding proteins (60, 135).
Oxidative Stress: Potential Consequences for HLA-B27
We have discussed how different cellular environments will have varying oxidative conditions, which could impact on cysteine residue reactivity and dimerization. However, the oxidative microenvironment of cellular compartments can be altered through various biological processes especially after infection and cellular stress.
Oxidative stress, through the production of reactive oxygen radicals, can have damaging effects on host proteins. Recently, oxidative stress, viral infection, and toll receptor engagement were demonstrated to lead to enhanced misacylation of tRNAs and it was proposed that host cells enhance methionine incorporation to protect proteins against oxidative stress (106). Such conditions could alter the amino acid composition of HLA-B27, thus potentially altering its folding dynamics. However, if reactive oxygen species trigger such translational changes, these may well be cell type specific. For instance, dendritic cells were demonstrated to control the production of reactive oxygen species within phagolysosomes by recruiting the NADPH oxidase NOX2 to reduce the proteolytic activity within these compartments, thus protecting antigenic peptides from complete degradation. However, such a buffering mechanism appears to be absent from macrophages (128). It has been postulated that unfolded proteins could lead to oxidative stress (94), which could in part explain why HLA-B27 expressing macrophages are more susceptible to UPR (149). Thus, the UPR, cell type, and oxidative stress could well contribute to the unusual biology of HLA-B27 and its pathogenesis.
Expression of three unpaired cysteines within the HLA-B27 heavy chain is likely to contribute to adverse folding events, which are active within different cellular locations (Fig. 5). Within the ER, the oxidizing conditions and folding events will expose C67, C101, and C164 to potential interactions leading to dimerization (Fig. 5). Due to the highly reducing environment of the cytosol, the cytoplasmic tail domain C325 does not normally participate in dimerization, but within the exosome lumen, conditions are favorable for C325-C325 interactions. Recycling of cell surface HLA-B27 could potentially enhance dimerization, predominantly through C67, aided by accompanying unfolding events (Fig. 5).
HLA-B27: Summary and Future Directions
The biosynthesis of HLA-B27 appears to be affected by redox conditions of local cellular environments, which would influence cysteine residue reactivity and chaperone involvement. The slow maturation of the HLA-B27 heavy chain and enhanced exposure of cysteine residues are contributory factors to HLA-B27 dimerization. The in vivo mechanism of dimerization remains undetermined and the association with the chaperones CNX, BiP, and the oxidoreductase ERp57 suggest that dimerization could be a chaperone-mediated event. These chaperones may exhibit enhanced associations with HLA-B27 to improve the folding of a heavy chain that due to the three unpaired cysteines at p67, 308, and 325 has inherent problems acquiring the appropriate conformation. Alternatively, these chaperones could function in the ERAD process and their enhanced associations with HLA-B27 could reflect a block in the disposal pathway. Determining the role of these chaperones in the biosynthesis and degradation of HLA-B27 will not only shed light on how and why HLA-B27 misfolds and dimerizes, but also will reveal their function in the MHC class I pathway.
Evidence indicates that misfolding events can contribute to inflammatory arthritic disease, and thus intervention strategies manipulating the redox status could be used as potential therapeutics. The importance of HLA-B27 heavy chain dimers in the development of the SpAs and whether these structures can be used as potential SpA disease markers remains undetermined. A key question is whether the misfolding of the monomeric heavy chain or the formation of dimers is a pathogenic event, and if so, does this lead to the induction of ER stress responses, which can lead to proinflammatory cytokine production. If ER stress is key to understanding the pathogenesis of the SpAs, this would have implications in how we categorize and treat these important group of diseases.
Footnotes
Acknowledgments
Antony N. Antoniou is supported by an Arthritis Research UK (ARUK) Fellowship (15293). David Guiliano is supported by an ARUK project grant (17222). Izabela Lenart is supported by ARUK studentship (17868).