
Abstract
Normal human pregnancy is considered a state of enhanced oxidative stress. In pregnancy, it plays important roles in embryo development, implantation, placental development and function, fetal development, and labor. However, pathologic pregnancies, including gestational diabetes mellitus (GDM), are associated with a heightened level of oxidative stress, owing to both overproduction of free radicals and/or a defect in the antioxidant defenses. This has important implications on the mother, placental function, and fetal well-being. Animal models of diabetes have confirmed the important role of oxidative stress in the etiology of congenital malformations; the relative immaturity of the antioxidant system facilitates the exposure of embryos and fetuses to the damaging effects of oxidative stress. Of note, there are only a few clinical studies evaluating the potential beneficial effects of antioxidants in GDM. Thus, whether or not increased antioxidant intake can reduce the complications of GDM in both mother and fetus needs to be explored. This review provides an overview and updated data on our current understanding of the complications associated with oxidative changes in GDM. Antioxid. Redox Signal. 15, 3061–3100.
I. Introduction to Gestational Diabetes Mellitus
Key points:
• Gestational diabetes mellitus (GDM) is commonly defined as any degree of glucose intolerance with first recognition during pregnancy.
• GDM is the most common type of diabetes found in pregnancy; its prevalence ranges between 1% and 14% of all pregnancies depending on the population studied and the diagnostic tests used.
• The prevalence of GDM is increasing worldwide, intensified with advancing maternal age, racial/ethnic disparities, and obesity.
• GDM is associated with markedly increased risk of adverse outcome for mother and infant, in both short- and long-term.
• The impact that GDM and its consequences has on community health, health economics, and human capital is substantial.
A. Incidence
The increasing prevalence of GDM parallels the temporal rise of type 2 diabetes in the nonpregnant population with low- and middle-income countries facing the greatest burden. There is a disparity by race and ethnicity with Asians having the highest rate, followed by Hispanics, African Americans, and Caucasians (49, 115). The International Diabetes Federation estimated that in 2010, India, China, the United States, Russia, and Brazil were the five countries with the largest numbers of people affected by diabetes (
B. Etiology and risk factors
GDM is a heterogeneous disorder involving a combination of factors responsible for decreased insulin sensitivity and inadequate insulin secretion. The underlying pathophysiology of GDM is in most instances similar to that of type 2 diabetes. The inability of the pancreatic beta cells to match increased insulin resistance to normalize systemic glucose translates into maternal hyperglycemia. Like type 2 diabetes, GDM is a multifactorial disease associated with both genetic and nongenetic/environmental risk factors (Table 1). Once the genetic predisposition of an individual is being challenged by increasing insulin resistance during pregnancy, there is an additional burden on the pancreatic β-cell, resulting in dysfunction, that is, inadequate insulin response to glucose challenges (33).
Genetic predisposition to GDM has been suggested since GDM tends to cluster in families. Several specific gene mutations identified in type 2 diabetes also increase the susceptibility to GDM although the underlying pathogenesis of the disease is still largely unknown (317). A growing number of common variants have been identified in candidate as well as noncandidate genes. The type 2 diabetes risk variants are located both in biological candidate and noncandidate genes. The pathophysiologic role of some of the risk alleles involves the impairment of β-cell function (KCNJ11, WFS1, CDKAL1, SLC30A8, HHEX/IDE, CDKN2A/B, and IGF2BP2), insulin resistance (PPARG), and obesity (FTO), which are major pathophysiological traits associated with type 2 diabetes loci. The strongest known type 2 diabetes association is found with variants in TCF7L2 (394). The number of molecular defects common to type 2 diabetes and GDM is now fueling the hypothesis that both are manifestations of the same disease. However, different environmental challenges may shadow the global figure as they occur with specific timing in the life cycle (198, 215). Although the expression of a gene variant significantly increases the risk of developing GDM, only a small proportion (<10%) of GDM can be accounted by genetic predisposition clearly pointing to other etiologic factors. Obesity, defined on the pre-gravid body mass index >30, high-fat diet, and advancing age represent the most important nongenetic factors that modulate the incidence of GDM (58).
C. Short- and long-term risks of GDM
1. GDM increases the risk of metabolic complications in the mothers
The link between GDM and postpartum diabetes in the mother has long been recognized. Approximately 5%–10% of cases of GDM are assumed to be previously undetected cases of diabetes, based upon background prevalence of diabetes in the population (131). The remaining majority of GDM cases is attributable to the metabolic challenge of pregnancy and impaired insulin secretory response (34). The reduced beta-cell reserve in GDM women can manifest as overt diabetes as late as one decade after pregnancy with a sevenfold higher risk than in women without history of GDM (19). It is associated with a higher risk of future maternal cardiovascular disease (42). The role of maternal micronutrients particularly low vitamin B12 and high folate associated with GDM may also increase the risk of gestational diabesity and later diabetes (191).
2. GDM increases the risk of metabolic programming for the offspring
GDM has long been managed with the primary goal of minimizing if not preventing the adverse outcomes for both the mother and her offspring. At the onset of the 21st century, serious perinatal complications associated with GDM or perinatal deaths have become uncommon. Macrosomia is the main factor linked to reported cases of peripartum complications in infants of women with GDM (248). However, establishing optimal regulation of maternal glucose levels continues to be a challenge in controlling perinatal morbidity. Failure in achieving early glycemic control and associated derangement in maternal metabolism contributes to impaired embryogenesis (322). Maternal hyperglycemia is considered as the primary teratogenic factor (6) although hyperketonemia, hypoglycemia, and excess free oxygen radicals have also been suggested to induce congenital anomalies (89).
3. GDM modifies fetal growth pattern
The macrosomia of fetuses of women with GDM has been defined using various criteria, including birth weight greater than the 90th centile, birth weight greater than 4000 g, and/or estimates of neonatal adiposity based on body composition measures. However, more than just affecting lean mass, GDM modifies body composition resulting in higher adiposity (45). What causes excess fetal growth in diabetic pregnancy? Since glucose is an easily measurable marker, most studies evaluating the effect of diabetes on fetal growth have used measures of glucose as a reference for satisfactory management. Results from the HAPO cohort have confirmed the relationship between glucose, birth weight, and adiposity while highlighting the role of insulin. Based on the early work of Pedersen, fetal insulin has long been considered as a primary anabolic factor for in utero fetal growth (311). Multiple studies have since confirmed the association between increased cord insulin concentrations and fetal macrosomia in both early and late gestation (246). When obesity is associated with GDM, additional mechanisms contributing to facilitate fetal adiposity relate to other maternal circulating nutrients for, for example, free fatty acids, and amino acids available in greater amount because of the plethoric environment of the mother (155).
In addition to the perinatal association with excessive fetal growth, there are significant long-term risks for the infants of women with GDM. There is an increase in the risk of obesity in children and adolescents in offspring of GDM women (46). The increased risk of obesity further extends to later life characterized by a higher incidence of type 2 diabetes, illustrating the impact of metabolic intrauterine programming (166). Clinical recognition of GDM is important because efficient therapy can reduce long-term sequelae in the offspring but also complications for the pregnant women. Besides insulin therapy, the interventions proposed for women with GDM are relatively noninvasive (diet, physical activity, lifestyle, and glucose monitoring alone) and have effects on maternal weight gain and energy metabolism limiting complications peri- and postpartum (137). Indeed, diet is critical for prevention and control of GDM and also in the prevention of induction of type 2 diabetes after GDM (98, 124). Animal studies have shown that maternal malnutrition predisposes offspring to develop insulin resistance and diabetes in the later life (98, 124). Although increased oxidative stress may be a mechanism of relevance in intrauterine programming of metabolic diseases further research in needed to clarify this point (225).
II. Brief Overview of Oxidative Stress
Key points:
• Oxidative stress is defined as a disturbance in the equilibrium status of pro-oxidants and antioxidants system in intact cells.
• Excessive oxidative stress can lead to massive cellular damage by acting on proteins, lipids, and DNA.
• Reactive oxygen species (ROS) is a term used to describe both free radical and nonradical derivatives of oxygen. ROS include superoxide anion (•O2 −), hydrogen peroxide (H2O2) and hydroxyl radical (•OH), and organic hydroperoxide (ROOH), peroxy radicals (RO•, alkoxy and ROO•), and hypochlorous acid (HOCl).
In the following section, a brief overview of ROS and antioxidants will be presented. More comprehensive reviews on free radicals and antioxidants can be found elsewhere (128, 354).
A. Reactive oxygen species
Lipid peroxidation results in the formation of a number of secondary products such as conjugated dienes and lipid hydroperoxides (LOOHs), and degradation products such as alkanes, aldehydes, and isoprostanes. Lipid oxidation intermediates are toxic for cells, mainly as a result of damage of the cell membranes. It has been etiologically involved in a variety of physiological, pathological, and clinical conditions. Lipid peroxidation leads to the production of the end products, including malondialdehyde (MDA) and 4-hydroxynonenal or trans-4-hydroxy-2-nonenal (4-HNE). MDA is a stable, toxic, and reactive aldehyde that is commonly used as a biomarker to measure the level of oxidative stress. It forms covalent protein adducts that are referred to as advanced lipoxidation end products, in analogy to advanced glycation endproducts (AGE). MDA forms a 1:2 adduct with thiobarbituric acid and produces thiobarbituric acid reactive substances (TBARS). The measurement of TBARS is a well-established method for screening and monitoring lipid peroxidation; however, this test is not specific for lipid peroxidation because MDA can also be formed by cyclooxygenase. LOOH, a marker of oxidative stress, is formed from unsaturated phospholipids, glycolipids, and cholesterol by peroxidative reactions under oxidative stress. 8-isoprostane (8-epi prostaglandin F2α or 15-F2t-IsoP) is an F2 isoprostane: one of a unique series of prostaglandin-like products derived from free radical-catalyzed, nonenzymatic oxidation of arachidonic acid independent of the cyclooxygenase enzyme. 8-isoprostane is considered to be an accurate, stable, and sensitive indicator of oxidative stress and endogenous lipid peroxidation.
ROS are able to oxidize proteins or convert lipid and carbohydrate derivatives to compounds that react with functional groups on proteins. Proteins can undergo different types of oxidation, including carbonylation, nitration of tyrosine, and oxidation of methionine to methionine sulphoxide. Carbonylation is an irreversible process that gives rise to protein carbonyl derivatives, which serve as more general and universal biomarker of oxidative stress.
B. Antioxidants
Antioxidants can be either soluble in water (hydrophilic) or in lipids (hydrophobic). Water-soluble antioxidants, which react with oxidants in the cell cytosol and the blood plasma, include vitamin C, glutathione, lipoic acid, and uric acid. Lipid-soluble antioxidants, which protect cell membranes from lipid peroxidation, include carotenes, vitamin E, and ubiquinol (coenzyme Q). Antioxidants may be derived from exogenous sources such as the diet or they may be endogenous antioxidants that are produced within the cell. The distribution and level of antioxidants in tissues and fluids varies greatly (354).
Enzymatic antioxidants are capable of detoxifying superoxide; superoxide is first converted to H2O2 and then further reduced to give water. Superoxide dismutase (SOD) enzymes catalyze the first step and then catalases and various peroxidases remove the H2O2. In mammals, there are 3 forms of SOD: copper/zinc SOD (CuZnSOD), manganese SOD (MnSOD), and extracellular SOD (ECSOD). MnSOD and ECSOD are localized in the mitochondrial matrix and on the outer surface of cell membranes, respectively, whereas CuZnSOD is found in the cytosol. The glutathione system, which includes glutathione, glutathione reductase (GSR), glutathione peroxidase (GPx), and glutathione S-transferases (GST), also controls superoxide formation. GPx, an inducible enzyme, contains four selenium-cofactors that catalyze the breakdown of H2O2 and organic hydroperoxides. Human have at least four different GPx isozymes; GPx1 is the most abundant and is most active against H2O2, whereas GPx4 is a very efficient scavenger of LOOHs.
III. Brief Overview of Nitrative Stress
Key points:
• Nitric oxide (NO) is a signaling molecule with a wide range of biological effects such as vasodilatation, formed by NO synthases (NOS).
• Diabetes leads to changes in NO production and bioavailability.
• Oxidation of NO leads to NO-derived molecules such as peroxynitrite, a powerful oxidant.
• Peroxynitrites induce damage to proteins, lipids, and DNA. Peroxynitrite-induced damage can lead to cellular dysfunction and apoptotic and necrotic signals.
NO is generated from the metabolism of L-arginine by the enzyme NOS, of which there are three isoforms: neuronal (nNOS), inducible (iNOS), and endothelial (eNOS). All the isoforms use L-arginine and molecular oxygen as substrates and require the cofactors nicotinamide adenine dinucleotide phosphate (NADPH), 6(R)-5,6,7,8 tetrahydrobiopterin (BH4), flavin adenine dinucleotide (FADH+), and flavin mononucleotide (FMNH+), as schematized in Figure 1. Uncoupling of the three NOS isoforms, which leads to the formation of O2 − instead of NO, can occur (270, 347, 380). Both low arginine and low BH4 are related to NOS uncoupling (193).
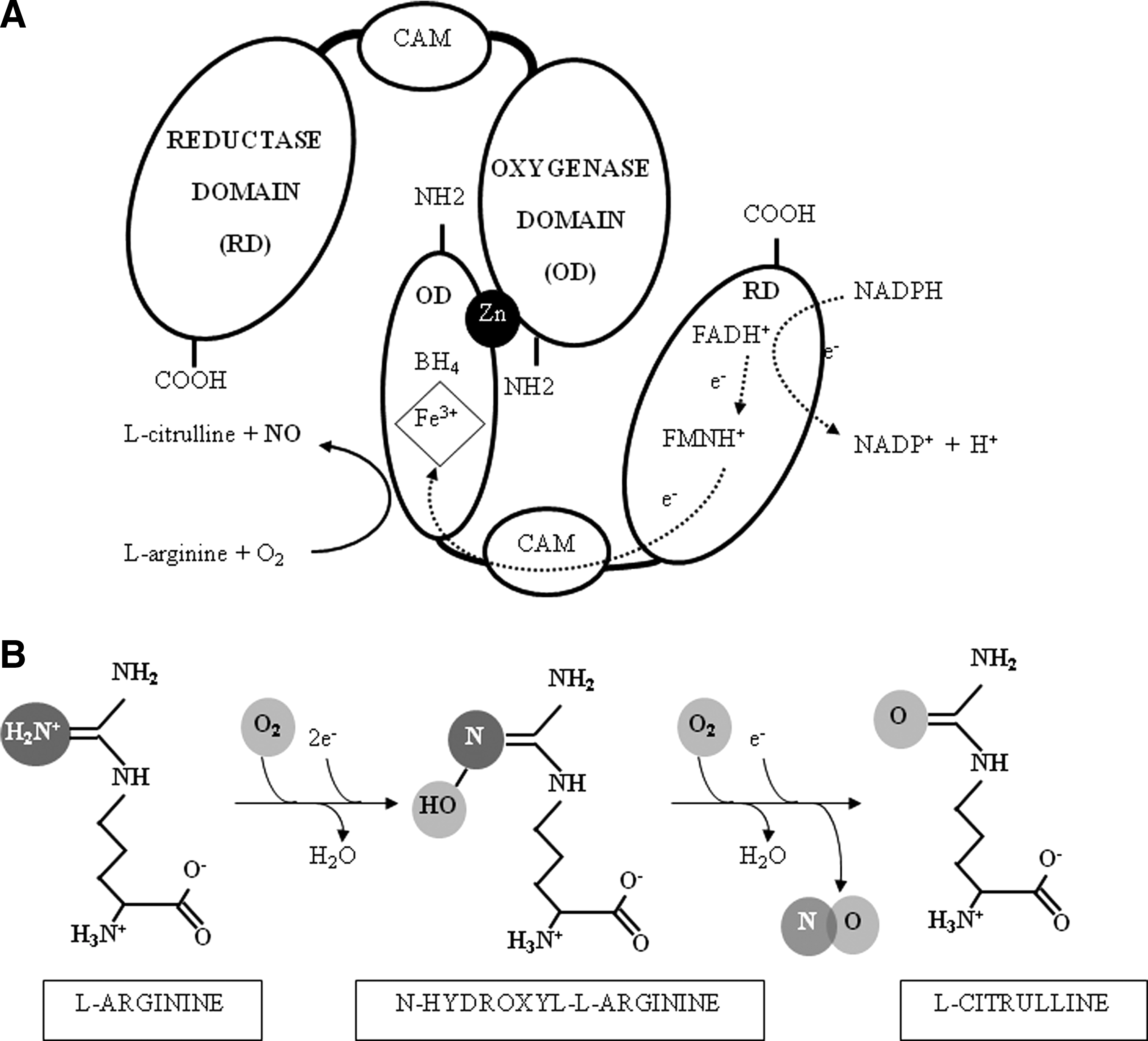
NO is a free radical, a simple diatomic gas, and a signaling molecule with a wide range of biological effects (Fig. 2). It classically exerts its action by binding iron-containing enzymes as soluble guanylate cyclase. By this classical pathway, NO transmits highly relevant biological effects such as muscle relaxation, blood pressure regulation, platelet aggregation, and neurotransmition (104, 146, 252, 255). In several pathological situations, including GDM, the NO-induced cGMPsignaling pathway is impaired due to further oxidation of NO, reduced NO bioavailability, and increased NO-derived S-nitrosylation and nitration pathways. Peroxynitrite is a powerful oxidant and nitrating molecule formed from O2 − and NO. Peroxynitrite can diffuse and cross cell membranes, but its half-life is very short and reacts very rapidly to nitrate or nitrosylate DNA, proteins, and lipids. DNA nitration leads to apoptosis of damaged cells, whereas protein nitration influences enzyme activities, and both protein and lipid nitration exerts influences on many signaling pathways (276).

The peroxynitrite-induced changes in DNA are the result of the capacity of peroxynitrite to induce DNA single-strand breaks, a damage that triggers the activation of the nuclear enzyme poly(ADP-ribose) polymerase (PARP). PARP uses NAD+ as a substrate to form poly(ADP-ribose), which plays a role in numerous physiological mechanisms, including DNA repair, regulation of genomic stability, and gene expression. However, pathological alterations are induced when PARP is overactivated and lead to cellular dysfunction, induction of apoptotic signals, and necrotic cell death (277).
The high amounts of NO produced by iNOS can have beneficial microbicidal, antiviral, antiparasital, and antitumoral activity (25, 229). On the other hand, aberrant iNOS induction is involved in the pathophysiology of human diseases such as asthma, arthritis, multiple sclerosis, colitis, psoriasis, neurodegenerative diseases, tumor development, transplant rejection, and septic shock (25, 150, 192). Hyperglycemia induces iNOS gene expression and is involved in the consequent generation of nitrosative/nitrative stress (190, 384).
IV. Oxidative Stress in the GDM Mother
Key points:
• A hyperglycemic environment is associated with oxidative stress. Likewise, in women with GDM, there is an overproduction of free radicals, and the radical scavenger function mechanisms are impaired.
• Oxidative stress in the GDM mother is presented below, and summarized in Table 2.
GDM, gestational diabetes mellitus; GPx, glutathione peroxidase; GST, glutathione S-transferases; GSH, reduced glutathione; MDA, malondialdehyde; LOOH, lipid hydroperoxide; TAC, total antioxidant capacity; TBARS, thiobarbituric acid reactive substances.
A. Oxidant species
1. Lipid peroxidation
Measures of lipid peroxidation include MDA, 4-hydroxynonenal, TBARS, and LOOH. Maternal MDA levels in serum and plasma are increased in GDM women compared to normal glucose tolerant (NGT) pregnant women (56, 174, 230, 282, 336). Likewise, enhanced levels of TBARS (239, 296) and LOOH (38, 239) have been reported in diabetic women. Further, there is a significant positive correlation relationship between maternal HbA1c and MDA (20, 167, 282), suggesting that higher levels of lipid peroxidation are evident in patients with poor glycemic control. No differences in TBARS and LOOH between GDM and control have also been reported (82, 273, 348).
2. Protein oxidation
Human serum albumin (HSA), the most abundant protein in amniotic fluid, can undergo modifications in response to oxidative stress (269). There is some evidence to suggest that amniotic fluid is oxic in the first trimester of pregnancy (227). In a very recent study, it was shown that there is increased protein oxidation in amniotic fluid of GDM women obtained before 15 weeks of gestation (27). Specifically, amniotic fluid HSA isoforms were compared between GDM and NGT women collected at 15 weeks of gestation and analyzed by mass spectrometry (27). The relative contribution of permanently oxidized HSA was greater and reversibly oxidized cysteinylated HSA was lower for GDM compared to NGT samples. These results show that amniotic fluid HSA is highly oxidized and that the increased oxidative stress associated with GDM alters amniotic fluid albumin toward the irreversibly oxidized isoforms, thus suggesting that the path toward GDM has been set in the first trimester of gestation. Currently, the accepted method of diagnosing GDM is via glucose blood testing between 24 and 28 weeks of gestation. Although a direct clinical benefit of the early diagnosis of GDM remains to be established conclusively, identification of women at greatest risk would allow triage of the patients to an appropriate model of care and identify a group who are at particular need of glucose tolerance assessment. Early diagnosis may also minimize exposure of the developing fetus to suboptimal conditions and prevent perinatal complications and their sequelae.
When proteins undergo oxidative damage, they become increasingly susceptible to proteolytic degradation. Erythrocytes contain proteolytic enzymes that can degrade oxidatively damaged proteins such as hemoglobin, thus preventing the accumulation of nonfunctional proteins and protein fragments. GDM is associated with higher levels of maternal erythrocyte proteolytic activity than NGT controls (167).
3. Transitional metals
The formation of the extremely reactive •OH from O2 − and H2O2 is catalyzed by iron; this is referred to as the Haber-Weiss reaction. There is now increasing evidence that increases in transitional metal, such as iron, may play a role in the generation of oxidative stress (287). Administration of an iron supplement during the third trimester of pregnancy is associated with significantly increased TBARS in the 27 supplemented women compared with controls (196). The positive association between body iron stores and the development of glucose intolerance and thus type 2 diabetes has long been recognized (241). However, data now also exist demonstrating a similar relationship between hemoglobin, serum ferritin, transferrin saturation, and/or iron concentrations and GDM (1, 24, 200 –202, 331).
Free copper or low-molecular-weight copper complexes catalyze the reaction between O2 − and H2O2 producing the •OH. In addition, copper binds to free thiols of cysteines, resulting in oxidation and subsequent crosslinks between proteins leading to impaired activity. GDM is associated with increased copper contents in serum when compared to NGT women (361).
B. Antioxidants
As presented above, in much of the literature, increased levels of ROS are consistently observed in the maternal circulation of women with GDM. However, there are discrepancies on the expression and activity of antioxidants in GDM, which may be due to differences in the criteria for diagnosis of GDM and gestation at which sample was collected. In the following section, all the literature pertaining to antioxidants in GDM will be reviewed. A complete summary of all the available data is presented in Table 2.
1. Nonenzymatic antioxidants
Maternal circulating levels of α-tocopherol are unchanged (82, 174, 282, 311, 327) or lower (121, 336) in GDM women. Lower (175, 336) and higher (82, 121) maternal plasma vitamin C levels have also been reported.
Glutathione is present in high concentrations and as such is considered of the most important cellular antioxidants. It can exist in either a reduced (GSH) or oxidized (GSSG) state. Both lower (296, 336) and higher (82) GSH levels in GDM women have been reported.
High levels of uric acid have casually been associated with insulin resistance for a while (134). However, it is only recently, in prospective follow-up studies (the Rotterdam Study), that high serum uric acid has been identified as a novel strong and independent risk factor for type 2 diabetes (76). Significantly elevated levels of serum uric acid are observed in GDM as compared to those in controls (175).
In addition to the above-mentioned antioxidants, several other natural antioxidants have been implicated in GDM. They include, but are not limited to, β-carotene, selenium, flavonoids, and vitamins B1 (thiamine), B9 (folic acid), and B6 (pyridoxine, pyridoxal, and pyridoxamine). When compared to NGT women, levels of serum selenium (177, 341) and folic acid (75) are significantly lower in pregnant woman with GDM.
2. Enzymatic antioxidants
Enzymatic antioxidants are capable of detoxifying superoxide; superoxide is first converted to H2O2 and then further reduced to give water. SOD enzymes catalyze the first step and then catalases and various peroxidases remove the H2O2. SOD activity significant increases (223, 336), decreases (56, 82, 121), or does not change (20, 282) in women with GDM when compared to NGT controls. On the other hand, catalase activity is GDM women (20, 336).
The glutathione system includes GSR, GPx, and GST. GPx activity is unchanged (273), higher (336), or lower (282) in GDM women. Likewise, there are higher maternal serum GST levels in patients with GDM when compared to NGT controls (82).
Paraoxonase and arylesterase are endogenous free-radical scavenges that act together to bind to high-density lipoprotein (HDL) and low-density lipoprotein (LDL), protecting them from oxidation by hydrolyzing activated phospholipids and lipid peroxide products. Basal and salt-stimulated paraoxonase and arylesterase activities are significantly lower in patients with GDM, and their activities in GDM patients inversely correlate with lipid LOOH levels (38). The authors suggested that the decreased serum paraoxonase and arylesterase activities might play a role in the potential early pathogenesis for atherosclerotic heart disease in GDM beyond their antioxidant properties. Oxidative stress plays a crucial role in the development of atherosclerosis through the oxidation of LDL that subsequently leads to the formation of foam cells (41), and HDL is a well-known anti-oxidant molecule that prevents atherosclerosis (14). Thus, the independent association between serum paraoxonase activity with serum HDL and LDL levels, but not with oxidative parameters, is in keeping with and GDM being associated with subclinical atherosclerosis (342).
Haptoglobin (Hp), an Hb-binding plasma protein, exists in two major allelic variants. Hp is an acute-phase protein that, in response to interleukin (IL)-6, is synthesized primarily in the liver and to some extent in fat tissue. Hp forms a complex with free Hb that can be rapidly cleared by the liver and macrophages. As such, Hp is thought to act as an antioxidant as free Hb catalyzes the generation ROS (in particular, •OH) by the Fenton reaction. Hp1 has higher Hb binding and antioxidant capacity compared with Hp2. Women missing Hp2 have an increased risk to develop impaired glucose tolerance during pregnancy (256).
Total antioxidant capacity (TAC) is considered a measure of the oxygen radical absorbance capacity or the capacity of the sample to inhibit an oxidant reaction; however, this is not always reflective of all major antioxidants. No difference in maternal plasma TAC between GDM and NGT has been reported (20, 348). However, when TAC was corrected for uric acid (uric acid is considered to be the major determinant of TAC), it significantly decreased in GDM compared to NGT in both the second and third trimesters (348). This is in keeping with other reports of lower TAC in GDM women (121, 296).
3. Transitional metals
Selenium, zinc, and copper are commonly referred to as antioxidant nutrients, but they do not themselves possess any antioxidant properties, instead they are required for the activity of some antioxidant enzymes. There is some evidence that the levels of zinc (23, 361) and selenium (23, 177) are lower in women with GDM. The activity CuZnSOD is sensitive to tissue copper as these enzymes require copper as a catalytic cofactor. Thus, the increase in CuZnSOD activity observed in GDM may be due to zinc and copper levels in GDM. Consistent with this hypothesis, compared with normal pregnant women, the copper contents in serum of pregnant women with GDM are increased (361). This is also consistent with the ability of copper to produce excessive amounts of ROS (as detailed above).
C. Concluding comments
The maternal circulating levels of free radicals and antioxidants are altered in GDM pregnancies. Given that the placenta provides the interface of the maternal and fetal circulations, it may play a crucial role in protecting the fetus from adverse effects of the maternal diabetic milieu, whereas disturbances in placental function may exacerbate this state. In the following section, the effect of diabetes in pregnancy, from both human and animal studies, on oxidative stress, nitrative stress, and antioxidants in the placenta and fetus is discussed.
V. Oxidative Stress in the GDM Placenta and Fetus
Key points:
• In GDM pregnancies, there is an overproduction of free radicals, placental oxidation reactions are accelerated, and the radical scavenger function mechanisms are impaired.
• The relative immaturity of the antioxidant system facilitates the exposure of embryos and fetuses to the damaging effects of oxidative stress.
• Animal models of diabetes and pregnancy have been very useful to address the presence of increased oxidative stress in the intrauterine compartment throughout diabetic gestations.
A. Oxidative stress in the placenta
The placenta is a rich source of oxidants and antioxidants. There is increased metabolic activity in the mitochondria of the placenta that generates ROS and superoxide generation from NADPH oxidase. The placenta is also capable of inducing protective enzymatic and nonenzymatic scavengers against these free radicals. The placenta provides the interface of the maternal and fetal circulations, and it may play a crucial role in protecting the fetus from adverse effects of the maternal diabetic milieu, whereas disturbances in placental function may exacerbate this state.
1. ROS in the GDM placenta
The GDM placenta is associated with increased expression of xanthine oxidase (XO) (20), MDA (20, 181), 4-HNE (261), and protein carbonyl (71). Likewise, the placental release of 8-isoprostane is greater from women with GDM compared to NGT pregnant women (71, 206). Quite interestingly, there was a significant positive correlation between plasma glucose (2 h after a glucose challenge at the time of diagnosis) and placental release of 8-isoprostane, suggesting that lipid peroxidation may be, at least in part, associated with glycemic control.
Several studies have found that diabetes leads to an aberrant ROS generation during intrauterine development in genetic and chemical-induced experimental models of type 1 and type 2 diabetes (162). In diabetic rat models, together with higher lipoperoxidation in maternal and fetal blood, lipoperoxidation is increased in the placenta at different developmental stages and is greater at term gestation (74, 114, 275, 374). Indeed, in the mild diabetic model obtained by streptozotocin administration to rat neonates, studies performed in the postplacentation period have shown that TBARS concentrations are increased in the decidua, the placenta, and the fetuses, but that lipoperoxidation is elevated in the decidua than in the placenta, and higher in the placenta than in the fetuses, thus suggesting a protective role of the placenta from oxidative stress (291).
Data from our laboratory have demonstrated that GDM placenta have a reduced capacity to respond to oxidative stress. Specifically, we have shown that when placental tissue was subjected to oxidative stress (hypoxanthine plus xanthine oxidase [HX/XO]), 8-isoprostane release increased by twofold in normal pregnant women, but was unchanged in GDM (70). This was associated with a decrease in catalase and GPx gene expression. In contrast, in GDM placentas, there was no effect of HX/XO on antioxidant gene expression (204). This response to HX/XO was specific to placenta; adipose tissue (both subcutaneous and omental) from women with and without GDM responded similarly to an oxidative challenge. That is, oxidative challenge stimulated 8-isoprostane release equally in normal and GDM omental and subcutaneous adipose tissue with no effect of HX/XO on antioxidant gene expression (204). Taken together, these data suggest that in both normal placenta, and normal and GDM adipose tissue, induction of oxidative stress displaces the prooxidant-antioxidant balance of this defense system, by increasing the prooxidants and depleting the antioxidant capacities. However, in the GDM placenta, we hypothesize that it may be preconditioned by transient intracellular oxidative stress, which attenuates its responsiveness to further oxidative insult. That is, it has been exposed to oxidative stress during the course of pregnancy and to counteract this, it has increased placental antioxidants. Thus, when it is further challenged with further oxidative challenges, it may be better able to respond as it has increased antioxidants. Certainly, there are data to suggest that the ability of cells to accommodate oxidative stress may be enhanced by preexposure or preconditioning to a mild oxidative challenge, thus inducing resistance to subsequent oxidative stress. In support, our data show that the GDM placenta does not alter its antioxidant capacity in response to HX/XO (204). Similarly, some tissues, in response to diabetes, overexpress the genes for the antioxidant enzymes, whereas other tissues are more susceptible to oxidative damage (237). It has also been proposed that in the early stages of diabetes there may be an initial elevation in antioxidant enzymes to counteract oxidative stress, whereas chronic diabetes continually depletes the sources of antioxidant enzymes.
2. Antioxidants in the GDM placenta
There is certainly much evidence to suggest that in GDM pregnancies, to maintain redox homeostasis, the placental production and the activities of antioxidant enzymes increase. Although it has been reported that catalase activity is decreased in placenta from women with GDM (20), we have found increased catalase and GSR mRNA expression in GDM placentas when compared to NGT placenta (204). There is no effect of GDM on GPx activity (20, 71) and GPx mRNA expression (204) in human placenta.
Discrepancies in the levels of SOD in GDM placenta have also been reported. The level of total SOD in placental tissues of GDM (both diet- and insulin-controlled) patients is lower (181) or did not significantly change (20, 204). However, our studies report that CuZnSOD activity is significantly higher in GDM placenta (71). This is particularly interesting as it is mainly compartmentalized in trophoblast cells within the placenta and may thus serve as an important antioxidant at the maternal-fetal interface (362). It is suggested that increased SOD activity may be a compensatory mechanism against increased XO activity and superoxide production. However, the relative ratio of CuZnSOD to 8-isoprostane or protein carbonyl was lower in GDM placentas, suggesting that the increase in SOD is not sufficient to compensate for the increased oxidative stress (71).
The lectin-like oxidized LDL receptor-1 (OLR1) is the principal scavenger receptor responsible for the uptake for oxidatively modified LDL in placental cells (278). Interestingly, the protein expression (but not mRNA expression) of OLR1 is increased in GDM placenta (92).
Apolipoprotein D (ApoD) is a lipocalin antioxidant that is a component of HDL. ApoD is higher in GDM placenta (261). As the trophoblastic cells and villous macrophages were positive for ApoD, the authors hypothesized that these cells may play a scavenger role protecting the diffusion of lipoperoxidation products from the mother to the embryo. This is in keeping with enhanced levels of arachidonic acid and docosahexaenoic acids observed in the placenta of the GDM women (21).
Similar to the studies in humans, in the placenta from experimental models of diabetes and pregnancy, antioxidant enzymes can be found either up-regulated, to compensate the oxidative dysbalance, or down-regulated, overwhelmed by the increased ROS. These changes are dependent on the developmental stage and are generated in response to the gradual increase in ROS, which is more pronounced at term gestation (291, 374). Besides, compensatory increases in the gene Txnip, which codifies the thioredoxin interacting protein (a protein involved in oxidative stress responses), have been reported in placentas from diabetic mice (389).
There is some evidence to show a beneficial effect of antioxidants in placenta. Specifically, studies in mitochondria of human placentas revealed a beneficial effect of ascorbate on lipid peroxidation, which is mediated by recycling of α-tocopherol. With the limitation of the nonspecific nature of TBARS, these data indicate that release of lipid peroxides from mitochondria can be prevented by ascorbate and α-tocopherol in human placental tissue (245).
3. Reactive nitrogen species in the GDM placenta
Oxidative stress plays a significant role in both NO overproduction and loss of NO bioavailability (116, 259, 346, 380). Oxidative stress leads to iNOS-dependent increases in NO production in different tissues. Although reductions in eNOS are related to diabetes-induced endothelial dysfunction, eNOS increases can also be induced by diabetes, and this is the change mostly found when eNOS is evaluated in gestational tissues in GDM. In addition, excess of NO production leads to changes in NOS function, yielding O2 − instead of NO. Moreover, increases in ROS lead to a reduction in NO bioactivity and increase the formation of peroxynitrites.
NO can also exert antioxidant effects (377). NO induces the expression of antioxidant enzymes MnSOD, CuZnSOD, and heme oxygenase-1 and increases intracellular glutathione concentration (250). Although NO stimulates O2 −-induced lipoperoxidation in membranes, it can also mediate protective reactions to inhibit O2 − and ONOO•− induced lipoperoxidation (305).
As a main regulator of vasodilatation, vascular remodeling, and angiogenesis, NO has been a focus of research in GDM. NO production has been found increased in the placenta, placental veins and arteries, and in umbilical vein endothelial cells from GDM patients (99, 288, 329, 359), although some studies have shown no significant increases in circulating nitrates/nitrites and no changes in placental NOS activity in placental tissues from GDM patients (83, 222). Accordingly, NOS expression is also altered, as iNOS has been found overexpressed in the placenta and eNOS increased in umbilical vein endothelial cells from GDM patients (308, 314). However, other studies have shown no changes in the expression of NOS isoenzymes in the placenta from GDM patients (83).
In GDM, increases in ROS and NO production, evident in the placenta and umbilical vessels, lead to peroxynitrite formation. Indeed, there is evidence of protein nitrosylation in placentas, umbilical arteries, and umbilical veins in insulin-treated GDM patients, although not in diet-treated GDM patients (144). This indicates that in those cases in which insulin is required to prevent aglycemia there is risk of peroxynitrite-induced damage. In platelets from GDM patients, elevated NOS activity and peroxynitrite production have been reported, possibly associated with platelet dysfunction and membrane damage due to increased lipid peroxidation (144, 239).
It is interesting that NO production, evaluated through the concentrations of nitrates/nitrites, is highly elevated in the placenta from mild diabetic rats at midpregnancy (40, 290). As strong protein nitration is found in term placentas from diabetic rats (40), increases in ROS, characteristic of term gestations and aggravated by the diabetic disease, may lead to the rapid conversion to peroxynitrites with the consequent loss of bioactive NO.
Collectively, these data provide evidence of reactive nitrogen species (RNS)-induced damage in GDM in the placenta and the vasculature of the mother, the placenta, and the umbilical cord, produced as a resulting consequence of exacerbated NO and ROS production.
B. Oxidative stress and the fetus
The above data demonstrate that the placenta is endowed with many antioxidants, some of which are increased in GDM. However, there is much data to indicate that maternal diabetes during pregnancy may induce oxidative stress in the newborn that may entail biochemical disturbances of the fetus (77, 259, 307). By using both chemical and genetic diabetes and pregnancy animal models, the studies carried out have confirmed the important role of ROS in the etiology of congenital malformations. Indeed, the relative immaturity of the antioxidant system facilitates the exposure of embryos and fetuses to the damaging effects of oxidative stress.
1. Oxidative stress and fetal malformations
Before the formation of the placenta, and despite the hypoxic state during embryo early organogenesis, there are increases in ROS, as determined by higher levels of isoprostanes and protein carbonyls in embryos from diabetic rats (51, 357). The increases in ROS, together with the impaired antioxidant activity and elevated RNS, as detailed below, are clearly related to the induction of malformations (88, 158). During early organogenesis, apoptosis is highly needed in an appropriate location and temporal pattern, and increased ROS are related to the increase in apoptosis induced in diabetic experimental models (55).
Congenital malformations are mostly induced in pre-gestational diabetic pregnancies, although they can also arise in GDM, possibly caused by a pre-existing diabetes first recognized during pregnancy (67). Congenital malformations, mainly cardiac and neural tube defects, are induced in both chemical-induced and genetic experimental models of diabetes (162). Interestingly, alterations in embryonic and feto-placental development in experimental models of diabetes have been associated with the increase in ROS in intrauterine tissues. Evidence of increased ROS has been found in embryos, fetuses, and placentas in streptozotocin- and alloxan-induced diabetes experimental models (89, 159, 274). Maternal diabetes-induced damage, clearly associated with the degree of ROS formed, is more marked in severe diabetic models (glycemia higher than 250 mg/dL) than in mild diabetic models (glycemia lower than 250 mg/dL) (158).
Interestingly, the susceptibility to the induction of malformations in chemical models of diabetes and pregnancy is clearly dependent on the rat strain and related to the embryonic concentrations of the antioxidant enzyme SOD, which are decreased in embryos from susceptible strains but not from malformation-resistant strains (53). Similarly, susceptibility to the induction of malformations in maternal diabetes is also clearly dependent on the embryonic levels/activity of antioxidant enzymes in genetic models of diabetes (364, 393). Of note, there is a decreased expression of GPx in malformed embryos from diabetic rats when compared to nonmalformed ones, an alteration localized in the developing heart (367).
There is evidence that several teratogens affect the developing embryo by increasing its oxidative stress. Human and animal studies show that the main mechanism of fetal damage induced by high levels of ionizing irradiation, cocaine and alcohol abuse, hypoxia, and cigarette smoking is also by increased embryonic oxidative stress.
Collectively, these data clearly indicate the relevance of ROS as teratogenic agents. Fetal malformations are mostly induced in the first trimester of pregnancy. As detailed in Section I, GDM is defined as glucose intolerance first detected at any time during gestation. Since GDM is mostly induced in the second trimester of pregnancy, its diagnosis is performed on weeks 24–28 of pregnancy, but, as detailed in Section IV, increased ROS have been detected in amniotic fluid in GDM before the 15th week of gestation. Therefore, the risk for fetal malformations is increased in pre-gestational diabetes and may affect GDM only in those cases in which the pathology is present in the first trimester of pregnancy, possibly as a result of the increased intrauterine ROS generated.
2. Nitrative stress and fetal malformations
As reviewed elsewhere, both NO production and NOS activity are enhanced in embryos from diabetic rats during organogenesis, alterations related to the induction of embryo malformations (88, 158). Moreover, peroxynitrite-induced damage has been found in the neural tube and developing heart in embryos obtained from diabetic rats during early organogenesis, being both neural tube defects and cardiac malformations the most common congenital defects induced by maternal diabetes both in patients and in experimental diabetic models (89, 160). It is interesting that there are many endogenous regulators of NO production in the embryo during organogenesis, suggesting that NO concentrations should be tightly regulated during organ formation. Indeed, leptin, Prostaglandin E2 (PGE2), 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2), and endothelin-1 can negatively regulate embryonic NO production (160, 161, 373). These bioactive molecules show changes in their concentrations and impaired capacity to regulate NO in diabetes and pregnancy models in which hyperglycemia is >250 mg/dL (160). The involvement of iNOS as the NOS isoenzyme responsible for NO overproduction in diabetic embryopathy has been addressed by the use of iNOS inhibitors and the evaluation of iNOS knockout mice, in which diabetes-induced congenital malformations have been found highly reduced (335). ROS-induced teratogenesis is also reduced in iNOS knockout mice (168).
The available data suggest that NO overproduction and NO-derived oxidant species, mainly peroxynitrites, are involved in the induction of congenital malformations. Therefore, RNS are likely to be related to the induction of fetal malformations in pre-gestational diabetes, and also in GDM if the metabolic impairments that lead to NO overproduction in intrauterine tissues are increased in the first trimester of pregnancy when most congenital malformations are induced.
3. Oxidative stress in the fetal organs
Studies performed in a genetic rat model of type 2 diabetes have shown that lipoperoxidation is enhanced in different fetal organs such as the heart and brain (275). Likewise, in the rat neonate, chemical-induced maternal diabetes induces increases in ROS and lipoperoxidation in different organs such as the liver, kidney, and brain (182, 298). In the offspring from diabetic rats, the plasma levels of lipoperoxides are elevated (385).
No changes, increases or reductions in the activity of SOD, catalase, and GPx, have been found in fetuses in different diabetes and pregnancy animal models, differences that are dependent on both the genetic background and the degree of metabolic impairment (162, 182, 275, 291). Studies performed in the offspring have shown that there are reduced concentrations of glutathione and SOD in different organs such as the liver, kidney, and brain in neonates born to chemical-induced diabetic rats (182, 298). Studies performed in chemical-induced pregestational diabetic rats have shown a decreased expression of CuZnSOD, MnSOD, and GPx in embryos from diabetic rats when compared to controls (324, 392). Similar decreases in SOD activity in the macrosomic offspring of diabetic rats have been demonstrated (385).
In animal models, under diabetic condition there was a significant decrease in the activity of endogenous antioxidant enzymes and of vitamins C and E in the embryos and their yolk sacs (274). Reduced concentrations of vitamin E have also been found in both embryos and livers from fetuses from diabetic animals (321).
The maternal surface of the trophoblastic microvilli of the human placenta is very rich in transferrin receptors (221). Iron from maternal transferrin is transferred from the syncytiotrophoblasts of the placenta into the fetus—a process that increases as pregnancy progresses (44). Although the oxidative stress and in vivo consequences caused by iron excess have been studied in detail, the effect of excess iron on the fetus is not known.
Relevant alterations in NO and peroxynitrite formation have been found throughout pregnancy in experimental models of diabetes (158). At the fetal stage, and even in mild diabetes and pregnancy animal models, NO production is increased (290).
A higher transfer of metabolic substrates from the maternal compartment, higher fetal insulin concentrations, and fetal macrosomia are characteristic in GDM patients. Interestingly, postmortem examinations in human fetuses with islet cell hyperplasia, an indicator of fetal hyperglycemia, have shown that only those fetuses that have increased insulin immunoreactivity in pancreatic islets show increased nitrotyrosine immunoreactivity in the central nervous system (141).
In summary, the available data indicate that maternal diabetes leads to oxidative and nitrative stress in many fetal organs, alterations associated with either impaired or compensatory responses in antioxidant enzymes. Although fetal organs can be evaluated mainly in experimental models of diabetes, the alterations in umbilical cord blood obtained at term gestations suggest that GDM exposes the fetus to both ROS and RNS, as detailed below.
4. Oxidative stress in the umbilical cord of GDM women
Certainly, evidence suggests that the fetus at term is exposed to oxidative stress, as higher ROS and lower antioxidants are evident in the umbilical cord blood of GDM women. The evidence is discussed below and summarized in Table 2.
Markers of reactive oxidative species are increased in diabetic pregnancies. MDA activity (20, 181, 230), GSH levels (181), and serum protein thiol levels (82) are increased in cord plasma from GDM women when compared to NGT women (20, 181, 230). The macrosomic offspring of women with GDM have enhanced TBARS levels (121). Others have reported no differences (273).
On the other hand, antioxidants are unchanged, higher, or lower in cord blood from diabetic women. Cord plasma vitamin E levels (20, 327), catalase activity (20, 336), GPx activity (20), SOD activity (20, 181), and TAC activity (20) are significantly decreased in GDM. Correlation analysis demonstrated a significant inverse relationship between maternal HbA1c (higher levels) and cord plasma TAC (lower levels), suggesting that oxidant stress and peroxidation reactions are in parallel with diabetic deterioration (20). These data suggest that maternal diabetes during pregnancy may induce oxidative stress in the newborn. On the other hand, SOD activity (20), erythrocyte GPx activity (82, 296), and apolipoprotein B (296) are elevated in cord blood from GDM women.
C. Concluding comments
In placenta and fetuses of GDM pregnancies, oxidative stress reactions are increased. In concert with this, there are alterations in the antioxidant defense mechanisms. The pathways that contribute to the increased oxidative stress observed in the GDM placenta are discussed below.
VI. Pathways Contributing to the Generation of Oxidative Stress in GDM
Key points:
• Hyperglycemia induces oxidative stress and cell and tissue damage through several metabolic mechanisms. These include the polyol pathway, formation of AGE, activation of protein kinase C (PKC), the hexosamine pathway, and increased oxidative stress generation by enhanced ROS production in the mitochondria.
• Pathological pathways involved in the generation of these metabolites are discussed below and illustrated in Figure 3.

A. Advanced glycation endproducts
An important source of free radicals in diabetes is the interaction of glucose with proteins leading to the formation of an Amadori product and then AGEs. AGEs, such as N ɛ-carboxymethyl-lysine (CML), are late-stage glycoxidation and glycation adducts of the Maillard reaction that form by nonenzymatic glycation through covalent attachment of highly reactive aldehyde or ketone groups of reducing sugars and the free amino groups on proteins, lipids, and nucleic acids. This can occur at intracellular and extracellular sites. In particular, elevated concentration of glucose metabolism products from glycolysis and the tricarboxylic acid (TCA) cycle initiate glycation of intracellular proteins. The interaction of aldehyde groups of glucose with free amino groups on proteins generates a Schiff's base. It spontaneously rearranges into a much more stable ketoamine, the Amadori product (16, 122). Amadori products are degraded into other reactive dicarbonyl compounds such as 3-deoxyglucosone and methylglyoxal, which can react directly with amino groups of intra- and extracellular proteins to generate AGE. This covalent modification of proteins has severe consequences and leads to altered protein function. Indeed, tissue and plasma of diabetic patients contain higher amounts of AGE (26, 117).
Extracellular AGE can bind to the AGE receptor (RAGE), a multi-ligand member of the immunoglobulin superfamily. Besides its capability to activate transcription factors such as nuclear factor-kappa B (NF-κB), it stimulates ROS formation by NADPH oxidase. In this way, AGE lead to cellular dysfunction and injury and to formation of ROS (303). There is also a soluble form of RAGE (sRAGE), which is a truncated form of the receptor produced by alternative splicing of RAGE mRNA (endogenous secretory RAGE, esRAGE) and proteolytic cleavage of membrane-bound RAGE by metalloproteinase action (proteolytically cleaved RAGE, c-RAGE). Circulating sRAGE is composed of only the extracellular ligand-binding domain lacking the cytosolic and transmembrane domains. It has the same ligand binding specificity of RAGE, and thus it competes with cell-bound RAGE for ligand binding, neutralizing AGE-mediated damage by acting as a decoy. Engagement of RAGE by AGE results in activation of intracellular signaling molecules resulting in oxidative stress and inflammation. Since oxidative stress generation and inflammation are closely associated with GDM (68, 206, 301), it is plausible that the AGE-RAGE system could play a role in the pathogenesis of this metabolic disease.
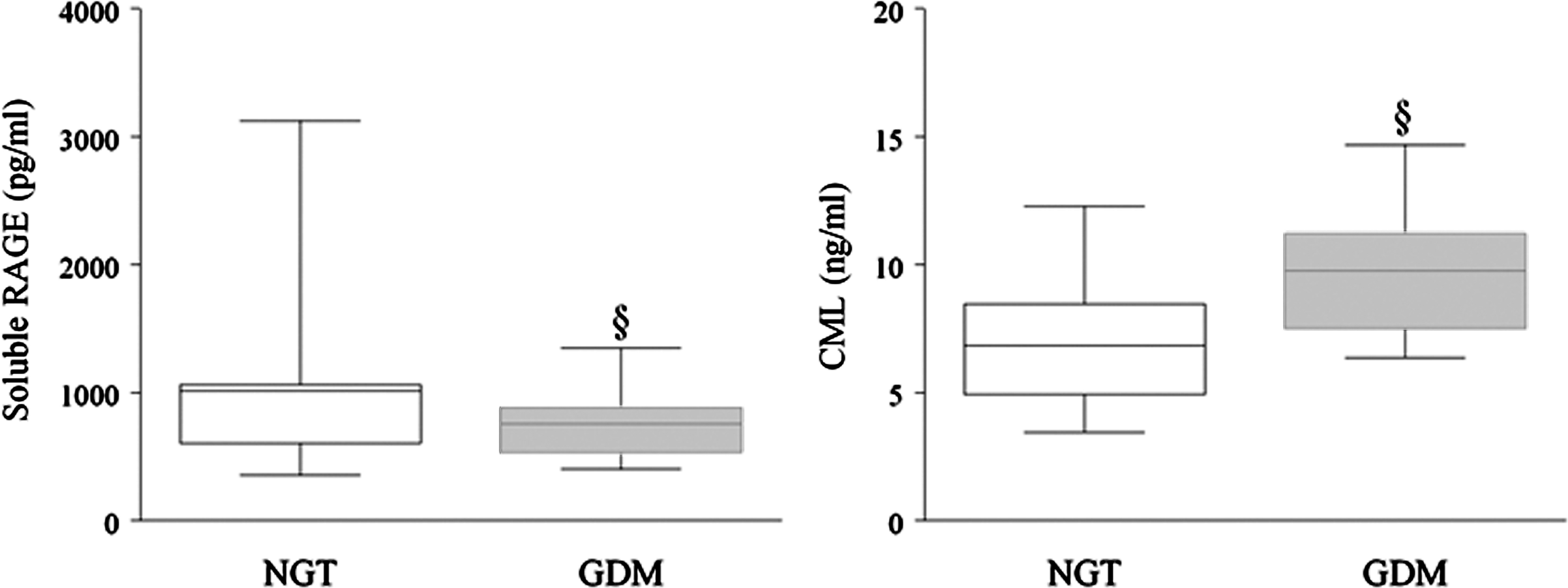
Although AGE and RAGE have recently been identified in human placenta (35, 57, 105) and increase in association with pregnancy and pregnancy complications (35, 57, 66, 96, 105, 279), to date, there are limited data available on the circulating levels of AGE in relation to GDM. In recent studies, we have profiled the maternal circulating levels of CML, sRAGE, and esRAGE during pregnancy from pregnant women with NGT and GDM. The maternal plasma concentrations of CML, sRAGE, and esRAGE were measured from 46 NGT women and 40 women with GDM at the time of term Caesarean delivery. Maternal circulating CML levels were significantly higher in GDM women than in NGT women. On the other hand, statistically lower sRAGE and esRAGE levels at the time of term delivery were also observed in GDM women compared to NGT women (Fig. 4).
During pregnancy, the AGE-RAGE axis may be involved in oxidative and inflammatory responses. By increasing intracellular oxidative stress, AGE activates NF-κB, thus promoting up-regulation of various NF-κB controlled target genes. For example, AGE-RAGE signaling plays a pivotal role in regulating the production and/or expression of pro-inflammatory mediators such as cytokines and oxidative stress, as well as endothelial dysfunction in type 2 diabetes via NF-κB (107). In previous studies, we used in vitro human tissue explant system to examine the potential inflammatory effects of AGE-BSA in human intrauterine tissues. Our data show that AGE-BSA, but not nonglycated BSA, has pro-inflammatory actions in human gestational tissues. Specifically, AGE-BSA stimulated the release of the pro-inflammatory cytokines IL-1β, IL-6, IL-8, and tumor necrosis factor (TNF)-α and prostaglandins PGE2 and PGF2α. These pro-inflammatory actions of AGE-BSA were elicited through a number of intracellular signaling pathways, namely, extracellular-signal-regulated kinase 1/2 and NF-κB (210). NF-κB and MAPK activate several pro-inflammatory genes, including pro-inflammatory cytokines, the adhesion molecules vascular cell adhesion molecule (VCAM)-1, and intercellular cell adhesion molecule (ICAM)-1, and RAGE causing cellular inflammation. This is consistent with GDM being closely associated with low-grade inflammation (68, 184) and atherosclerosis (7, 129). Additionally, the activation of cytokines by AGE in human placenta may also be involved in insulin resistance associated with GDM (63). We also reported increased release of 8-isoprostane, a marker of oxidative stress, from human placenta in the presence of AGE (210). Thus, elevated circulating AGE observed in this study may contribute to elevated oxidative stress concentrations observed in GDM (71, 113). In isolated human first-trimester trophoblasts, AGE stimulate secretion of chemokines such as macrophage inflammatory protein (MIP)-1α and MIP-1β, induces apoptosis, and suppresses the secretion of human chorionic gonadotropin, an effect that could be suppressed by inhibitors of NOS or the NF-κB pathway (187), thus suggesting that RNS as well as ROS contribute to AGE-mediated actions in the human placenta. There is also much evidence to show that AGE activates the expression of adhesion molecules such as VACM-1, ICAM-1, and E-selectin (15).
B. Hexosamine pathway
Hyperglycemia elevates oxidative stress and increases the activation of the hexosamine biosynthetic pathway (295). This pathway of glucose metabolism uses fructose-6-phosphate derived from glycolysis to metabolize glucosamine-6-phosphate by glucosamine-6-phosphate amidotransferase. Glucosamine-6-phosphate is a competitive inhibitor of glucose-6-phosphate dehydrogenase (G6PD), the rate-limiting enzyme of the pentose phosphate pathway. The pentose phosphate pathway is an alternative pathway for glycolysis and produces the major portion of NADPH in cells. The activation of G6PD converts glucose-6-phosphate into 6-phosphogluconate, and, subsequently under formation of NADPH, to cellular ribose-5-phosphate (30). The NADPH generated is used to maintain the redox state through the reduction of GSSG to its reduced form GSH. G6PD is the rate-limiting enzyme of the pentose phosphate pathway and its inhibition, for instance, by glucosamine-6 phosphate produced in the hexosamine pathway, leads to decreased NAPDH concentrations, diminished cellular GSH levels, and elevated oxidative stress (395). The activity of G6PD also rapidly increases in response to intracellular ROS production (148). Therefore, G6PD seems to constitute a critical cytosolic antioxidant enzyme essential for the maintenance of the cytosolic redox status. In an animal studies, it prevents embryopathies (2), demonstrating the protective role of G6PD against oxidative stress. G6PD is present in human placental trophoblasts (238); however, the effects of GDM on the G6PD levels in placenta are not known.
UDP-N-acetylglucosamine is generated as a product of the hexosamine pathway, which is a substrate for glycosylation of several transcription factors, leading to activation of gene expression (303). In addition, nuclear and cytoplasmic proteins are modified by N-acetylglucosamine via phosphorylation. Thus, activation of the hexosamine pathway by hyperglycemia may cause changes in gene expression as well as in protein function (31). Again, there is a paucity of data with respect to placenta and GDM.
C. Polyol pathway
The polyol pathway leads to the generation of ROS via a number of mechanisms. Under normal conditions, the enzyme aldose reductase has a low affinity for glucose; however, hyperglycemia promotes the conversion of glucose to polyalcohol sorbitol. Since sorbitol does not cross cell membranes, it subsequently accumulates within the cells and causes cell and tissue damage (93). Sorbitol can be further oxidized by sorbitol dehydrogenase to fructose with concomitant reduction of NAD+ to NADH. Enhanced cytosolic NADH to NAD+ ratio inhibits the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) activity and, therefore, provides increased substrate availability to complex I of the mitochondrial respiratory chain. Inhibition of GAPDH also elevates the concentration of AGEs (31, 100, 303). The polyol pathway also results in decreased levels of NADPH, glutathione, and antioxidant equivalents, thus leading to an overproduction of intracellular ROS. The effect of GDM on placental polyol levels in GDM pregnancies is not known.
D. NADPH oxidase
NADPH oxidase is a cytosolic enzyme complex accounting for ROS generation by electron transport and especially important in redox signaling. It first was discovered in neutrophils, where it plays a crucial role in nonspecific host-pathogen defense. It is composed of membrane-bound subunits gp91 phox (Nox2)/Nox1/Nox4, p22 phox, the catalytic site of the oxidase, and the cytoplasmic regulatory components p47 phox and p67 phox. Under diabetic conditions it can be stimulated by AGE, insulin, and angiotensin II. Hypoxia possibly induces all these stimuli, which can activate NADPH oxidase (Fig. 5). Once activated in response to high glucose NADPH oxidase catalyzes the transfer of electrons from NADPH to molecular oxygen to produce O2 − and H2O2 (106, 125). High glucose levels lead to generation of ROS by stimulation of NADPH oxidase (379).
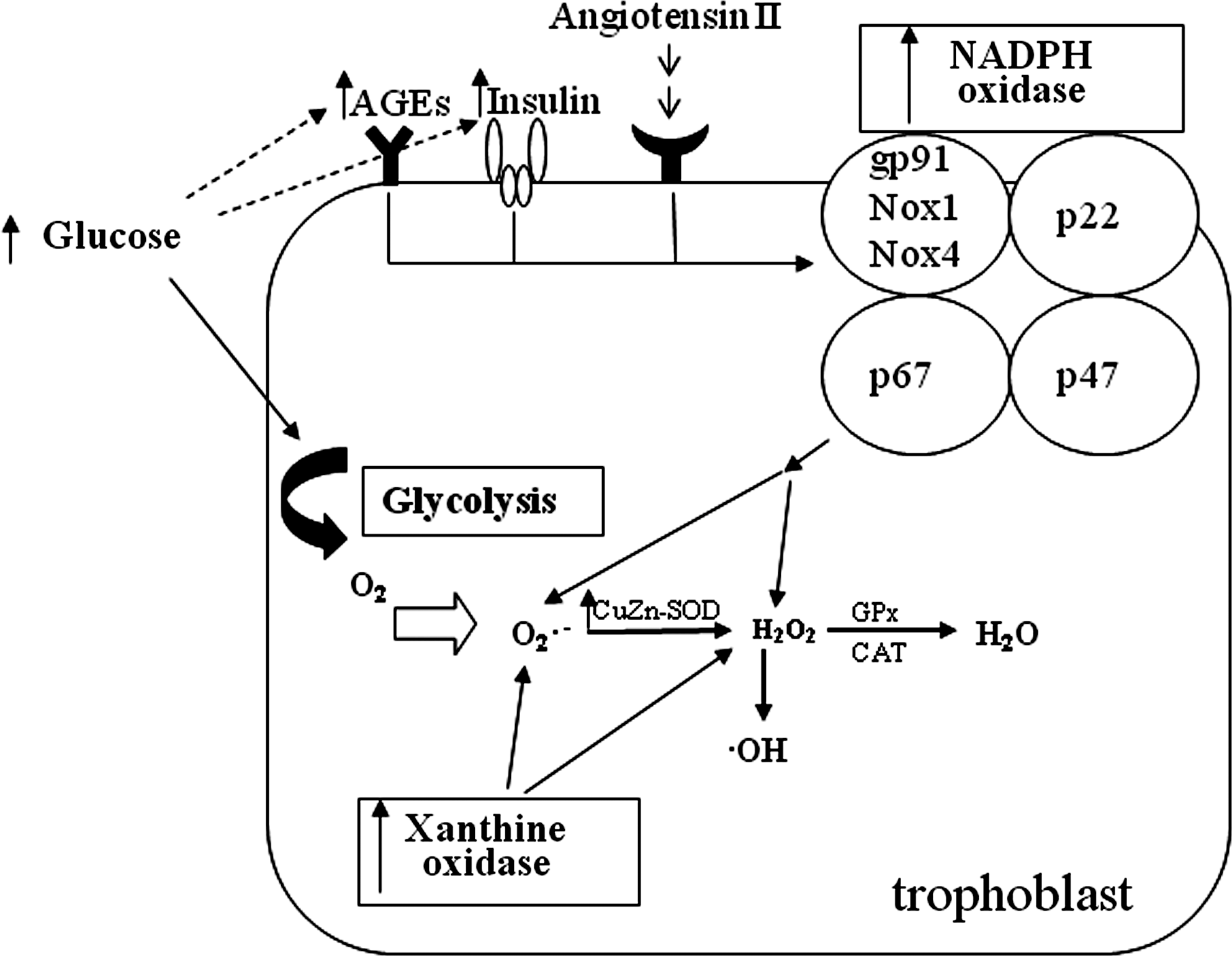
Under physiological conditions ROS is eliminated by cellular defense mechanisms, including diverse enzymes and vitamins. However, imbalance of ROS production and antioxidant systems of a cell can lead to an upregulation of antioxidant gene expression through activation of nuclear antioxidant response elements (ARE) by the redox-sensitive transcription factor nrf2 (106). Hyperglycemia causes excessive ROS formation, thus activating the Nrf2/ARE pathway (382). It appears that basal activity of NADPH oxidase provides ROS production to trigger Nrf2/ARE-mediated antioxidant gene expression to sustain redox homeostasis (236). NADPH oxidase was shown to be higher expressed and activated in endothelial cells of pregnant women with GDM (310). Expression of Nox1 was observed in the syncytiotrophoblast, in villous endothelium and in some stromal cell of the human placenta (73). This is paralleled by superoxide production via NADPH. On this basis, NADPH oxidase was suggested to represent the major enzymatic source of superoxide in the placenta (286).
E. Protein kinase C
PKC represents a family of highly homologous kinases, including several isoforms, which differ in their activation requirements and substrate specificities. Some isoenzymes (primarily β and δ) are activated by 1,2-diacylglycerol, a glycolysis intermediate compound, in the presence of phosphatidylserine. Activated PKC isoforms are capable to induce a variety of biological processes, that is, cell proliferation and differentiation, transmembrane ion transport, glucose and lipid metabolism, smooth muscle contraction, and gene expression (10, 31). Recently, a study has shown an activation of the PKC β2 isoform by hyperglycemia in heart and superior mesenteric artery as well as in cardiomyocytes of diabetic mice (260). PKC promotes the activation of mitochondrial NADPH oxidase, thereby leading to increased oxidative stress events. Once stimulated, NADPH oxidase reduces glutathione levels and impairs the cellular antioxidant defense systems (183).
F. Xanthine oxidase
Xanthine oxidoreductase (XOR) belongs to a group of enzymes known as molybdenum iron-sulfur flavin hydroxylases. It exists as two inter-convertible forms: XO and xanthine dehydrogenase (XDH). XOR is the rate-limiting enzyme in the conversion of hypoxanthine to xanthine and of xanthine to urate. XOR is present in vivo as XDH but it can be easily oxidized to XO. XDH does not lead to raised ROS production because of its greater affinity for NAD+ compared to oxygen molecules. During XO re-oxidation the enzyme transfers its six free electrons onto molecular oxygen causing the production of H2O2 and O2 − (Fig. 5). Observations in diabetic mice showed increased activity of XO in tissue and serum. Treatment with the XO inhibitor allopurinol reduced the activity of XO back to normal levels (297). Immunohistochemistry demonstrated XO expression in the human placenta (286). Higher levels of active XO were also detected in umbilical cord blood from fetuses of GDM pregnancies, suggesting fetal ROS formation in this condition (20).
G. ROS production via mitochondria
In most mammalian cells mitochondria are the major source of ROS. ROS are produced as a result of incorrectly coupled electron transport in the mitochondrial respiratory chain by oxidative phosphorylation. Once generated, ROS can either mediate mitochondrial damage or it can play a crucial role in redox signaling from the mitochondrion to the rest of the cell. Superoxide cannot move across the mitochondrial membrane. Therefore, it is converted to H2O2 by MnSOD. Subsequently, the hydrogen radical is formed that diffuses across the mitochondrial membrane. H2O2 itself can be further degraded to water by GPx. This represents the primary elimination process of ROS in mitochondria (395).
Normal human pregnancy is considered a state of enhanced oxidative stress. This is because there is increased metabolic activity in the mitochondria of the placenta, which generates ROS, superoxide generation from NADPH oxidase, as well as altered antioxidant scavenging capacity. The role of the mitochondria in the generation of oxidative stress in normal pregnancy has previously been reviewed (259). In general, in vitro hyperglycemia-induced ROS production is reduced in extra-placental tissues by an inhibitor of electron transport chain complex II, by an uncoupler of oxidative phosphorylation, by MnSOD, and by uncoupling protein-1 (UCP1), which is exclusively expressed in brown adipose tissue. These data suggest that the TCA cycle is the major source of high glucose-induced ROS production (263). Further, it was shown that elevated ROS levels caused by hyperglycemia are involved in morphological changes of mitochondria (388). However, the effect of hyperglycemia on mitochondrial ROS generation and its contribution to increased oxidative stress in GDM is not known.
H. Concluding comments
There are a number of pathways that may contribute to oxidative stress observed in the GDM placenta. In the placenta, ROS and RNS are an important source of growth and signaling factors, and are susceptible to ROS-mediated apoptosis. In the following section, the effect of oxidative stress on placental function is discussed.
VII. The Biological Role of Oxidative Stress on Placental Function in GDM Pregnancies
Key points:
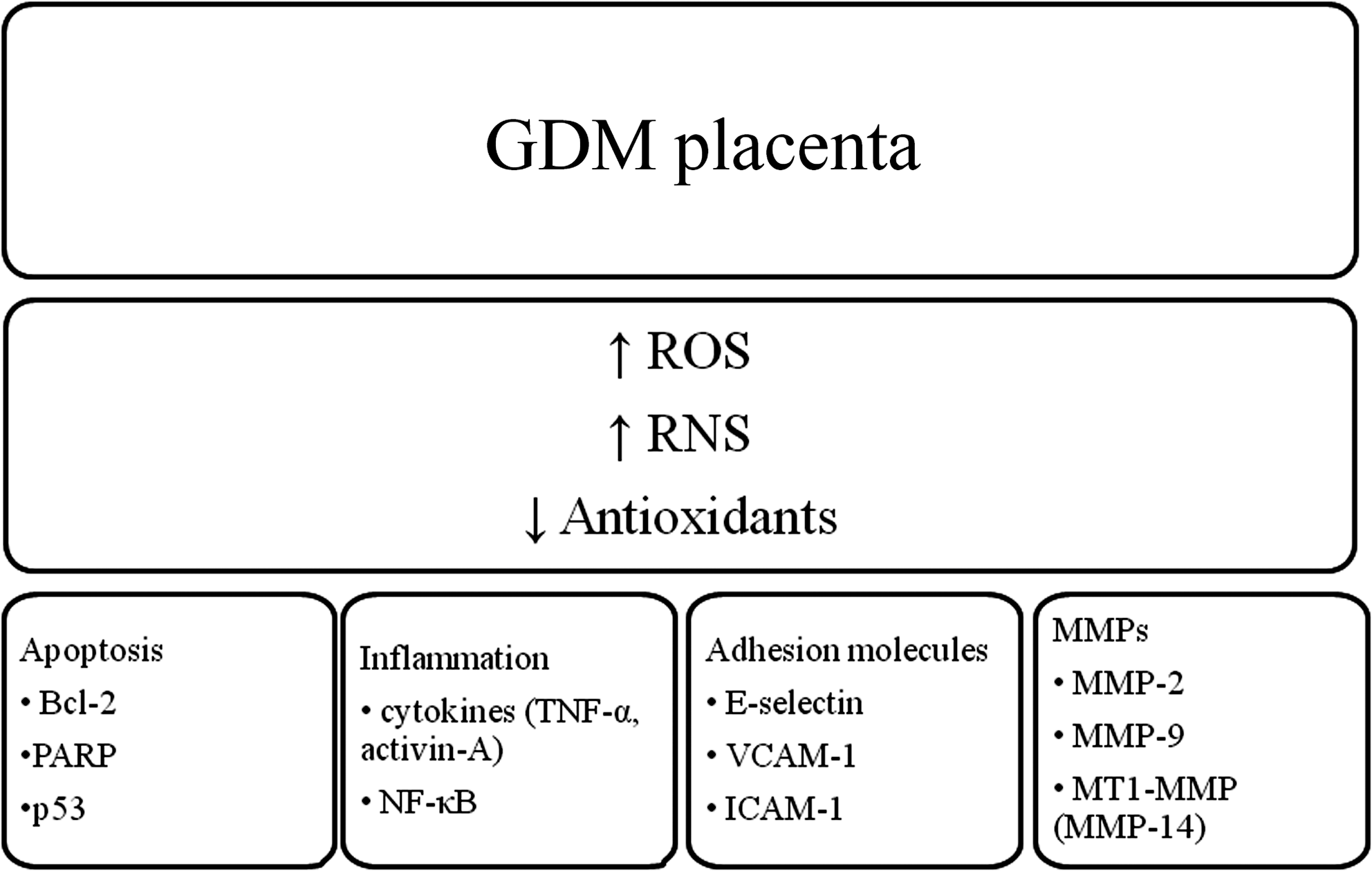
• Oxidative stress has a number of important roles, including regulation of pro-inflammatory cytokines, matrix metalloproteinases (MMPs), adhesion molecules, apoptosis, and the redox-sensitive transcription factor NF-κB.
• The biological functions of oxidative stress in pregnancy are summarized in Figure 6 and discussed below.
A. Inflammatory cytokines
Low-grade chronic inflammation is a central feature of GDM. Maternal systemic inflammation with increased concentration of adipokines and inflammatory cytokines is increased by the accumulation of functional macrophages in the interstitial stroma of both the placenta and maternal adipose tissue of women with GDM (17, 78, 185, 293). Although the initial stimulus triggering inflammation in pregnancy with GDM is not currently known, potential candidates include dietary and environmental factors such as caloric overload or changes in microbiota in the pregnant women (39, 214). Thus, the combined condition of inflammation and metabolic dysfunction of women with GDM may be regarded as a state of metabolic inflammation as proposed for other metabolic diseases associated with insulin resistance (145).
Data from our laboratories demonstrate that GDM placentas are less responsive to an oxidative challenge than placental tissue from normal women (70, 204). In normal placenta, oxidative stress induced a significant increase cytokine expression and release, an effect that was blunted in the GDM placenta (204). In marked contrast, adipose tissue obtained from women with and without GDM both respond to oxidative stress by increasing cytokine release. This may represent adaptive mechanism to protect the fetus from any further damage.
GDM is associated with elevated levels of activin A (281) and lower follistatin-like-3 levels, an inhibitor of activin A (344). The release of activin A, together with TNF-α, is considered to be one of the very first responses to inflammation (165). Oxidative stress induced by HX/XO increases placental and endothelial cell activin A secretion (234). Collectively, this suggests that oxidative stress associated with GDM may be a mechanism underlying the increased levels of activin A present in women with GDM.
B. Metalloproteinases
Placental development requires proper trophoblast invasion and tissue remodeling, processes involving MMPs—proteases that degrade various components of the extracellular matrix (ECM). Members of the MMP family include collagenases, gelatinases, stromelysins, matrilysins, and membrane-type MMPs (MT-MMPs). Oxidative stress is a potent activator of MMPs. Both ROS and NO can disrupt the cysteine switch that maintains the latency of proMMPs, leading to the activation of several different MMPs, including the gelatinases MMP-2 and MMP-9 (207, 231, 291, 292). H2O2 enhances MMP activity in the maternal side of the placenta and in the fetuses from control and diabetic rats (291). On the other hand, SOD reduces MMP activity in the maternal side of the placenta and in the fetuses from control and diabetic rats (291), and the antioxidant N-acetyl-cysteine (NAC) reduces MMP-9 activity in human placenta (207). Of note, placental MMPs are increased in the placenta from pre-gestational diabetic patients, in which both NO overproduction and peroxynitrite-induce damage has been found (188, 228, 288). MT1-MMP expression in first-trimester placental tissue is upregulated in type 1 diabetes and by TNF-α (138).
C. Apoptosis
Oxidative stress may trigger apoptotic cascades in the human placenta. For example, hypoxia-reoxygenation in vitro is a potent stimulus of apoptosis in the syncytiotrophoblast and that apoptosis can be modulated by the addition of antioxidants (59).
The available data demonstrate that GDM is associated with apoptosis. Specifically, a higher incidence of TUNEL-positive nuclei and lower expression of the anti-apoptotic protein Bcl-2 was reported in placental villous trophoblasts in GDM groups compared to the NGT pregnant women (316). Further, in GDM pregnancies, the leukocyte activity of the pro-apoptotic protein PARP is elevated as early as the middle of pregnancy (144); a positive linear correlation was observed between the severity of carbohydrate intolerance (the 2 h oral glucose tolerance test value) and PARP activity of circulating leukocytes. Additionally, hyperglycemia upregulates p53, triggering the mitochondrial death cascade pathway in the mouse placenta (251), and increases the rate of apoptosis in cultured trophoblast cell lines (363). Although alterations in RNS-induced S-nitrosylation have not been addressed in GDM, they might be related to the pathogenesis of GDM. Indeed, NO-induced S-nitrosylation affects key enzymes involved in the apoptosis cascade-like caspases and PARP (179, 320). Apoptosis effects of NO are highly dependent on the context and the levels of NO.
D. Vascular molecules
E-selectin, VCAM-1, and ICAM-1 are cell adhesion molecules expressed only on endothelial cells and as such play a role in the pathogenesis of vascular disease, which eventually leads to the development of atherosclerosis. Circulating E-selectin and VCAM-1 are increased in GDM (171). There is some evidence to show that oxidative stress activates the expression of E-selectin (294). However, there is limited data with respect to placenta and GDM.
Among the growth factors produced by trophoblasts, it is believed that vascular endothelial growth factor (VEGF) and its receptor family (including fms-like tyrosine kinase receptor [Flt-1]) play an important role in regulating trophoblast survival and angiogenesis in the placenta. Peroxynitrite have been shown to alter VEGF angiogenic signaling pathways (86). Hypoxia-induced increases in VEGF production and soluble Flt-1 (sFlt-1) expression have also been found in trophoblast cell lines (173, 219). The increased sFlt-1 production was positively correlated with increased lipid peroxide production (219). Increased sFlt-1 and VEGF release into maternal circulation could contribute to vascular endothelial dysfunction.
E. Nuclear factor-kappa B
NF-κB is an ubiquitous and inducible transcription factor that is a central regulator of immune and inflammatory responses, cell adhesion, differentiation, redox metabolism, and apoptosis. More comprehensive reviews on the role and regulation of NF-κB in human pregnancy can be read at (212, 300).
Oxidative stress stimulates NF-κB translocation into the nucleus, thus inducing up-regulation of genes associated with inflammatory response. Our previous studies have shown that in human placenta, NF-κB is activated in response to oxidative stress (70, 207) and this is associated with increased expression of pro-inflammatory cytokines, prostaglandins, and MMPs (70, 207, 213, 300). Likewise, antioxidants such as NAC can attenuate oxidative stress NF-κB activation. This positive regulatory loop may amplify and perpetuate local inflammatory reactions. However, GDM is associated with decreased placental NF-κB DNA-binding activity (70), suggesting that a regulatory mechanism may exist in GDM placenta.
Low concentrations of NO control NF-κB by activation of IκB kinases, whereas high concentrations inhibit NF-κB by increasing the stability of the NF-κB inhibitor IκB (169). However, it is of note that iNOS can both up- and down-regulate NF-κB independent of IκB as well.
F. Concluding comments
Growth factors, cytokines, MMPs, and apoptosis play important roles in placental structure and function. ROS leads to chronic inflammation, dysregulation of MMPs, and apoptosis. The sequelae of elevated ROS on trophoblast health, which may influence development of conceptus, are discussed below.
VIII. The Role of Oxidative and Nitrative Stress in the Pathogenesis of GDM
Key Points:
• Reactive oxygen and nitrogen species play a number of important roles throughout pregnancy, including embryo development, implantation, angiogenesis, placental development and function, and thus fetal development and subsequent adult diseases.
• The involvement of oxidative and nitrative stress in impairing developmental processes in diabetic pregnancies is discussed below.
• The involvement of oxidative and nitrative stress in altering placental function and transport in diabetic pregnancies is also discussed below.
A. Decidualization and implantation
Diabetes in pregnancy is associated with suboptimal decidualization (110). Implantation involves the interaction between several vasoactive agents, including cytokines, prostaglandins, and NO, which lead to increases in MMPs (158, 268, 345). Indeed, NO plays a key role in decidualization and embryo implantation (265). It increases vascular permeability, vasodilation, and blood flow in the uterus, and is a component of the decidual cell reaction (345, 353). In rat decidua, when NO is inhibited, apoptosis is increased, suggesting that NO plays a role in survival of decidual cells (338). In the rat uterus, NO increases during the peri-implantation days in rats, and diminishes after the implantation period (158). Accordingly, eNOS and iNOS expression, and NOS activity are enhanced during the peri-implantation days of pregnancy (266, 315). Implantation, however, occurs at normal rates in the diabetic rats; it is thought that implantation under these pro-inflammatory conditions may be involved in the increased resorption and malformation rate evident in experimental diabetes and pregnancy models (158, 267).
The early steps of decidualization and implantation should not be affected when GDM is induced in the second trimester of pregnancy, but, as stated in other sections, if glucose intolerance and its related changes in NO and ROS production are present in the first trimester of pregnancy, this may affect these early processes, impairing the initial steps of embryonic and placental development.
B. Trophoblast invasion
Normal placentation requires trophoblast invasion of maternal spiral arteries, and development of a high-flow, low-resistance uteroplacental circulation. Trophoblast invasion is influenced by a number of factors such as cytokines and growth factors, adhesion molecules, MMPs, and oxygen tension (224, 375). As detailed in Section VII, oxidative and nitrative stress can regulate these factors, which may then play an important role in abnormal trophoblast invasion. Certainly, the ability of the trophoblast to invade the uterus is related to NO production during implantation and during the remodeling of uteroplacental arteries (170, 345). Trophoblast-derived NO seems to serve to dilate the vessels and turn the uterus receptive to trophoblast penetration. The role of eNOS in the trophoblast is highlighted by the presence of this NOS isotype in villous and labyrinthine cytotrophoblasts and the syncytiotrophoblast, feto-placental endothelium, cell columns of anchoring villi, and invasive cytotrophoblasts (353). On the other hand, NO overproduction can also induce trophoblast apoptosis, although this may rely on iNOS-derived NO (390). Indeed, iNOS, the NOS isotype mostly related to NO overproduction, is also expressed in the placental microvascular endothelium, syncytiotrophoblast, and cytotrophoblast cells, and has been found to be involved in alterations in feto-placental circulation in pathological situations (91).
Interestingly, nitrosylation of MMPs at the edge of migrating trophoblasts has been found associated with an increase in iNOS expression and related to the process of trophoblast invasion (130). All this suggests that trophoblast invasion and remodeling of uteroplacental arteries are processes highly related to the mature placental function that can be highly affected by hyperglycemia-induced oxidative and nitrative stress.
C. Organogenesis
As detailed in Section IV, diabetes during pregnancy is associated with embryonic dysmorphogenesis. Due to its capacity to regulate cell survival, apoptosis, differentiation, and ECM remodeling, oxidative and nitrative stress play a significant role in embryo organogenesis. Indeed, low and high levels of NO can leads to embryonic maldevelopment, possibly due to an improper regulation of apoptotic events, which should occur in appropriate space and temporal location to allow the formation of the organs (216, 285). Further, a particular pattern of expression of NOS during organogenesis supports its role as a morphogen (22, 349, 387).
During embryo and fetal development, NO has been found to be relevant in regulating differentiation of cell types (e.g., cardiomyocytes and neuronal cells) and organ formation (e.g., lung branching morphogenesis, cephalic morphogenesis, heart development, and nephrogenesis) (22, 97, 283, 340). Fetal growth is also regulated by NO, and fetal growth is restricted in the presence of NOS inhibitors (262).
The higher 8-isoprostane levels observed in the offspring of diabetic animals (368) have its own teratogenic potency (366), thus supporting a link between oxidative stress and increased malformation rate in embryos exposed to a diabetes-like environment. Diabetic embryopathy is also associated with inhibition of GAPDH activity and GAPDH gene expression resulting from an excess of ROS in the embryo (365), an effect that can be restored by treatment with the antioxidant NAC.
Impaired induction of transcription factors, such as paired box (PAX)-3 and peroxisome proliferator-activated receptor (PPAR) δ, has been found to be involved in the induction of both neural tube and heart malformations, the most common malformations induced by maternal diabetes in both humans and experimental models of diabetes (139, 194, 220). The impaired induction of PAX-3, which leads to an increase in apoptosis of neural crest cells and an impairment of the process of neural tube closure and neural crest cell migration, is clearly related to increases in oxidative stress. Indeed, different antioxidants such as α-tocopherol and gluthatione ethyl ester increase expression of PAX-3 and prevent apoptosis and the induction of hyperglycemia-induced neural tube and heart defects (54, 254).
In summary, there is clear evidence of the capacity of oxidative and nitrative stress to impair embryo organogenesis. Thus, if glucose intolerance and its related changes in NO and ROS production are present in the first trimester of pregnancy, organogenesis can be profoundly affected.
D. Endothelial and vascular dysfunction
Placental development is associated with significant increases in both angiogenesis and vasodilatation, giving rise to a dramatic elevation of placental blood flow during pregnancy. This increased blood flow is directly correlated with fetal growth and survival as well as neonatal birth weights and survivability. ROS and RNS can modulate angiogenesis, vasculogenesis, and vessel reactivity in the feto-placental circulation. Studies performed in ewes have been helpful to understand the role of NO in controlling vasodilation during pregnancy. NO production is increased in ewes carrying multiple fetuses compared to singletons, possibly due to the increased demand of uterine blood flow to sustain multiparity and to potentially aid in the remodeling of the vascular bed (360). The reduction of NO production through the inhibition of NOS leads to a reduction in uterine blood flow in pregnant ewes (304). Similarly, NO production is increased in pregnant rats, as evidenced by the increased plasma and urinary levels of nitrates/nitrites, and the increased urinary levels of cGMP (65). Interestingly, pregnant rat inhibition of NO leads to pre-eclampsia-like effects (383). Formation of peroxynitrites, generated from NO and ROS, reduces NO bioavailability. Indeed, in perfusion studies, feto-placental vasoreactivity has been shown to be clearly altered by the addition of peroxynitrites (188). These alterations are relevant when addressing the effects of RNS in GDM, as summarized in Figure 7 and described below.
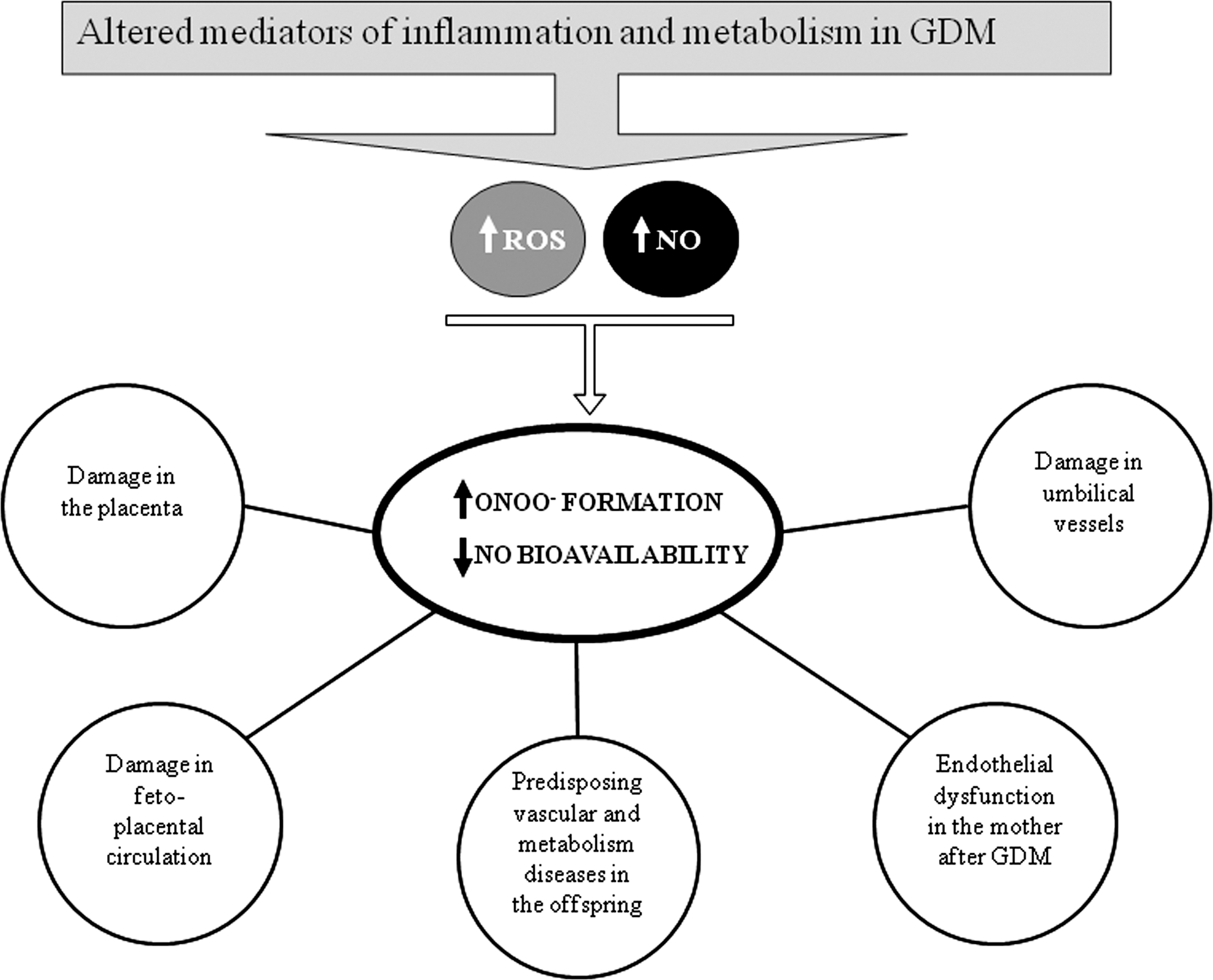
Several alterations in both maternal and fetal endothelial dysfunction have been observed in GDM patients (43, 257, 308). Considerable evidence exists that oxidative stress plays an important role in endothelial dysfunction. As maternal hyperglycemia leads to fetal hyperglycemia and exposure to hyperglycemia leads to AGE formation, oxidative stress will be induced in the feto-placental endothelium. In support, 8-isoprostane is capable of inducing vasoconstriction in the placenta (217). Thus, placental secretion of 8-isoprostane into the maternal circulation could contribute to vasoconstriction observed in GDM. Nitrative stress is also likely to increase nitration of proteins in GDM, leading to endothelial and vascular dysfunction. NO produced by eNOS in the endothelial cells may interact and form peroxynitrite, which covalently modifies proteins and DNA. In a pregnancy complicated by diabetes, eNOS expression is increased in placental arteries, veins, and HUVECs (11, 95, 356). Moreover, expression of transporters for L-arginine (the precursor of NO synthesis) is increased in microvascular placental endothelial cells and HUVEC from GDM pregnancies (328), further promoting NO synthesis. Excess generation of NO and ROS may therefore stimulate peroxynitrite formation. Elevated levels of placental nitrotyrosine residues have been shown in type I diabetes, insulin-treated GDM patients, and experimental models of diabetes (144, 228, 371). The resulting nitration of proteins will alter protein function, resulting in vascular damage and dysfunction.
Fibrosis is characterized by excessive ECM deposition in various organs and the vasculature and often by a change in the quality of the ECM, as well as angiogenesis. There is certainly evidence to show that diabetes is associated with fibrosis in a number of tissues (12), and that GDM is associated with placental angiogenesis (163). Thus, by activating MMPs (207, 231, 291, 292), oxidative stress may also induce a fibrotic response and disrupt the structural integrity of placental endothelial cells, and influence angiogenesis by degrading matrix molecules, loosening the cellular network, and releasing growth factors sequestered in the ECM. Further, the oxidative stress-mediated increases in adhesion molecules may cause the endothelial cell dysfunction and atherosclerosis observed in GDM (7, 129).
Collectively, the changes described show profound alterations in the placental and umbilical vasculature and dysfunction of endothelial cells derived from excessive oxidative and nitrative stress. These changes, together with changes in maternal metabolites and impairments in placental transport function (described in Section F), will lead to complex alterations in the quality and quantity of nutrients and in the oxygen available to the fetus in GDM.
E. Placental nutrient transport mechanisms
Pregnancy requires specific adaptations of maternal nutrient metabolism (i.e., carbohydrate, lipid, and protein) to meet the increase energy needs of the mother and growing fetus. These adaptations are exacerbated in women with GDM (306). As the primary target of maternal environment, the placenta is naturally exposed to all modifications of maternal homeostasis. The maternal metabolic changes have been long documented to affect placental transport function and metabolism of energy nutrients in women with GDM (80, 151). The potential impact of oxidative stress on glucose, lipid, and amino acid metabolism in the placenta is discussed below.
1. Glucose transport
a. Glucose metabolism in pregnancy
To sustain continuous growth and development of the fetus, the maternal metabolism changes with and adapts to pregnancy. The first trimester of pregnancy is characterized by higher insulin sensitivity, which augments the anabolic effects of insulin, ultimately allowing the mother to build up energy and nutrient stores for the second and third trimester (48, 186). When gestation advances, rapid growth of the fetal tissue increases its nutrient demand and the anabolic glucose metabolism shifts to a catabolic state. Glucose is the main energy source for both the fetus and the placenta and, hence, has to be easily available. Therefore, in sharp contrast to the first trimester, in the second half of pregnancy maternal metabolism shifts into an insulin resistant state resulting in decreased glucose uptake by insulin target tissues of the mother. This facilitates trans-placental glucose transport to the fetus and enables adequate glucose supply of 30–50 g glucose per day (133).
In normal pregnancy, the increase in insulin resistance is compensated by a concomitant increase in insulin production, which is the result of hypertrophy and hyperplasia of β-cells (355). In patients with GDM, insulin resistance is either comparable or greater than that in nondiabetic pregnancy, whereas insulin secretion appears to be compromised. Failure to adequately compensate insulin resistance (32) ultimately leads to maternal hyperglycemia. Although β-cell dysfunction seems to be the principal cause of GDM, higher levels of leptin (172), and higher villous secretion of TNF-α in response to glucose (68) have been reported. Both are inhibitors of insulin signaling at the receptor level and may be responsible for the maternal peripheral insulin resistance.
Along its transport-route through the placental barrier into the fetal blood, glucose has to pass the syncytiotrophoblast layer, which covers the placental villi. In segments of the syncytiotrophoblast characterized by exchange activity, fetal capillaries are located underneath the syncytiotrophoblast. Glucose transport through plasma membranes is accomplished by a family of glucose transport proteins designated as glucose transporters (GLUTs), which facilitate glucose uptake. They enable glucose transport along a concentration gradient by facilitated, carrier-mediated sodium-independent diffusion (9). Different GLUT isoforms are expressed in a tissue-specific manner and vary in their regulation and substrate specificity. GLUT-1 is the ubiquitous transporter involved in the trans-placental movement of glucose and is expressed in the apical and basal membrane of the syncytiotrophoblast as well as in all other cell types of the villi. Various other GLUTs, such as GLUT-3 (132), GLUT-4 (381), GLUT-8 (120), and GLUT-12 (123), have also been identified within the placenta. However, the localization of GLUT-3, -4, and -12 in cells of the placental endothelium or the villous stroma makes their involvement in trans-placental glucose transport unlikely (87). Once having traversed, the syncytiotrophoblast glucose will pass through the paracellular clefts between the endothelial cells that line the feto-placental vessels. Hence, the two membranes of the syncytiotrophoblast represent the placental barrier for glucose (Fig. 8). GLUTs expressed on the endothelial cells will serve to cover the energy requirements of the endothelial cells. Presence of high-affinity GLUT-3 on the endothelial cells ensures energy supply even in situations of low glucose levels.

The placenta is a metabolically highly active organ and uses a considerable part of the glucose derived from the mother for itself. Only 40%–50% of the glucose taken up is released to the fetus, whereas the rest is metabolized by the placenta. Oxidative glucose metabolism is low and about 80% of the glucose used by the placenta is converted to lactate, which is released to the maternal and fetal circulation, or stored as glycogen mostly in placental endothelial cells and pericytes (79, 81, 164). The placental processing of glucose and its downstream metabolites uses involves and affects a variety of metabolic pathways. Therefore, an increase in glucose availability may induce multiple cellular changes. Maternal and fetal hyperglycemia may result in the activation of glycolysis, one of the key metabolic pathways of glucose metabolism and will lead to production of adverse side products or to an accumulation of metabolic intermediates in the placenta. The glucose gradient as a key determinant of flux has an important clinical consequence. Adequate glycemic control of the mother alone does not avoid hyperglycemia of the fetus. Fetal hyperinsulinemia stimulates fetal glucose metabolism, steepens the gradient, and results in glucose stealing from the mother (264).
The effect of oxidative stress on placental glucose metabolism is not known. However, in nongestational tissues, there is certainly plenty of evidence demonstrating oxidative stress regulates GLUT-1 and/or GLUT-3 dependent glucose uptake and transport. Thus, we suggest that oxidative stress may also positively or negatively regulate glucose metabolism in human placenta, and this would have implications for the amount of glucose that is being delivered to the fetus (Fig. 9). On the other hand, we suggest that the GDM placenta will be less sensitive to oxidative stress due to the heightened level of antioxidants. In keeping with this, preliminary data from the Lappas group supports this hypothesis. Specifically, we have shown that the NGT placenta decreases glucose uptake in response to HX/XO, an effect that can be restored by the co-incubation with antioxidants. On the other hand, in the GDM placenta, there is no effect of HX/XO on glucose uptake (Fig. 9). Although it is not known what effect HX/XO has glucose transport to the fetus, our preliminary data show that HX/XO increases glucose transport in placental BeWo cells (Lappas, personal communications). Overexpression of thioredoxin-1 reduces oxidative stress in the placenta of mice and promotes fetal growth by increasing placental glucose availability through up-regulation of GLUT-1 expression (351). Collectively, these data suggest that the placental antioxidant systems play a role in influencing fetal growth.

b. Insulin signaling in GDM
Several studies investigated GDM-associated changes in insulin signaling components in maternal muscle and adipose tissue samples, classical target tissues of insulin-stimulated glucose uptake (Table 3). Basically, the changes observed were similar between adipose tissue and skeletal muscle. In tissues, decreased expression of insulin receptor substrate (IRS)-1 and GLUT-4 as well as increased phosphatidylinositol 3-kinase (PI3K) p85 expression was shown. Binding of only the p85 subunit of PI3K to IRS-1 prevents binding of the PI3K signaling complex composed of p85/p110 heterodimers. Hence, upregulation of p85 PI3K will decrease the PI3K-mediated IRS-1 signaling. A study investigating long-term changes in skeletal muscle insulin signaling after a GDM pregnancy (102) found elevated TNF-α expression, reduced insulin receptor auto-phosphorylation, and serine phosphorylation of IRS-1 one year postpartum. This indicates that insulin resistance once commenced in pregnancy is a long lasting condition.
=no change.
↓decreased in GDM.
↑increased in GDM.
GLUT, glucose transporter; PI3K, phosphatidylinositol 3-kinase; IRS, insulin receptor substrate.
Maternal GDM, but not obesity, is associated with reduced levels of IRS-1 and GLUT-4 also in the placenta. In contrast to maternal adipose tissue and muscle, protein expression of placental p85 PI3K was decreased (63). Oxidative stress has also been shown to reduce IRS-1 and GLUT-4 expression by protein degradation (IRS-1) and reduction of gene expression (GLUT-4). This suggests that the GDM-associated changes in maternal tissue and placenta are a result of oxidative stress.
Recent data showing the physiological and pathological function of oxidative stress in insulin signaling and glucose metabolism indicate an even tighter interplay of hyperglycemia and insulin resistance than thought for long (Fig. 10). Under normal conditions, physiological levels of ROS promote and stimulate adequate insulin signaling. This is also reflected by the finding that addition of antioxidants inhibits insulin signaling (313). Most likely, this is because the insulin signaling pathway leads to low levels of ROS production itself and ROS act as second messengers of which disposal impairs insulin signaling. Insulin-induced ROS production is accounted for by activation of the NADPH oxidase NOX4 through PI3K. The ROS pathway subsequently activates kinases or induces gene expression by redox-sensitive transcription factors.
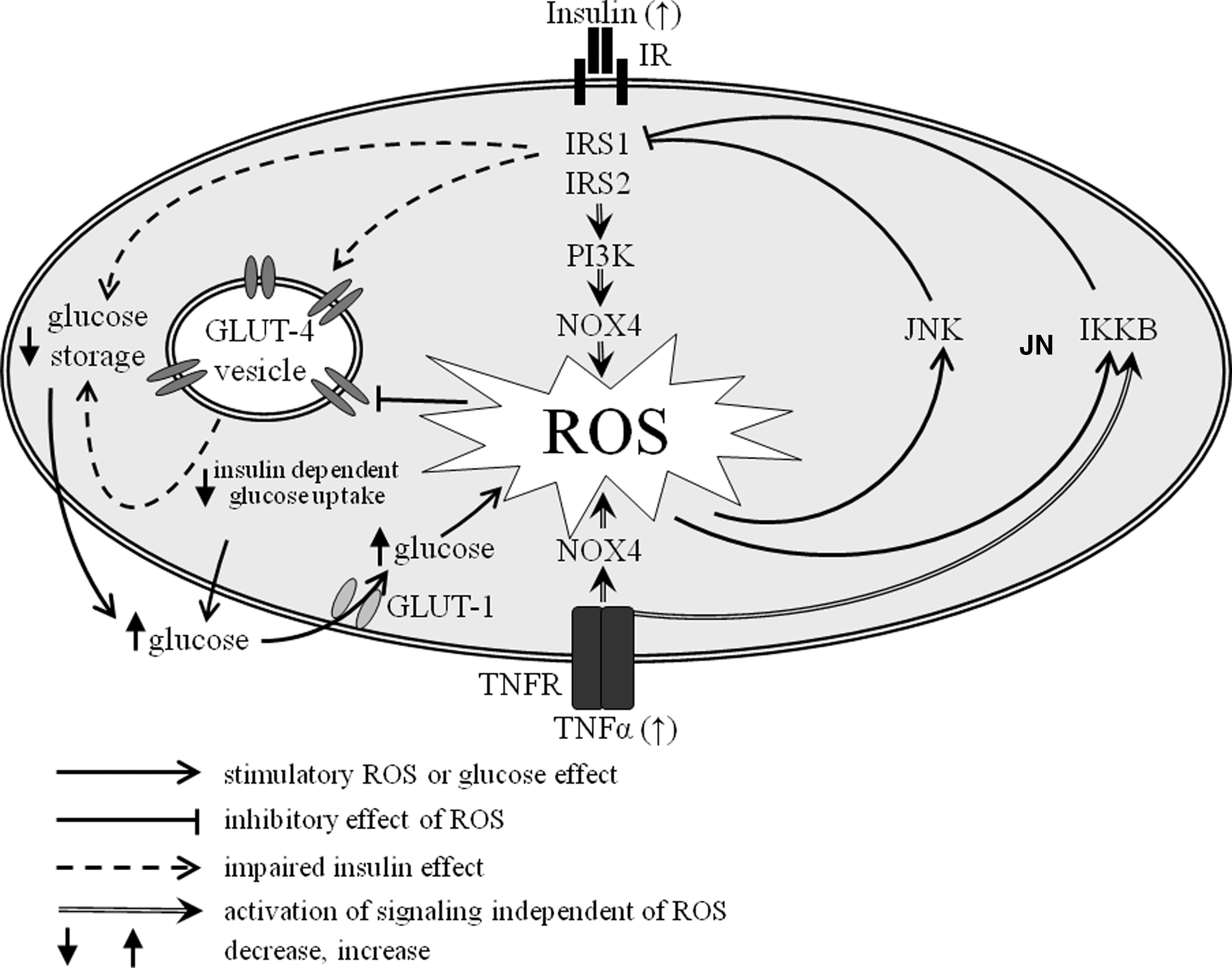
Chronic hyperinsulinemia as present in obesity and pre-diabetes may cause increased ROS generation and result in impaired insulin signaling, insulin resistance, and oxidative stress (112). Additionally, in the hyperglycemic environment of diabetes, excessive, pathological ROS levels overwhelm the ROS levels generated by and required for insulin signaling. This leads to attenuated insulin signaling. The stress responsive c-Jun N-terminal kinase (JNK) as well as the inhibitor kappa b kinase complex (IKKβ) become activated by ROS and phosphorylate IRS-1 at serine and threonine residues, thus inhibiting its function and insulin receptor signal transduction (3, 108). Inhibition of IRS-1 leads to reduced GLUT-4 translocation and glycogen synthesis in response to insulin. This is further reinforced by decreased GLUT-4 transcription in response to ROS (280). Impaired GLUT-4 translocation lowers insulin-stimulated glucose uptake by muscle and adipose tissue, and glucose storage as glycogen, hence increasing extracellular glucose levels. Virtually, all cells express insulin-independent GLUTs for their own energy supply. This basal glucose uptake is accomplished mainly by GLUT-1 and -3. Thus, resulting from facilitated diffusion of glucose through these transporters, extracellular hyperglycemia increases also intracellular glucose. High glucose, however, contributes to ROS formation, which in turn will impair insulin signaling (Fig. 10). The levels of TNF-α are higher GDM, which may also contribute to the accelerated ROS production by increasing NOX4 expression (235) and activation (18). Hence, not only hyperglycemia, but also the inflammatory conditions of diabetes, can induce insulin resistance, thereby creating a vicious circle, that is, promoting hyperglycemia and failure of glucose lowering insulin action.
As detailed above, GDM is associated with increased placental oxidative stress. GDM is also associated with higher levels of glycogen deposited around the feto-placental vessels (164), demonstrating higher levels of intracellular glucose in GDM. The effects of oxidative stress on placental glucose metabolism itself, however, remain elusive. In recent studies, we have demonstrated that pro-inflammatory cytokines, which we have previously shown to exhibit pro-inflammatory and pro-oxidative effects in placenta and adipose tissue and are increased in GDM pregnancies (208), increase GLUT-4 mediated insulin signaling and thus glucose uptake in placenta, whereas in adipose tissue they cause insulin resistance (Lappas M, personal communications). Adipose tissue insulin resistance makes glucose available for uptake by the placenta and thus potentially more glucose available for transport to the fetal circulation, resulting in larger babies.
2. Fatty acid transport
The highest growth rate in a human life span occurs during the fetal period. Deposition of fat stores in the fetus is high, where body fat growth occurs essentially during the last trimester of intra-uterine life. During this rapid growth, the fetus requires significant amounts of fatty acids. The fetus has an absolute requirement for the n-3/n-6 fatty acids, and docosahexaenoic acid (22:6 n-3; DHA), in particular, is essential for proper development (127, 135). Fatty acids are needed as structural components of membrane phospholipids, precursors of important bioactive compounds (such as the prostacyclins, prostaglandins, thromboxanes, and leukotrienes), a source of energy, precursors to signaling molecules, and are critical for organogenesis.
Pregnancy is characterized by changes in maternal adiposity and thus changes in lipid metabolism. There is an accumulation of maternal fat stores in early and mid pregnancy (anabolic phase), and enhanced fat mobilization in late pregnancy (catabolic phase). In late pregnancy, the overall proportion of maternal fat oxidized is reduced, and plasma maternal circulating concentrations of triacylglycerols (TAGs), phospholipids, nonesterified fatty acid, and glycerol are increased. This implies that the mobilization is to increase the availability of fatty acids for the fetus. The cellular mechanisms that trigger the transition from lipid storage to increased lipolysis during pregnancy are unknown; however, current dogma suggests an important role for insulin (84) and placentally derived hormones (218). In early pregnancy, increased estrogen and progesterone, and increased insulin sensitivity favor lipid deposition and inhibit lipolysis. On the other hand, late pregnancy hyperinsulinemia and insulin resistance promote the mobilization of fat stores, which is responsible for the hypertriglyceridemia of pregnancy (323). In obese pregnant women and women with GDM, peripheral insulin resistance is even more pronounced (62), and changes in hepatic and adipose metabolism alter circulating concentrations of TAGs, fatty acids, cholesterol, and phospholipids (243). GDM is accompanied by a threefold increase in plasma TAG concentrations during the third trimester of pregnancy, elevation of plasma postprandial fatty acids, delayed postprandial clearance of fatty acids, and elevation of the branched-chain amino acids (136, 147).
The effect of GDM on placental lipid metabolism is poorly understood. The insulin resistance of GDM and obese mothers may be a potential factor to enhance substrate availability to the fetus, which may either result in immediate (i.e., large baby) or long-term consequences (i.e., obesity and/or diabetes in later in life). The availability of fetal energy substrates is regulated in the first place by their maternal circulating concentrations and to the extent that they are transported across the placenta (Fig. 11). Whether an excess of placental lipids are mobilized from maternal adipose tissue and exported into the fetal circulation is critical information. Further, in placenta, there are three coordinated pathways that control the amount of fatty acids that are available for the fetus in response to GDM: (i) rate of fatty acid uptake from maternal circulation; (ii) esterification/storage capacity; (iii) mobilizing activity for export. In well-controlled GDM pregnancies, maternal lipids are strong predictors for fetal lipids and fetal growth. Infants with abnormal growth seem to be exposed to a distinct intrauterine environment compared than those with appropriate growth (312).
The effect of oxidative stress on placental lipid metabolism is not known. However, recent data highlight important roles for pro-inflammatory cytokines in the regulation of lipid metabolism in placenta, which may be expected to facilitate the transfer of lipids through the placental barrier to the fetus. Leptin induces a reduction in triglyceride levels through mechanisms that do not involve a reduction in de novo lipid synthesis (372), and in cultured primary human trophoblast cells, IL-6 stimulates fatty acid accumulation (197). We hypothesize that oxidative stress-induced pro-inflammatory cytokines may regulate placental lipid metabolism (Fig. 12).

3. Amino acid transport
Alterations in placental amino acid transport may contribute to accelerated fetal growth in pregnancies complicated by diabetes (152). This results in increased uptake of neutral amino acids across the microvillous membrane, which may be used in placental metabolism, or be delivered to the fetus to contribute to accelerated fetal growth in these patients.
Together with the increased NO production, L-arginine, substrate from NO synthesis, is increased in plasma from GDM patients, and L-arginine transport is increased in umbilical vein endothelial cells from GDM patients (37, 329, 330). In addition, in umbilical vein endothelial cell cultures from GDM patients the concentration of adenosine, a purine nucleoside that regulates vascular tone in the placenta, is increased (299, 356). Further to this, a very recent study has also shown that umbilical vein blood adenosine concentrations are higher in GDM (370). As profoundly addressed and reviewed elsewhere (91, 308, 369), a complex interaction between adenosine, its receptors from the equilibrative nucleoside transporters family, and the NO substrate L-arginine, is involved in the regulation of NO synthesis in the umbilical circulation and the placental endothelial microvasculature. Adenosine-induced NO/endothelium-dependent umbilical vein relaxation was lower in GDM (370). Changes in this delicate system, together with those related to NO reactivity with ROS, lead to an impaired NO synthesis and bioavailability in GDM (259, 308). Adenosine acts as an antioxidant (232); thus, the increase in adenosine in umbilical veins of GDN patients suggests an important protective role.
GDM is a state of insulin resistance by the fetus. Recent studies have shown that there are defects in its biological actions in the placenta from GDM and normal pregnancies. For example, in HUVECs, insulin restores the decrease in adenosine transport induced by GDM, an effect that could be blocked by the NOS inhibitor L-NAME (370). Similarly, insulin induces HUVEC relaxation by increasing HUVEC L-arginine transport (119). Collectively, this suggests that insulin acts to protect against endothelial dysfunction, a characteristic of GDM.
It is well known that after pregnancy, GDM predisposes to type 2 diabetes. As endothelial dysfunction has been considered a putative causal role in insulin resistance (284), endothelial dysfunction has been studied in women with previous GDM. Studies have revealed endothelial dysfunction in forearm conduit arteries and skin microvasculature of women with previous GDM (7, 129). Besides, formation of asymmetrical dimethyl-L-arginine, a product of the methylation of L-arginine that competitively inhibits cellular L-arginine uptake and NOS activity in endothelial cells (61), is also increased in sera from patients with previous GDM (249).
In addition to L-arginine, there are a number of other amino acid transport systems that are altered in GDM (154). Neutral amino acid transport capacity by System A is increased in both GDM and type I DM with large for gestational age babies (152). In addition to this, leucine is also increased in GDM placentas (152). It is therefore interesting that ROS reduces amino acid activity (176), perhaps representing an adaptive response where substrate availability may be increased. Collectively, the above data suggest that amino acid transport is a key regulator of fetal growth.
4. Placental ion transport mechanisms
Oxidative stress in human pregnancy may be linked to dysregulation of ion transport mechanisms, which allow the transport of ions into the placenta. It is well established that ROS has deleterious actions on a number of ion transport mechanisms, including ion channels (including Ca2+, K+, and Na+ and Cl− channels), transport enzymes, ion exchangers, and other transporters (189). However, ROS and a peroxidised lipid environment inhibit the activity of the transient receptor potential (TRP)-type Ca2+-permeable nonselective cation channel polycystin-2 (PC2) in term human syncytiotrophoblast (253). It is hypothesized that the inhibitory function of ROS on PC2 activity may play an important regulatory role by preventing a Ca2+ overload in stressed cells in the placenta. In support of this hypothesis, hypoxic placental villous fragments have increased TBARS and reduced plasma membrane Ca2+-ATPase activity, as compared to normoxic villous fragments (28). Collectively, the above data are consistent with neonates of women with GDM being associated with hypocalcinemia.
F. Cervical ripening and labor
At term, animal and human studies have shown that NO has the ability to induce cervical ripening, which occur as a result of the close relationship between prostaglandins and NO production (352). Indeed, cervical release of NO is increased during labor, and NO induces relaxation of cervical smooth muscle and stimulates prostaglandin production (118). NO overproduction has been found related to the mechanisms of induction of septic abortion (4). Likewise, ROS has also been implicated in the mechanisms of human labor. Oxidative stress is increased in placental tissues after labor (60), which may be related to its ability to induce NF-κB activation (207). Thus, it is tempting to speculate that the increase in oxidative stress observed in women with diabetes may explain why they have higher rates of spontaneous preterm labor (242). In keeping with this, as detailed in Section III, iron concentrations are higher in women with GDM (1, 24, 200 –202, 331), and iron-dependent oxidative stress has been linked to the pathophysiology of preterm labor.
G. Intrauterine programming
The in utero environment in which a fetus grows and develops may have long-term effects on subsequent health and survival. Intrauterine programming of metabolic, cardiovascular, and renal disease has been addressed in experimental models of diabetes and pregnancy (98, 162). Cardiac, vascular, and renal dysfunction in adult offspring from diabetic pregnant rats reflects impairments in NO synthesis and decreased endothelium-dependent vasodilatation, although preserved vascular response to exogenous NO (143, 302). Maternal diabetes-induced hypertension in rat offspring is prevented by supplementation with L-arginine, a treatment that also improves glomerular hypertrophy (50).
GDM also has long-term implications for the mother. Interestingly, in a recent study performed in a mouse model of GDM, impaired endothelial vascular reactivity has been found in the mothers after GDM, an alteration related to increases in superoxide production and peroxynitrite formation in the vessels (333). These data suggest that GDM affects endothelial function and may contribute to an increased risk of cardiovascular disease later in life.
H. Concluding comments
Oxidative stress disturbs placental function, leading to perpetrations in fetal growth and development. Thus, of vital important is the management of oxidative stress in women with GDM. In the following section, we will discuss the therapeutic approaches that may be of benefit in these pregnancies.
IX. Effects of Therapeutic Approaches on GDM
Key points:
• Evidence suggests that by decreasing oxidative stress, we may be able to reduce or even ameliorate oxidative stress-induced impacts on the fetus of GDM women.
• In diabetic animal models, antioxidant treatment has been shown to be effective in reducing the deleterious effects of GDM on the offspring.
• There are only a couple of clinical studies evaluating the potential benefits antioxidants in GDM women. These studies have not evaluated the effects on the fetus.
• Alternative approaches (including flavonoids or PPAR agonists) may be of potential benefit, as their efficacy in the treatment of a variety of diseases has been demonstrated in numerous clinical studies.
A. Can antioxidant treatment reduce oxidative stress in GDM?
There is a higher incidence of congenital malformations in the offspring of diabetic women, and there is some evidence to suggest that higher lipid peroxidation levels and lower antioxidants may be a causative factor (88, 324).
Together with the characterization of several impairments in antioxidants in intrauterine tissues from diabetic animals throughout pregnancy, animal models of diabetes and pregnancy have been highly relevant in the understanding of the relevance of antioxidants that counteract these insults in the prevention of maternal diabetes-induced embryopathy. Both vitamin E and C have been shown to reduce diabetic embryopathy (324, 325, 358). These vitamins reduce lipoperoxidation and change the activity of SOD and catalase in different fetal organs (275). Lipoic acid reduces resorption and malformation rates and prevents damage in the placental vasculature in animals rendered diabetic through streptozotocin administration during pregnancy (5, 378). Likewise, pregnant diabetic rats fed the antioxidant butylated hydroxytoluene show decreased occurrence of malformations in offspring (90). Administration of vitamins E and C reduces fetal dysmorphogenesis and TBARS concentrations in fetal livers from diabetic rats, although the doses needed to normalize development in offspring of diabetic animals are above physiological levels, probably due to the difficulties to reach the embryonic tissues to exert their protective functions (52). Indeed, transgenic mice that overexpress SOD are protected from chemical diabetes-induced embryo malformations (126). Folate treatments are capable of reducing malformation rate in chemical models of diabetes and pregnancy, and are also able to increase the expression of antioxidant enzymes in the embryonic yolk sacs, and to reduce the concentrations of apoptotic proteins in the embryos from diabetic animals (109, 391). Antioxidant treatments also reduce lipoperoxidation and correct the altered ratio between vasodilator and vasoconstrictor prostanoids in the diabetic placenta (374).
Thus, supplementation with some antioxidants could be beneficial in the treatment of diabetes. Certainly, in type 1 diabetes, vitamins E and C have been shown to improve oxidative stress in children and improves fetal outcome in experimental diabetic pregnancy (88, 149). However, in GDM, blood glucose levels rise following prenatal vitamins (332), which has been linked to the development of the components of metabolic syndrome in adulthood for the male offspring (339). The DAPIT study has shown no benefits of vitamins E and C supplementation in preventing preeclampsia in type I diabetic pregnant women, although the work suggests that a subgroup of diabetic women with lower antioxidant plasma concentrations could be a group that may be benefited with these treatments (240). A very recently published study has found that women who had taken probiotics in pregnancy had a reduced frequency of GDM (226). Clearly, more studies are required to fully understand the short- and long-term health benefit of these dietary supplements.
Therefore, despite the potential benefit of treatments capable of reducing oxidative stress in GDM (77, 159, 258), there are a few clinical studies evaluating its potential beneficial effects in GDM. Thus, whether or not increased antioxidant intake can reduce the complications of GDM in both mother and fetus needs to be explored. However, studies examining the efficacy of antioxidants in the treatment of diabetic complications such as micro- and macro-angiopathy have been disappointing (334). After intensive investigations for decades, the current consensus is that the evidence obtained so far is not strong enough to recommend supplementation with antioxidants in the prevention or amelioration of diabetic complications, and tight glucose control remains the best, most efficacious measure to prevent diabetes induced damage.
Collectively, the above data show clear beneficial effects of antioxidants on diabetes and pregnancy animal models, indicating the relevance of oxidative stress in diabetes-induced pregnancy complications. There are difficulties in studying these issues in human diabetic pregnancies, and no safe and efficacious dose of antioxidants has been found so far to be helpful in GDM. Future understanding of human maternal-to-fetal transport of antioxidants may help addressing this important issue.
B. Flavonoids as potential antioxidant supplements to reduce oxidative stress in GDM
There is some evidence to suggest that dietary phytochemicals, including flavonoids, may be of potential benefit in the management of GDM. Epidemiological studies have suggested beneficial health effects of the chronic consumption of flavonoid-rich foods (8, 247). These health effects have been mostly attributed to their antioxidant properties (343).
Resveratrol is a polyphenol phytochemical that has been identified in almost 70 species of plants, including red grapes, peanuts, vegetables, berries, beverages, and herbal medicines (36, 85, 309). It is known to have a wide range of biological activities, including antioxidant, antiviral, anti-inflammatory, cardioprotective, and chemopreventive properties (318). Resveratrol modulates the expression of a series of intracellular signaling proteins, cellular proliferation, inflammation, and apoptosis in different cell lines (318).
Resveratrol contains two aromatic groups that enable it to function as an antioxidant by forming stable radicals via resonance structures, thereby preventing continued oxidation (326). In vitro studies show that resveratrol protects LDLs against peroxynitrite-mediated oxidation (94). Resveratrol has been shown to protect against oxidative stress-induced endothelial dysfunction in type 2 diabetes (396). It inhibits vascular NADH/NADPH oxidase, thus decreasing both basal cellular superoxide generation and NO removal. It also increases eNOS and iNOS, which are likely to play a role in resveratrol-mediated cardioprotection (reviewed in Refs. 29, 318). Although there are no data on the beneficial effect of flavonoids in GDM, studies from our own laboratory have demonstrated that resveratrol has potent anti-inflammatory (203) and antioxidant (Lappas, personal communications) properties in human placenta. Further, in these studies, we showed that resveratrol was able to alleviate inflammation via activation of sirtuin (SIRT)-1.
C. PPARs as targets to reduce oxidative and nitrative stress in GDM
PPARs (isoforms α, δ, and γ) are ligand-activated transcription factors that heterodimerise with retinoid X receptor (RXR). It has been shown that agonists of PPAR possess antidiabetogenic, anti-inflammatory, and antioxidant effects (244). Indeed, adipose tissue from obese GDM women has shown reduced PPARγ expression compared to uncomplicated pregnant obese patients (47). Likewise, our previous studies have shown lower placental PPARγ, PPARα, and RXRα expression in women with GDM (142, 156). In keeping with this, there is increasing evidence for a role of the PPAR system in regulating the metabolic and pathways involved in the pathophysiology of GDM (376). PPAR ligands, including the natural ligand 15d-PGJ2 and the synthetic anti-diabetic drug troglitazone, are potent anti-inflammatory and/or anti-oxidative agents that repress the expression of a number of inflammatory genes, oxidative stress, and regulating factors, including the transcription factor NF-κB in human placenta (205, 209). The prostaglandin 15d-PGJ2 is an endogenous agonist of the nuclear receptor PPARγ with relevant anti-inflammatory properties, capable of reducing NO production in the placenta from healthy patients (156). Placentas from GDM patients show reduced 15d-PGJ2, diminished PPARγ concentrations, and impaired capacity to downregulate NO production in the presence of 15d-PGJ2, possibly contributing to the observed increases in placental NO production (156). PPARγ agonists have also been shown to be a negative regulator of NO production in the diabetic rat placenta, also capable of regulating the diabetes-induced dysbalance in MMPs and their endogenous inhibitors named tissue inhibitors of metalloproteinases (40, 289).
Sulfonylurea agents, including glimepiride and glibenclamide, exhibit PPARγ activity (103). A randomized, controlled trial showed that both the insulin- and glyburide-treated women were able to achieve satisfactory glucose control and had similar perinatal outcome (199). Although the effect on oxidative stress was not determined, previous studies have demonstrated the effectiveness of these drugs in reducing diabetes-induced oxidative stress (272).
Diets enriched in n-3 polyunsaturated fatty acids (PUFAs) prevent the high plasma levels of lipoperoxides observed in the offspring from diabetic rats (385). PUFAs are activators of PPARδ, and recent studies have shown that PPARδ agonists are able to reduce lipoperoxidation in the placenta from diabetic rats (195). Interestingly, a recent study demonstrated that diets enriched in PPAR ligands prevent diabetes-induced embryonic NO overproduction, congenital malformations, and resorptions (140).
Overall, these data suggest that overproduction of reactive oxygen and nitrogen species can be regulated by PPAR ligands in different gestational tissues in experimental models of diabetes, as well as in the GDM placenta.
X. Future Directions
The involvement of oxidative stress in GDM is well documented and has been discussed above. However, there are very few studies available on the impact of oxidative stress on placental development and function and thus fetal development and subsequent adult diseases.
Cell adhesion molecules are important mediators of cellular contacts, proliferation, differentiation, and invasion. Dysregulation of these processes is associated with many complication of pregnancy, including GDM. There is, however, a paucity of data on the relationship between oxidative stress and adhesion molecules in placenta. Our preliminary data demonstrate that HX/XO regulates the expression of adhesion molecules (selectins, cadherins, and catenins) in human placenta (Lappas, personal communications). Clearly, more studies are required to determine the effect of ROS, RNS, and antioxidants on these molecules.
To compensate for the increasing needs of the embryo, there is an increase in the capacity of the mitochondria in the placenta. Mitochondrial dysfunction causes cell damage and death by compromising ATP production and calcium homoeostasis and increasing oxidative stress. Certainly, in pre-eclampsia, the mitochondria are an important source of oxidative stress and lipid peroxidation (258). The role and function of mitochondria in GDM placentas and their role in the generation of ROS also needs to be further evaluated.
Nutrient transport from the mother to the fetus occurs via the placenta; however, there is a paucity of data on the impact of oxidative stress on these processes. Certainly, our preliminary data suggest that ROS may play an important role in the regulation of maternal to fetal transport (Lappas, personal communications). Using placental BeWo cells as a model, we have shown that HX/XO increases transcellular fatty acid transport using transwell system. Specifically, there is an increase in glucose transport from the maternal circulation (upper chamber) to the fetal circulation (lower chamber). This would favor fetal fat accretion as well as stimulating oxidative stress in the fetus. However, further studies are required to fully elucidate the effect of ROS, RNS, and antioxidants on nutrient transport in the placenta.
Animal models have been particularly useful in delineating the effects of oxidative stress on the fetus. Although antioxidant therapies have been useful in the treatment of these fetal abnormalities, there is a paucity of data for their effectiveness in GDM women and their offspring. Thus, whether or not increased antioxidant intake can reduce the short- and long-term complications of GDM in both mother and fetus needs to be explored. Of potential benefit may be flavonoids as these are found naturally in many plants. Diets enriched in unsaturated fatty acids capable of activating PPARs to regulate both oxidative and nitrative stress may also be of benefit in GDM.
The appropriate development of the placenta is crucial to normal fetal programming. It is feasible that oxidative stress may disrupt normal intrauterine programming, thus leading to metabolic diseases later in life. Thus, future studies addressing the role of oxidative stress in GDM-induced intrauterine malprogramming are encouraged.
XI. Conclusions
Pregnancy is a state of oxidative stress as a consequence of high metabolic activity in the feto-placental compartment. During normal pregnancies, oxidants have many physiological functions, promoting and controlling cellular fate and playing a crucial role in normal development through cellular signaling. In absence of a parallel increase in antioxidative activity, oxidative stress will result. The elevated ROS levels in a normal pregnancy are exceeded in GDM. The level of oxidative stress might change the course and the severity of the side effects of the disease. Overproduction of ROS can lead to massive cellular damage by acting on proteins, lipids, and DNA. In the case of a systemic oxidative stress such as in maternal diabetes, it can also entail biochemical disturbances of the fetus (77, 259, 307). It is possible, that the management of oxidative stress, along with tight glycemic control, could be beneficial both preconceptionally and during pregnancy in women at risk of GDM. However, whether or not antioxidant supplementation or eating a diet rich in antioxidants can improve the consequences of oxidative stress in the offspring is yet to be elucidated. Clearly, more studies are required to fully understand the short- and long-term health benefit of reducing oxidative stress during diabetic pregnancies.
Footnotes
Acknowledgments
Dr. Martha Lappas is a recipient of a National Health and Medical Research Council (NHMRC) RD Wright Fellowship (grant no. 454777). The work from M. Lappas's laboratory was funded by project grants from NHMRC (grant no. 454310), Diabetes Australia Research Trust (DART), Melbourne Research Grant Scheme (MRGS), ANZ Charitable Trust (Medical Research and Technology Grant), and the Medical Research Foundation for Women and Babies. Work from A. Jawerbaum's laboratory was supported by ANPCYT (PICT 2006 00084 and PICT 2005 32268).