
Abstract
Cells must be able to maintain their intracellular homeostasis in the face of changing conditions. Typically, they respond by invoking complex regulatory mechanisms, including a global inhibition of translation. This reduction in protein synthesis may prevent continued gene expression during potentially error-prone conditions as well as allow for the turnover of existing mRNAs and proteins, whilst gene expression is directed toward the production of new molecules required to protect against or detoxify the stress. However, it is becoming increasingly recognized that not all translation is inhibited and translational control of specific mRNAs is required for survival under stress conditions. Control of protein levels via translational regulation offers a significant advantage to the cell due to the immediacy of the regulatory effect. This review describes how protein synthesis is regulated in response to oxidative stress conditions induced by exposure to hydrogen peroxide. Translational control can be mediated via direct oxidative regulation of translation factors as well via mRNA-specific regulatory mechanisms. Additionally, increasing evidence suggests that oxidative damage to the translational apparatus can itself alter the proteomic output. The resulting translational reprogramming is fundamental for adaptation to the oxidative stress. Antioxid. Redox Signal. 15, 191–203.
Introduction
Oxidative stress can potentially affect the protein synthetic apparatus via several mechanisms. For example, increasing evidence suggests that the translation apparatus is itself a primary target of oxidative damage and this can have profound effects on the translational output. Alterations in the translational output can also occur via oxidative regulation of specific translation factors that can either down- or upregulate the bulk rate of protein synthesis. Additionally, oxidative stress can regulate translation in an mRNA-specific manner, which is important when the resultant reprogramming of gene expression is required for adaptation to the stress condition. This review describes how oxidative stress induced by exposure to H2O2 affects translation and, in particular, the consequences for the cellular proteome and stress tolerance.
Regulation of Translation Factors in Response to Oxidative Stress
Translation initiation is a key regulated step in gene expression
The initiation phase of protein synthesis is the main target of translational regulation and represents a key control point for eukaryotic gene expression. It is a complex process involving at least 12 eukaryotic initiation factors (eIFs) interacting with ribosomal subunits, initiator methionyl-tRNA (Met-tRNAi met), and mRNAs. Translation initiation has been extensively reviewed (41, 45, 81), and only an overview is provided here. Initiation requires the formation of a 43S preinitiation complex, joining with the 60S ribosomal subunit and correct positioning of an 80S ribosome at the AUG codon. It is triggered by the formation of the ternary complex (TC), which consists of eIF2, guanosine 5′ triphosphate (GTP), and Met-tRNAi met. eIF2 is a heterotrimer (α, β, and γ subunits) that plays a key role in regulating initiation through its interaction with eIF2B. After each round of initiation, eIF2 is released from the ribosome as a binary complex with GDP. GDP is removed and replaced by GTP in a guanine-nucleotide exchange reaction catalyzed by eIF2B. Met-tRNAi met can only bind the eIF2/GTP complex, so translational control can be regulated by the activity of eIF2B (Fig. 1). The TC binds to the small (40S) ribosomal subunit, aided by eIF1, eIF1A, and eIF3, and the resulting 43S preinitiation complex binds mRNAs. The eIF4F cap-binding complex, comprising eIF4E, eIF4G, and eIF4A, functions in the recruitment of ribosomes to mRNAs. This is accomplished through recognition and binding to the 5′cap structure by eIF4E and by the ATP-dependent unwinding of secondary structures in the mRNA 5′ region by eIF4A. Binding of eIF4E to the 5′cap is a key regulated step in translation initiation. The 43S complex scans the mRNA until it encounters an AUG codon. This requires ATP-dependent unwinding of secondary structures and linear movement along mRNAs. Initiation codon recognition and base pairing is followed by hydrolysis of eIF2-GTP, mediated by the GTPase activating activity of eIF5. Joining of the 60S ribosomal subunit and initiation factor dissociation results in the assembly of an 80S ribosome that is competent to begin translation elongation.
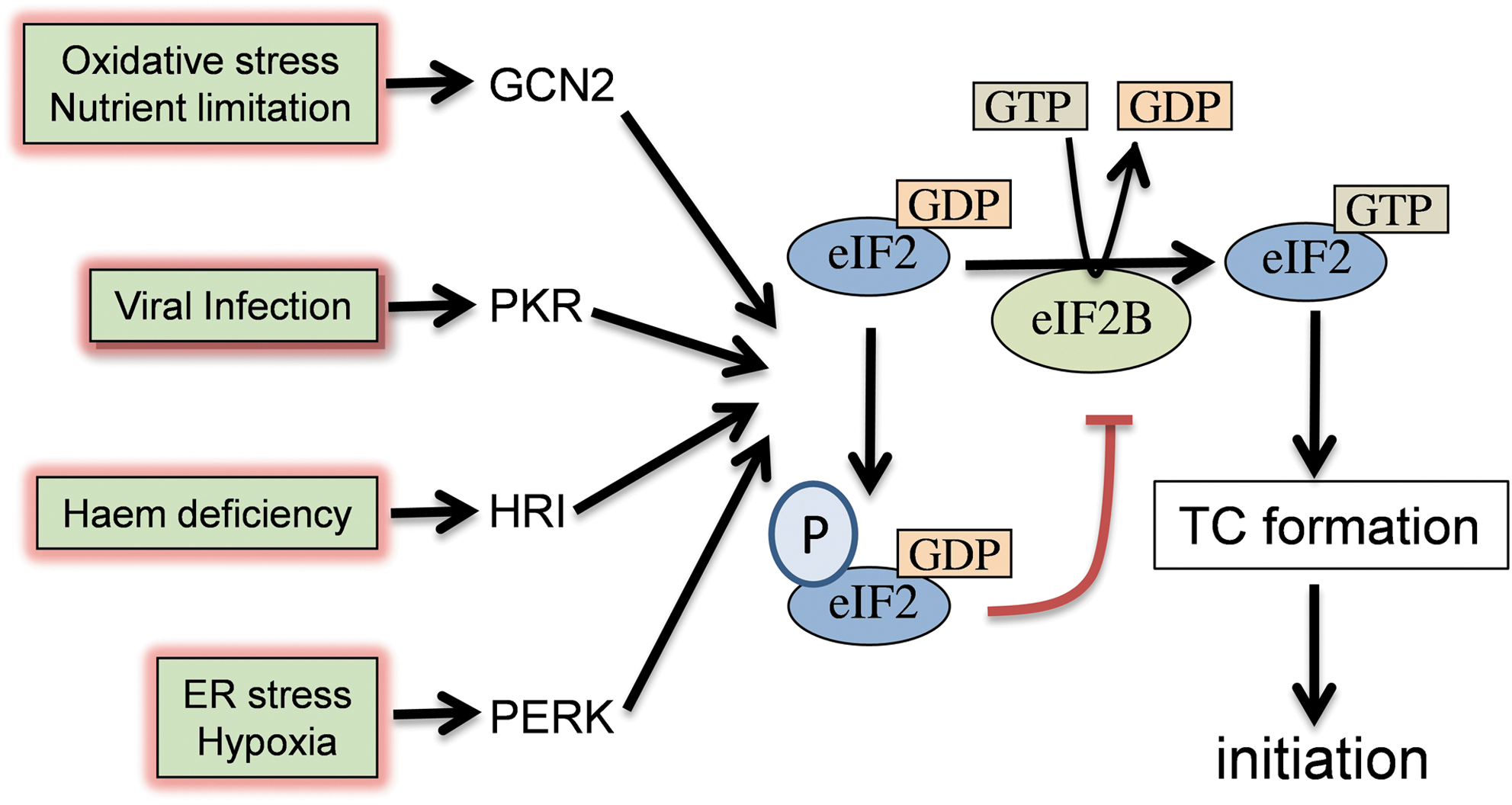
Regulation of TC formation
Phosphorylation of eIF2 is a key regulated step in translation initiation (Fig. 1). In eukaryotic cells this is achieved by phosphorylation of the alpha subunit of eIF2 at a conserved serine (Ser51) residue (33, 79). Phosphorylation converts eIF2 from a substrate to a competitive inhibitor of the guanine nucleotide exchange factor eIF2B and the resulting decrease in eIF2B activity leads to reduced TC levels, which inhibits translation initiation (37). Four mammalian kinases have been identified that inhibit translation initiation by phosphorylating eIF2α. GCN2 (the amino acid control kinase), PKR (the double-stranded RNA-activated protein kinase), HRI (the heme-regulated inhibitor), and PERK/PEK (the PKR-like endoplasmic reticulum eIF2α kinase) are regulated independently in response to different cellular stresses (19, 83). For example, PERK has been found in all multicellular eukaryotes and is a component of the unfolded protein response (UPR). Consistent with its central role in the endoplasmic reticulum (ER) stress response, cells lacking PERK fail to phosphorylate eIF2α and do not downregulate protein synthesis during ER stress conditions (7). Attenuating protein synthesis may act to reduce the burden of newly synthesized ER client proteins on the ER folding machinery. However, phosphorylation of eIF2α not only represses general translation, but it also induces translation of specific mRNAs, such as that encoding the metazoan activating transcription factor 4 (ATF4) (57, 111). ATF4 mediates the integrated stress response whose targets include genes that encode proteins involved in amino acid metabolism and resistance to oxidative stress, ultimately protecting against the deleterious consequences of ER oxidation (34). Genetically, uncoupling stress activation of PERK has been used to show that PERK-mediated phosphorylation of eIF2 preconditions cells to oxidative and ER stress conditions through enhanced expression of antioxidant and stress protective molecules (58). Similarly, increasing the phosphorylation of eIF2α by using the phosphatase inhibitor, salubrinal, increases oxidative stress tolerance via ATF4 activation (53). ATF4-mediated responses to oxidative stress are discussed in more detail under mRNA-specific translational control mechanisms.
Recent studies indicate that moderating protein synthesis in response to ROS through eIF2α phosphorylation is a common mechanism (83). The fission yeast Schizosaccharomyces pombe has fewer eIF2 kinases than mammalian cells, containing only Gcn2 and two HRI-related protein kinases called Hri1 and Hri2 (118). Hri2 is the primary kinase that is activated in response to heat shock, arsenite, or cadmium stress, whereas Gcn2 responds to nutrient downshift, amino acid starvation, and osmotic stress. Both Hri2 and Gcn2 are activated and phosphorylate eIF2α in response to oxidative stress conditions induced by exposure to H2O2. The role of these eIF2 kinases during oxidative stress conditions is further confirmed by the finding that the growth of a mutant lacking GCN2 and HRI2 is sensitive to H2O2 (118). More recently, phosphorylation of eIF2α and inhibition of protein synthesis in fission yeast has been shown to be under the dual control of the stress-activated protein kinase (SAPK) pathway and eIF2 kinases (6, 23). In the yeast Saccharomyces cerevisiae, Gcn2 is the sole eIF2 kinase and was originally shown to phosphorylate eIF2α in response to amino acid starvation (19). Phosphorylation of eIF2α by Gcn2 reduces global protein synthesis, but paradoxically, translation of the GCN4 mRNA is activated in response to low TC levels in a mechanism involving four short upstream open-reading frames (uORFs) (38). Gcn4 is a transcription factor that activates gene expression of many targets, including amino acid biosynthetic genes (70). Thus, analogous to the mammalian integrated stress response, activation of Gcn4 serves to overcome the imposed starvation, which initially led to the translational control (Gcn4-mediated responses to oxidative stress are discussed further under mRNA-specific translational control mechanisms). H2O2 activates Gcn2-phosphorylation of eIF2α and inhibits translation initiation in a dose-dependent manner (93). This also causes upregulation of GCN4 expression and Gcn4 is required for H2O2 tolerance (60). Regulating translation initiation via phosphorylation of eIF2α appears therefore to be a common response mechanism to ROS in eukaryotic cells.
The exact mechanism by which oxidative stress activates eIF2 kinases in eukaryotic cells is not known. It appears to be a general response to ROS, since yeast Gcn2 can be activated by exposure to an organic hydroperoxide (cumene hydroperoxide), a thiol oxidant (diamide), and a heavy metal (cadmium) (60). Yeast Gcn2 is now known to be activated in response to a variety of conditions, including nutrient starvation (amino acids, purines, and glucose) and exposure to sodium chloride, rapamycin, ethanol, and volatile anesthetics (38, 76). Depletion of amino acids leads to an accumulation of uncharged tRNA, which activates Gcn2 via its histidinyl-tRNA synthetase (HisRS)-related domain. It is likely that other stress conditions ultimately influence the levels of uncharged tRNAs in the cell. For example, volatile anesthetics inhibit amino acid uptake (76) and Gcn2 is activated by glucose starvation partly through an effect on vacuolar amino acid pools (117). The signals activating Gcn2 in response to rapamycin or NaCl are not well understood. Rapamycin appears to work by blocking Tor-mediated phosphorylation of Gcn2 at Ser577 (38). However, activation of Gcn2 by rapamycin and NaCl still requires the HisRS-related domain of Gcn2, as well as Gcn1 and Gcn20, which are thought to mediate the activation of Gcn2 by uncharged tRNA (69). Similarly, the inhibition of translation initiation in response to H2O2 requires Gcn1 and Gcn20 (93). Oxidative stress conditions appear to activate Gcn2 via an effect on uncharged tRNA levels since mutations in the HisRS-like domain abolish H2O2 activation of Gcn2. H2O2 activates Gcn2 in a prototrophic yeast strain (a strain that does not require exogenous amino acids from the growth media) ruling out any effects on amino acid uptake (60). This raises the question as to how ROS cause an amino acid starvation and/or affect cellular uncharged tRNA levels. Oxidative stress may conceivably cause an accumulation of uncharged tRNA through a variety of mechanisms. Free amino acids and amino acids in proteins are highly susceptible to oxidation by ROS (98). Oxidized amino acids can be detected in yeast cells; for example, oxidized phenylalanine (m- and o-Tyr) is elevated after exposure to similar concentrations of H2O2 as those that activate Gcn2 (80). Although the levels of oxidized amino acids detected in yeast cells exposed to H2O2 are relatively low, representing about one modification per 103 phenylalanine residues, the total oxidative load on the amino acid pool may trigger an amino acid starvation response. Alternatively, the proteins and nucleic acids that are required for tRNA-aminoacylation may be susceptible to oxidation resulting in an accumulation of uncharged tRNA and activation of Gcn2. For example, oxidative damage to RNAs, including tRNAs, has been implicated in the pathogenesis of Alzheimer's disease (21, 39) and ROS can affect the stability and activity of aminoacyl-tRNA synthetases (100).
Regulation of mRNA binding by eIF4E
A key control point for regulating eukaryotic translation initiation is via binding of initiation factors to the mRNA cap (Fig. 2). The initiation factor eIF4E recognizes the mRNA cap structure and also interacts with the multiadaptor protein eIF4G (87). This process is critical for the recruitment of ribosomes to the 5′ end of mRNAs, and translation initiation can be regulated by the competitive binding of three mammalian eIF4E-binding proteins (4EBP1, 2, and 3) to eIF4E (86). Hypoxia (oxygen deprivation) inhibits protein synthesis via effects on both translation initiation and elongation (55). Hypoxia appears to increase the generation of ROS, most prominently H2O2, from mitochondria (56). This is important physiologically since hypoxia can occur in many pathological conditions, including tumor formation and inflammation. The inhibition of translation initiation occurs via PERK-dependent phosphorylation of eIF2α and regulation of 4EBP1 binding to eIF4E. 4EBP1 is phosphorylated by the mammalian target of rapamycin complex 1 (mTORC1), which prevents its binding to eIF4E and so releases active eIF4E to initiate translation (Fig. 2). Hypoxia inhibits mTORC1, decreasing 4EBP1 phosphorylation. Hypophosporylated 4EBP1 binds eIF4E and causes inhibition of cap-dependent translation initiation. Similarly, H2O2 exposure has been shown to inhibit mTORC1 phosphorylation of 4EBP1, which is associated with cell death in neuronal cells (13). Multiple 4E-BPs (e.g., Maskin, Bicoid, and Cup) have also now been described, which provide mRNA specificity through additional protein–RNA interactions that target the 4E-BP to individual mRNAs (87). Few examples have been studied in detail, but they have been proposed to promote mRNA-specific translational control during diverse growth and stress conditions. It is tempting to speculate that many of these growth conditions will involve ROS generation, and hence oxidant regulation of mRNA translation by 4E-BPs is likely to provide a research area of increasing interest.

Two eIF4E-binding proteins have been identified in yeast, Caf20 and Eap1 (1, 16). These 4EBPs have broad functions in cell growth, proliferation, and development (40, 77). Eap1 (but not Caf20) is required to inhibit translation initiation in response to oxidative stress induced by cadmium and diamide exposure. In contrast, peroxide-induced inhibition does not appear to require Eap1, highlighting the different translational control mechanisms invoked by these different oxidants (60). Yeast cells respond to a membrane stress by attenuating translation initiation via a mechanism that is mediated by Gcn2 and Eap1 (18). This response may serve to prevent the mislocalization of proteins after disruption of vesicular transport pathways during ER stress conditions. This may be important since yeast cells do not contain a PERK homolog that can regulate translation initiation via eIF2α phosphorylation during ER stress conditions.
Regulation of translation elongation during oxidative stress conditions
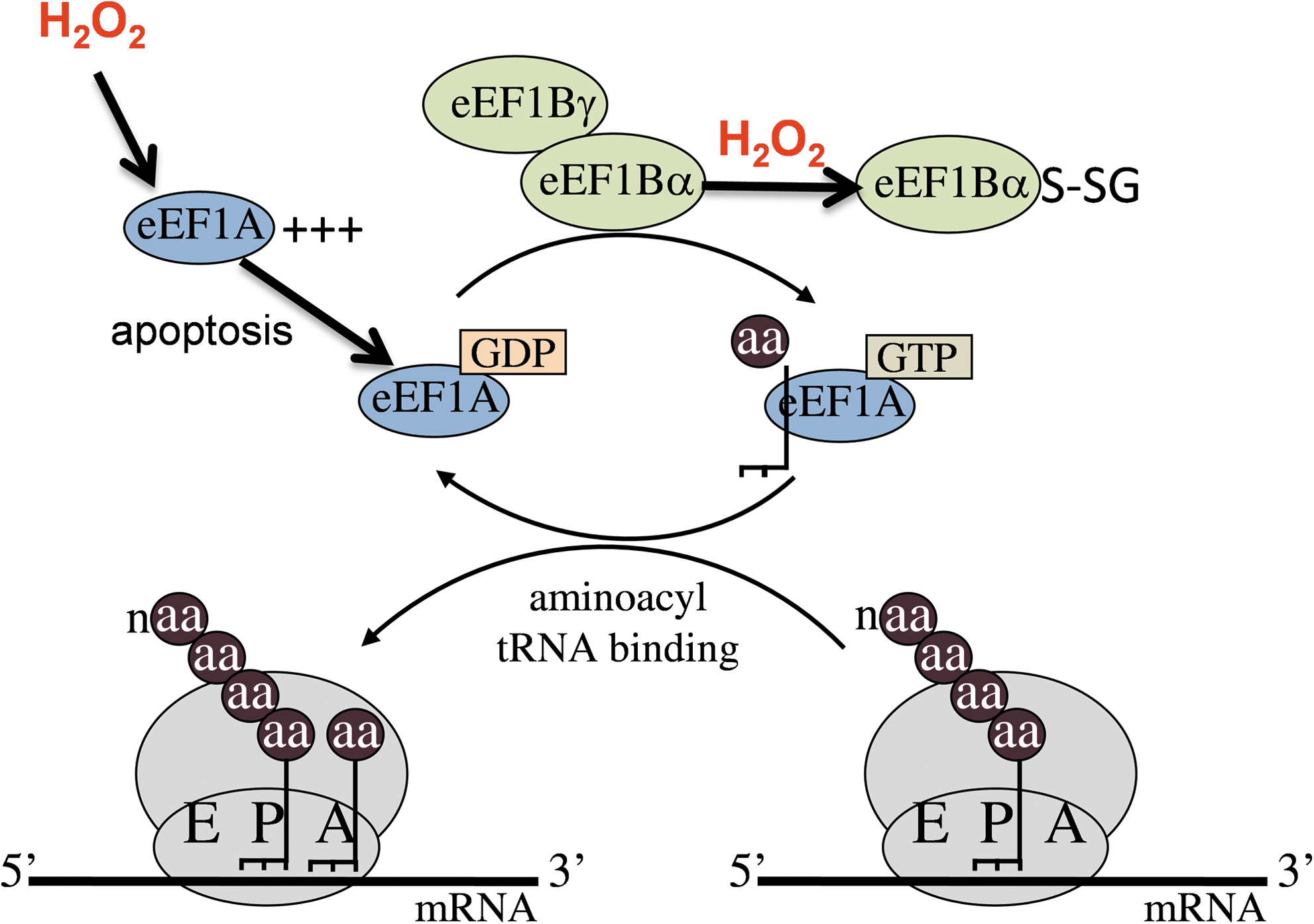
Ribosomes contain three tRNA-binding sites designated the aminoacyl (A) site, where the aminoacyl-tRNA binds; the peptidyl (P) site, where the peptide bond is formed; and the exit (E) site, where the deacylated tRNA exits the ribosome (89). The initiator tRNA binds directly to the P-site of the small ribosomal subunit and the A-site is poised to receive an aminoacyl-tRNA (46). During chain elongation, each additional amino acid is added to the nascent polypeptide chain, which requires correct positioning of an aminoacyl-tRNA in the A-site of the ribosome, formation of the peptide bond, and shifting the mRNA by one codon relative to the ribosome (Fig. 3). This is facilitated by two elongation factors designated eukaryotic elongation factor 1 (eEF1) and eEF2 (10). The insertion of an aminoacyl-tRNA into the empty A-site of an elongating ribosome is an important step in protein synthesis that must be performed with a high degree of fidelity to ensure that the correct amino acid is inserted. It is controlled by eEF1, which comprises a G protein and guanine nucleotide exchange factor (2, 10). eEF1A is the G-protein that in its GTP-bound (active) form binds an aminoacyl-tRNA and delivers it to the ribosome (Fig. 3). The correct codon–anticodon interaction stimulates GTP hydrolysis and eEF1A is released in its GDP-bound (inactive) form. This leaves a deacylated tRNA in the P-site and a newly formed peptidyl-tRNA in the A-site. eEF1B is the guanine nucleotide exchange factor that recycles eEF1A-GDP back to its active GTP-bound form (64). There is increasing evidence that eEF1 plays important roles in the control of gene expression, particularly during the cell cycle. eEF1A has been proposed to function in diverse cellular processes, including signal transduction, cytoskeletal organization, apoptosis, and oncogenic transformation (49). Similarly, eEF1B function has been implicated in control of the cell cycle and change in its expression levels and phosphorylation status has been implicated in oncogenesis and viral transformation (52). The ribosome then moves along the mRNA by one codon and eEF2 catalyzes the GTP-dependent translocation of the deacylated tRNA in the ribosomal P-site and the peptidyl-tRNA in the ribosomal A-site into the E- and P-sites, respectively. The next codon is then available for interaction with another aminoacyl-tRNA, and the process is repeated until a stop codon is encountered.
Evidence that H2O2 causes inhibition of translation elongation has come from yeast studies. Deletion of GCN2 in yeast abrogates phosphorylation of eIF2α and results in translational resistance to amino acid starvation (38). Similarly, no inhibition of translation initiation is observed in a gcn2 mutant in response to H2O2 stress. However, in contrast to amino acid starvation, protein synthesis is still inhibited by H2O2 in a gcn2 mutant and is not restored to wild-type levels (93). These data indicate that in addition to the regulation of translation initiation mediated by Gcn2, translation is also regulated at the postinitiation phase in response to H2O2. It agreement with this idea, the average ribosomal transit time on mRNAs has been shown to increase by ∼50% after exposure to H2O2, confirming that oxidative stress can influence the rate of elongating ribosomes (93). Regulating mRNA expression levels by modulating translation elongation is relatively poorly understood at present. It is known that cells can alter the bulk rate of protein synthesis in response to different growth conditions or hormones by changing the overall rates of elongation and/or termination (49, 82). However, there are very few characterized examples where the expression of individual mRNAs are regulated via alterations in ribosomal transit times (61). Attenuating elongating ribosomes in response to oxidative stress conditions, as opposed to ribosomal initiation, offers the advantage that ribosomes remain bound to mRNAs and can rapidly resume protein synthesis once the stress is removed or detoxified. This is important because translation elongation requires significant energy input in the form of GTP for elongation factor activity and ATP for the formation of aminoacyl-tRNAs. For an oxidative stress condition, inhibiting elongation would also prevent continued protein synthesis during potentially error-prone conditions.
Regulation of eEF1A
Several studies have indicated functional links between eEF1A levels and apoptosis in mammalian cells. For example, H2O2 conditions that induce apoptosis in mammalian cells also increase the expression of eEF1A (Fig. 3). This upregulation is thought to be regulated at the posttranscriptional level and may be required for increased protein synthesis of the apoptotic machinery (12). The mouse eEF1A-2 isoform has been shown to interact with a peroxiredoxin (Prdx-I), and this interaction may promote increased H2O2 tolerance through a mechanism that increases expression of the Akt pro-survival factor (11). Interestingly, disruption of eEF1A-1 in Chinese hamster ovary cells has been shown to increase resistance to H2O2 and to protect against lipotoxic cell death induced by palmitate exposure (9). eEF1A-1 is rapidly induced in response to palmitate along with markers of ER stress and ROS, and may mediate specific changes in protein translation that are required for apoptosis and cell death. Similar changes were demonstrated in a mouse model of lipotoxic cardiomyopathy, suggesting that this cellular response may be involved in the pathogenesis of cardiomyopathy in metabolic diseases (9).
Regulation of eEF1B
In all eukaryotes, eEF1B contains at least two subunits (α and γ, Fig. 3). eEF1Bγ is thought to enhance the GDP/GTP exchange activity of eEF1Bα, but its precise in vivo role remains unclear (10, 43). Biophysical studies indicate that the yeast EF1 complex is organized as an [EF1A-EF1Bα-EF1Bγ] trimer containing a single eEF1Bγ subunit (44). The N-terminal domain of eEF1Bγ has sequence motifs characteristic of the Theta class of glutathione transferases (GSTs) (48), enzymes that catalyse conjugation of electrophillic compounds to glutathione (GSH) (91). However, the GST domain of eEF1Bγ purified from diverse plant and microbial species does not show any GST activity with the model substrate 1-chloro-2,4-dintrobenzene (63). A link with oxidative stress has been demonstrated since yeast mutants lacking either of two nonidentical eEF1Bγ subunits (Tef3, Tef4) are resistant to oxidative stress conditions, including exposure to H2O2 (75). Further, yeast eEF1Bα (Tef5) and eEF1A (Tef1) have been identified as oxidative targets that are modified by protein S-thiolation in response to H2O2 stress (Fig. 3), although the physiological significance of this post-translational modification is unknown (92).
Regulation of eEF2
Oxidative stress in mammalian cells promotes phosphorylation and oxidative modification of eEF2 (4, 55, 78). Additionally, reactive aldehydes formed during peroxide stress have been shown to decrease the levels of eEF2 causing reduced rates of elongation, particularly during aging (3). Phosphorylation of eEF2 is thought to inhibit translation by reducing the affinity of eEF2 for GTP and by decreasing ribosome binding (22, 67). Interestingly, this regulatory mechanism appears to be conserved in yeast. eEF2 is phosphorylated by the mitogen-activated kinase, Rck2, in response to osmotic stress conditions, resulting in translation inhibition (102). Rck2 is itself a substrate for phosphorylation by the Hog1 mitogen-activated protein kinase (MAP kinase), indicating a link between MAP kinase signaling and the control of protein synthesis (102). Both Hog1 and Rck2 are phosphorylated in response to oxidative stress, indicating that this signaling pathway can respond to diverse stress conditions (8). Rck2 has also been implicated in the attenuation of elongating ribosomes during oxidative stress conditions (99). Exposure to oxidative (tert-butyl hydroperoxide) or osmotic stress (NaCl) was found to cause a pronounced dissociation of polysomes in an rck2 mutant. Microarray analysis indicated that a number of weakly transcribed mRNAs associate more avidly with polysomes during stress conditions, consistent with a role for Rck2 in mRNA-polysome association. Rck2 has been proposed to target the interchange of mRNAs between polysomes and stress granules, which are thought to act as storage factors that sort mRNAs for degradation or later translation after the relief of stress conditions (99). Storage or degradation of mRNAs in P bodies and stress granules may play an important role in regulating gene expression during oxidative stress conditions and at least one yeast study has shown that the number of P bodies is significantly increased after H2O2 stress (62).
Regulation of translation termination during oxidative stress conditions
If a stop codon (UAA, UAG, or UGA) is translocated into the A-site, it is recognized by the eukaryotic release factor 1 (eRF1) (88, 113). When it binds to a stop codon, eRF1 activates hydrolysis of the ester bond between the completed polypeptide chain and tRNA in the P-site (Fig. 4). eRF3 is a GTPase that associates with eRF1 and is essential for the termination reaction. Although the exact molecular details of the termination reaction are still unknown, it is clear that termination must proceed with a high degree of fidelity so as to avoid any errors in the decoding process. This is crucial since suppression (readthrough) of stop codons might potentially have profound effects on the proteome through the production of C-terminally extended proteins. Few examples of translational regulatory mechanisms that affect termination factor activity are currently known. This is perhaps surprising given that arresting translation at the termination stage would provide a pool of mRNAs that are poised to rapidly release a completed polypeptide once the termination block has been removed.
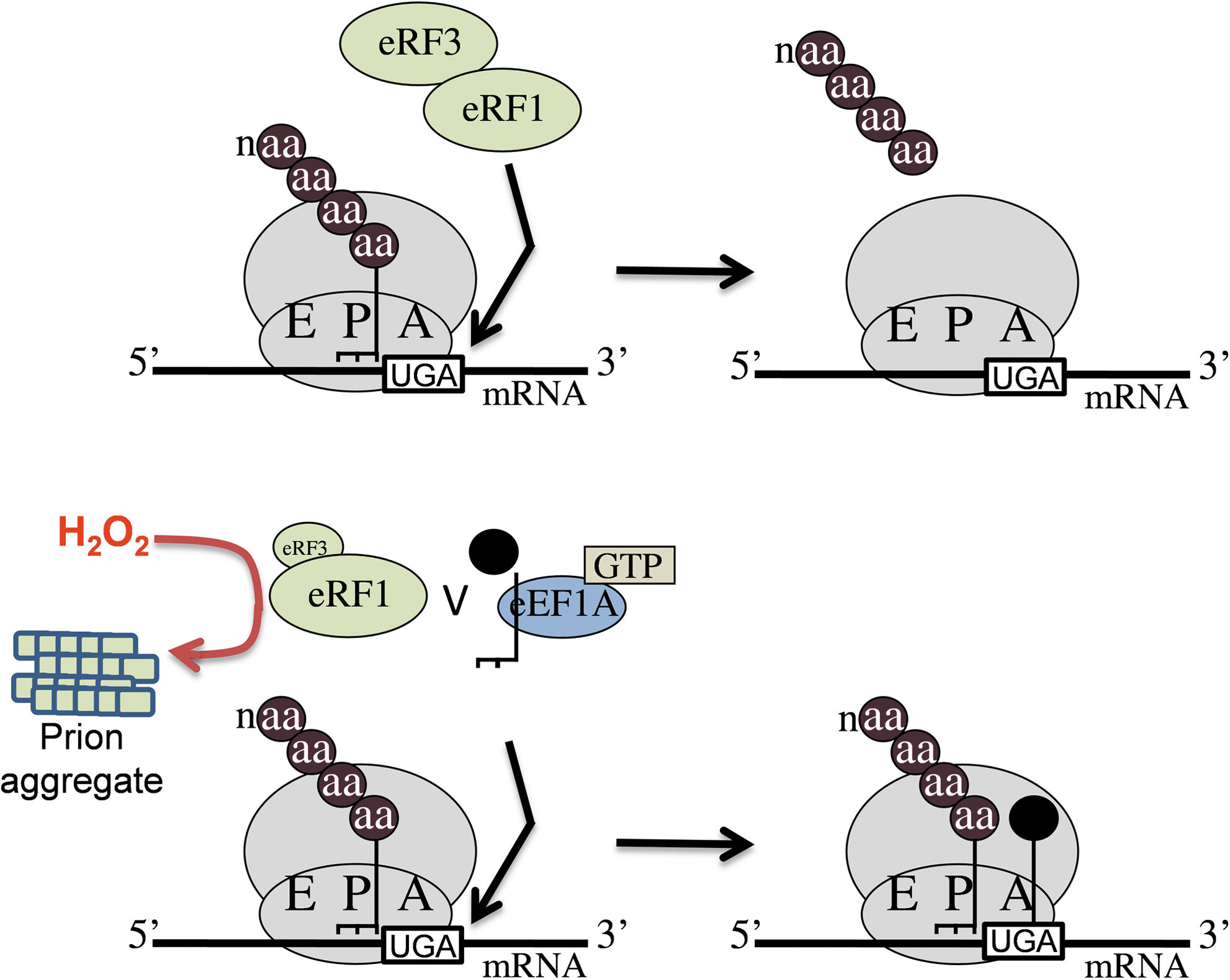
One good example of termination regulation is provided by the eRF3 (Sup35) from S. cerevisiae, which is uniquely able to form prion aggregates known as [PSI+ ] (114). Prions are novel protein-only infectious agents associated with a group of transmissible neurodegenerative diseases typified by human Creutzfeldt Jakob disease. Formation of Sup35 prion aggregates sequesters it away from its normal function in translation termination and elevated readthrough of termination codons is therefore a well-known phenotype of the yeast [PSI+ ] prion (Fig. 4). The molecular basis of how prions form spontaneously into infectious amyloid-like structures is poorly understood at present. The frequency with which the yeast [PSI +] prion form of Sup35 arises de novo is controlled by a number of genetic and environmental factors and, interestingly, has been shown to increase in response to stress conditions, including exposure to H2O2 (110). Additionally, the frequency of de novo formation of [PSI +] is greatly elevated in yeast mutants lacking the Tsa1 and Tsa2 peroxiredoxins (94). Peroxiredoxins play multiple roles in protection against stress, including acting as antioxidants and suppressing potential harmful oxidative damage to proteins after oxidative stress. Molecular oxygen is required for [PSI+ ] prion formation in a tsa1 tsa2 mutant, and oxidative stress (exposure to H2O2) elevates the frequency of de novo [PSI+ ] formation in this mutant (94). Taken together, these data suggest a role for oxidative damage in de novo prion formation. It has been suggested that the shift to the [PSI +] prion state may allow cells to reprogram gene expression such that new genetic traits become uncovered which aid survival during the altered conditions (110). In agreement with this idea, resistance to several environmental stress conditions correlates with the [PSI +] versus [psi −] status of cells (107, 108). Additionally, [PSI +]-induced effects on frame shifting increase the levels of antizyme, a negative regulator of polyamine biosynthesis. The resulting alterations in polyamine synthesis can account for many of the reported changes in sensitivity to environmental insults that have been correlated with the [PSI +] prion status of cells (68). Similarly, [PSI+ ] prion formation provides yeast cells with an adaptive advantage under oxidative stress conditions, since elimination of the [PSI +] prion from tsa1 tsa2 mutants renders the cells hypersensitive to H2O2 (94). The Tsa1/Tsa2 peroxiredoxins appear to function in protecting the translation machinery against H2O2 stress, but when they become overwhelmed, [PSI+ ] prion formation provides a mechanism for uncovering genetic traits that aid survival during oxidative stress conditions.
mRNA-Specific Translational Control Mechanisms
Adaptation to H2O2
It is now known that many cells can adapt to oxidative stress such that they become more resistant to a subsequent high dose after exposure to a low, nonlethal dose of a particular ROS (50, 96, 115). Adaptation depends on altering global expression patterns of genes encoding antioxidants and other stress-protective molecules (103). Adaptive responses have been extensively studied in the yeast S. cerevisiae, where it is now well established that there are a number of cellular responses that ensure the survival of the cell after exposure to oxidants such as H2O2, or products of oxidative damage (15, 42, 109). Although there has been considerable research on the cellular antioxidant defense systems that protect against ROS, less is known about how adaptation occurs. Its nature depends on the treatment. For example, heat shock induces resistance to H2O2, but H2O2 does not have the reverse effect. Cells adapted to H2O2 treatment become resistant to menadione (a superoxide generator), but not vice versa (24). This hierarchical response to stress may indicate the existence of a number of different adaptation systems that have overlapping components (103). Adaptation may be explained by the pattern of proteins that are produced in response to different oxidant treatments. Extensive transcriptional reprogramming is evident during adaptive treatments with H2O2 (26), but it is as yet unclear how these changes are translated into the cellular proteome. De novo protein synthesis is known to be required for the induction of resistance to H2O2 since it is abrogated by treatment with translation inhibitors such as cycloheximide (15).
Metabolic labeling and two-dimensional gel electrophoresis has shown that a substantial number of proteins are induced or repressed in response to low adaptive concentrations of H2O2 (28). Further insights into translational reprogramming are now possible using expression-profiling techniques, such as microarray analysis, that can be used to identify translational control patterns by comparing polysome and monosome-associated mRNA pools (5) (Fig. 5). For example, microarray/polysome analysis has been used to examine the nature and extent of mRNAs that are translationally regulated in response to two stresses that lower eIF2B activity (95). Amino acid starvation leads to an accumulation of nonaminoacylated-tRNAs and subsequent activation of Gcn2, whereas exposure to the fusel alcohol butanol inhibits eIF2B activity. Surprisingly, however, even though the stresses regulate the same translation initiation factor, eIF2B, they have quite different outcomes in terms of the specific mRNAs that are translationally controlled. This creates a highly specific stress response that facilitates adaptation to the particular stress condition (95). Although these stresses both act via eIF2B, there must be other stress-specific modulatory inputs on the translational pathway that ultimately change which mRNAs are translationally selected after stress. Similarly, H2O2 inhibits translation initiation via inhibition of eIF2B, yet the proteomic output is distinct to that of butanol and amino acid starvation (93). It is likely that regulation of translation elongation after H2O2 stress accounts for some of the differences with other eIF2B-specific response, but further investigation of individual mRNA regulatory systems will be required to understand these complex control pathways.
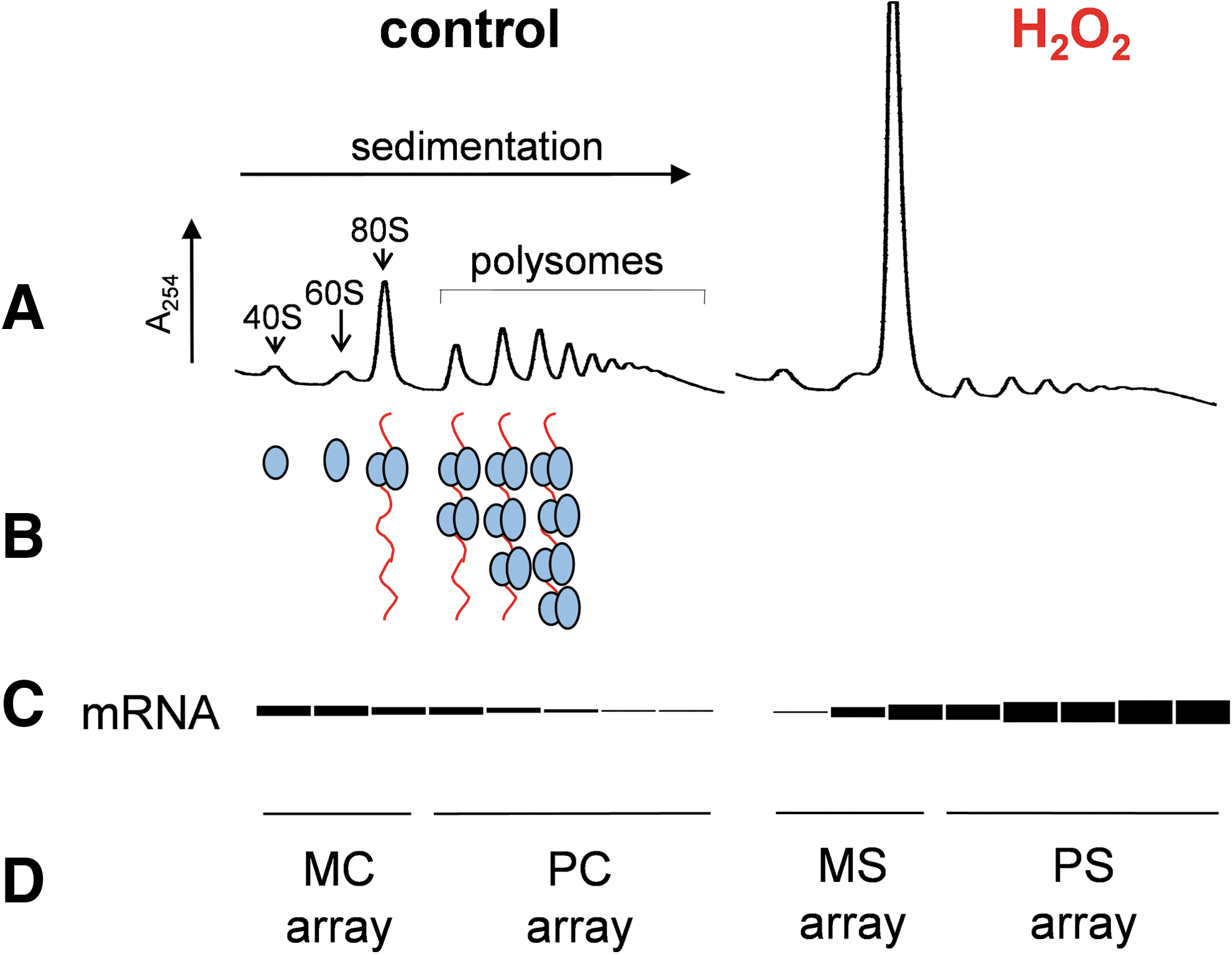
Polysome/microarray analysis revealed that low adaptive concentrations of H2O2 translationally induce mRNAs encoding yeast antioxidant defenses, including catalase, peroxiredoxin, thioredoxin reductase, and GST, which can directly detoxify H2O2 and its reaction products (93). Additionally, a number of metabolic genes were found to be translationally up- and downregulated consistent with a significant requirement for metabolic reconfiguration of energy requiring processes during oxidative stress conditions. Similar translation profiling studies have revealed extensive re-programming of translation during hypoxic conditions. Again, these data indicate that although the bulk of proteins synthesis is inhibited, significant translation of mRNAs encoding products that are thought to be important for adapting to hypoxia continues (97).
Examples of mRNA-specific translational control that are regulated by H2O2
Gcn4
There are relatively few characterized genes that are translationally regulated in response to H2O2 stress. The yeast Gcn4 is a particularly well-studied example of translational control, which is upregulated in response to phosphorylation of eIF2α (38). Translation of the GCN4 mRNA is activated in response to low TC levels in a mechanism involving four short uORFs. This translational control mechanism depends on the sequence context immediately surrounding the stop codons of the uORFs, which modulate the ability of ribosomes to reinitiate translation. During normal unstressed conditions, ribosomes translate uORF1 and can reinitiate at inhibitory uORFs2-4. Decreased TC levels mean that ribosomes take longer to bind the TC and are more likely to scan past uORF2-4 and reinitiate at GCN4 (38). However, uORFs are relatively rare in the yeast genome (∼13% of genes have them) and so cannot account for the large number of genes that are induced in response to H2O2 stress (51).
Despite the finding that phosphorylation of eIF2α appears to be a general response to oxidative stress, not all oxidants induce translational expression of GCN4 (60). GCN4 is induced by the well-known, Gcn2-dependent eIF2α phosphorylation mechanism in response to peroxide stress, but its translation must also be resistant to the postinitiation block in protein synthesis. This suggests that translation of the GCN4 mRNA is specifically able to continue during conditions where global translation is inhibited at both the initiation and postinitiation phases (93). Presumably, GCN4 expression is inhibited by other translational control mechanisms that act at the postinitiation phase in response to other oxidants.
Not only is Gcn4 upregulated in response to H2O2, but also it is specifically required for hydroperoxide resistance (60). Transcriptional profiling studies have shown that ∼10% of the yeast genome is regulated by Gcn4 in response to amino acid starvation, suggesting that Gcn4 acts as a master regulator of gene expression (70). However, Gcn4 is required for a more limited set of genes in response to H2O2 stress, and there is not a strong correlation with the amino acid starvation response at the genome-wide level, suggesting that other factors moderate the transcriptional output of Gcn4 during H2O2 stress conditions (60). Cluster analysis revealed a complex pattern of transcriptional regulation in gcn4 mutants. The requirement for translational regulation of GCN4 was most clearly shown by genes that require both GCN4 and GCN2 for their regulation. Upregulated genes include several metabolic genes affecting amino acid, lipid, carbon, and nitrogen metabolism, consistent with the idea that extensive metabolic reconfiguration is required after oxidative stress. Several genes encoding stress defenses or antioxidants are also upregulated, including a catalase gene (CTT1) that functions to detoxify H2O2. Downregulated genes include a number affecting energy-consuming processes, which is thought to be a common response during environmental stress responses (26). Loss of GCN4 was also found to alter the peroxide-regulated expression of many genes independent of Gcn2, indicating that Gcn4 activity may be regulated at the transcriptional or post-translational levels (60). In contrast to mutants lacking GCN4, gcn2 mutants are unaffected in H2O2 tolerance. This suggests that basal levels of expression of Gcn4-target genes are sufficient for peroxide tolerance. The requirement for Gcn2 is only apparent during adaptive conditions when it is required to inhibit translation initiation and to induce the expression of GCN4 (60).
ATF4
In eukaryotic cells, disulphide bonds are formed in nascent proteins within the lumen of the ER and are essential for the folding of proteins that are secreted or localized through the secretory pathway. Genetic evidence has identified a pathway for disulphide formation where oxidizing equivalents are transferred between thiol containing proteins and secretory proteins (30). Protein homeostasis in the ER represents a fine balance between the synthesis, folding/refolding, and degradation of damaged or misfolded proteins. An accumulation of misfolded proteins in the ER triggers the highly conserved UPR, which alters the expression of many genes involved in ER quality control. Oxygen is the major terminal electron acceptor and the UPR has been shown to contribute to cell death after protein misfolding, through the accumulation of ROS (34, 36). ER oxidoreductin 1 (Ero1) generates H2O2 as part of its reaction mechanism in disulphide bond formation (29). Crucially, the ER-transmembrane kinase PERK phosphorylates eIF2α in response to ER oxidative stress conditions (59). Downregulating protein synthesis by PERK-dependent phosphorylation of eIF2α serves to lower the unfolded ER-protein burden and generation of ROS, as well as to decrease the energy requirements associated with continued protein synthesis. Translation of the mammalian ATF4 transcription factor is upregulated under these conditions via an uORF-dependent mechanism analogous to yeast GCN4 (59, 111). Upregulation of ATF4 allows cells to adapt to ER stress as part of the integrated stress response. ATF4 activates genes encoding proteins involved in amino acid metabolism and import, particularly those required for glutathione biosynthesis. Other target genes include antioxidants ultimately protecting against the deleterious consequences of ER oxidation (34). GADD34 is a regulatory phosphatase subunit that selectively dephosphorylates eIF2α. Its activity is tightly regulated at the transcriptional level and is stress-induced as part of the integrated stress response. This means that it functions as part of a negative feedback loop that terminates signaling, facilitating the resumption of translation after reprogramming of the transcriptome as part of the integrated stress response (74).
Nrf2
Nrf2 is a basic leucine zipper transcription factor that is a key factor regulating the antioxidant response in mammalian cells through activation of the antioxidant response element in genes encoding many antioxidant and stress-defense molecules (73). Control of Nrf2 activity is a complex process, and is controlled in part by its tethering in the cytosol by another protein called Keap1. Keap1 appears to promote Nrf2 ubiquitination and degradation in a constitutive manner through the cullin-3-dependent pathway. Exposure to oxidative stress increases the abundance of stable Nrf2, which can translocate to the nucleus and activate antioxidant response element-dependent gene expression. Regulation of Keap1 activity is thought to be mediated through oxidation of its reactive cysteine residues in a mechanism, leading to stabilization of Nrf2 (25). The abundance of Nrf2 is also increased in response to H2O2 by a translation control mechanism (84). This mechanism depends on a cap-independent internal ribosomal entry site (54). This enables Nrf2 to be translationally upregulated during H2O2 stress conditions that inhibit global cap-dependent translation. Phosphorylation of eIF2α was found not to be sufficient to promote internal initiation on Nrf2 and it has been speculated that it may involve eIF4E phosphorylation, although the exact mechanism remains to be determined (54, 84). Interestingly, Nrf2 has been identified as a direct PERK substrate that is activated during the UPR (17). This may enable Nrf2 to play a complementary role with the ATF4 transcription factor during ER and oxidative stress conditions.
Oxidative Damage to the Translation Apparatus
There is increasing evidence that the translational apparatus is a target of oxidative damage, which might itself alter translation expression patterns. H2O2 exposure has been shown to inhibit protein synthesis in primary neurons, correlating with RNA oxidation and disruption of ribosomes (21). This may be a contributing factor during disease processes since oxidative damage to the protein synthetic machinery has been implicated in the pathogenesis of neurodegenerative disorders, including Alzheimer's and Parkinson's diseases (20, 39). Global analysis of protein carbonylation in yeast has identified a large number of ribosomal proteins that become oxidized in response to H2O2 stress, representing ∼80% of possible ribosomal protein targets (65). Many of these ribosomal proteins were found to form protein–RNA cross links that may contribute to ribosomal protein aggregation during peroxide stress (66). Further, exposure to high concentrations of H2O2 causes prominent upregulation of genes involved in ribosome biogenesis and rRNA processing, indicating that there may be a requirement to replace ribosomal proteins and rRNA, which become damaged during H2O2 stress (93). The yeast TSA1 peroxiredoxin has been shown to localize with ribosomes, where it appears to specifically function in protecting the translational apparatus against oxidative damage (85, 106). Oxidative disruption of the translation apparatus may therefore be more widespread than previously recognized, with ribosomal proteins representing important targets that must be protected against the formation of non-native protein structures and aggregates during stress conditions.
RNA appears to represent an important target that can become oxidized during oxidative stress conditions. For example, targeted mRNA oxidation has been implicated in Alzheimer's disease that might potentially produce aberrant proteins contributing to the pathogenesis of the disease (90). Translation studies using in vitro oxidized mRNAs have shown that translational errors are increased during translation of these mRNAs resulting in the production of short polypeptides (101). This decrease in translational fidelity suggests that oxidized mRNAs can be actively translated and the production of aberrant proteins may contribute to protein accumulation and aggregation diseases. Endonucleolytic cleavage of tRNAs appears to be a common controlled response to H2O2 stress in eukaryotic cells (105, 116). This may act to inhibit translation as an alternative mechanism to phosphorylation of eIF2α. H2O2 stress has also been shown to significantly elevate the rates of misacylation of methinione residues to nonmethionyl tRNAs in HeLa cells (72). These Met-misacylated tRNAs are used in translation and actively increase the incorporation of methionine into proteins. Given the ROS scavenging properties of methionine, it has been proposed that this increase in protein–methionine content acts to protect cells against ROS damage during viral infection and other oxidative stress conditions. Further evidence indicates that translational accuracy may play an important role in the response to oxidative stress since oxidative damage has been correlated with the fidelity of translation (47). Yeast strains containing error-prone rRNA mutations show fewer markers of oxidative damage compared with hyper-accurate strains, and it has been suggested that the energy that is required to maintain translational accuracy may reduce the energy available to combat ROS. Alterations in translational fidelity may therefore represent a common effect of oxidative damage to the translational apparatus.
Future Perspectives
H2O2 has frequently been used as a model oxidant, but care needs to be taken in extending conclusions made using H2O2 to other oxidants, since no one oxidant can be described as truly representative of oxidative stress despite the widespread use of single compounds such as H2O2 (103). While phosphorylation of eIF2α is clearly stimulated in response to diverse oxidants, future studies will be required to define the ROS specificity of a number of the translational control mechanisms discussed in this review article. Translational control mechanisms may prove to be more widespread than previously anticipated given the increasing evidence implicating ROS generation in cell growth, differentiation, and aging, as well as in the pathology of numerous disease and pathological conditions.
It is clear that H2O2 causes diverse effects on the protein synthetic machinery that can inhibit or stimulate the global rates of translation as well as regulate mRNA-specific translation. The changes in gene expression that are induced in response to H2O2 stress are complicated, requiring extensive translational and transcriptional reprogramming. The environmental stress cluster encompasses ∼900 yeast genes that are transcriptionally activated or repressed by a large number of stress conditions, including ROS such as H2O2 (26). Surprisingly, however, there is a poor correlation between transcriptional and translational regulation. For example, relatively few (∼15%) of the mRNAs that are translationally upregulated in response to H2O2 show concomitant increases in transcript levels (93). Some transcripts are increased in the absence of active translation and may provide a source of mRNAs that can become rapidly translated once the stress is removed. Conversely, some transcripts decrease in abundance, yet they show increased ribosome association in response to H2O2 stress. It is difficult at this stage to rationalize these complicated changes in transcription and translation, which presumably indicate a requirement to fine-tune gene expression during oxidative stress conditions (Fig. 6). It will be important in future studies to apply a more systematic approach that integrates studies on transcriptional and translational regulation with changes in the cellular proteome to fully understand the translational reprogramming, which is induced in response to oxidative stress conditions.
