
Abstract
The gastrointestinal epithelium transports solutes and water between lumen and blood and at the same time forms a barrier between these compartments. This highly selective and regulated barrier permits ions, water, and nutrients to be absorbed, but normally restricts the passage of harmful molecules, bacteria, viruses and other pathogens. During inflammation, the intestinal barrier can be disrupted, indicated by a decrease in transcellular electrical resistance and an increase in paracellular permeability for tracers of different size. Such inflammatory processes are accompanied by increased oxidative stress, which in turn can impair the epithelial barrier. In this review, we discuss the role of inflammatory oxidative stress on barrier function with special attention on the epithelial tight junctions. Diseases discussed causing barrier changes include the inflammatory bowel diseases Crohn's disease, ulcerative colitis, and microscopic colitis, the autoimmune disorder celiac disease, and gastrointestinal infections. In addition, the main cytokines responsible for these effects and their role during oxidative stress and intestinal inflammation will be discussed, as well as therapeutic approaches and their mode of action. Antioxid. Redox Signal. 15, 1255–1270.
Introduction
The expression of the proteins forming the epithelial TJ barrier of the intestine depends in a segment-specific manner on the function of the respective intestinal segment as shown by several studies on rat or mouse intestine (40, 56, 69, 90) (Fig. 1). Among the claudins that tighten the paracellular barrier of the gastrointestinal tract are claudin-1, -3, -4, -5, and -8. These claudins have their highest expression in the colon; and it follows that the colon has the highest epithelial resistance. The expression of pore-forming TJ proteins such as claudin-2, -7, and -12 are at a maximum in jejunum and ileum; and these segments have the lowest epithelial resistances. Taking only the small intestine into account, the duodenum exhibits the strongest expression of the majority of the “tightening” TJ proteins and the highest resistance. Occludin (for review see Blasig et al. (13), in this Forum), the exact barrier function of which is hitherto not clearly assigned, is expressed in the same pattern as the “tightening” TJ proteins, with a maximum abundance in the colon and a minimum in jejunum (69).

Studies on human intestine confirm the expression of claudins-1, -2, -3, -4, -5, -7, and -8 in the colon, expression of claudins-1, -2, -3, and -4 in the duodenum, and expression of claudins-2 and -4 in the jejunum (19, 33, 115, 125, 133). However, there are not many studies on the distribution and abundance of TJ proteins in man, especially along the small intestine.
Intestinal Inflammation and Other Diseases Causing Leak Flux Diarrhea
Inflammatory processes affect the barriers of the respective organ [for reviews, see Frey and Antonetti (39), Coisne and Engelhardt (29), both in this Forum] and in the intestine they are accompanied by disruption of the epithelial barrier. The resulting passive loss of solutes into the intestinal lumen and the subsequent osmotically driven water flow results in “leak-flux diarrhea”, one of the main consequences of intestinal inflammation. The increased uptake of antigens and macromolecules from the intestinal lumen mediated through this epithelial barrier dysfunction can further exacerbate the inflammatory process, ending up in a vicious circle. In this manner, barrier dysfunction is a perpetuating principle during gastrointestinal inflammation. Since epithelial TJs are important in the maintenance of barrier function, regulatory changes in their function that are commonly found during intestinal inflammation can have severe consequences. Other transmembrane proteins relevant for intestinal inflammation are junctional adhesion molecules [JAMs; reviewed by Bazzoni (11), in this Forum]. Moreover, epithelial cell apoptosis, epithelial gross lesions like erosions or ulcers as well as epithelial endocytosis with transcytotic antigen uptake do contribute to inflammatory barrier dysfunction.
Crohn's disease
Crohn's disease (CD) is one of the two main inflammatory bowel diseases (IBD) that leads to segmental inflammation of gastrointestinal segments, especially along the distal small and/or large intestine. Barrier dysfunction as indicated by the decrease in epithelial resistance has been shown to be present in inflamed intestinal segments (132). The resultant loss of barrier function results in ion passage and leak-flux diarrhea, which is one of the main symptoms of CD. Factors further contributing to the barrier dysfunction in CD are a strong upregulation of epithelial apoptosis and transcellular antigen uptake most likely via endocytosis (105, 111).
A number of morphological findings contribute to the loss of barrier function. These include a diminished number of horizontally oriented TJ strands, a reduced depth of the TJ mesh, an increased number of TJ strand breaks >25 nm, and the appearance of particle-type TJ strands. Each of these appears to contribute to the increased permeability of mannitol across the intestinal barrier in CD (100, 133). The downregulation of “tightening” TJ proteins in combination with an upregulation of pore-forming TJ proteins enhances TJ permeability, which in turn promotes leak-flux diarrhea. For occludin and the “tightening” claudins-3, -5, and -8, a reduced expression in CD has been reported, although claudin-3 may play only a minor role, as it becomes intracellularly localized and not within the TJ. Furthermore, an upregulation of the expression of the pore-forming claudin-2 was observed (89, 133). Additionally and perhaps at least as important, the “tightening” claudins-5 and -8 were redistributed off the TJ into the basolateral membrane and the cytoplasm, further increasing the permeability of the TJs (133) (Fig. 2).
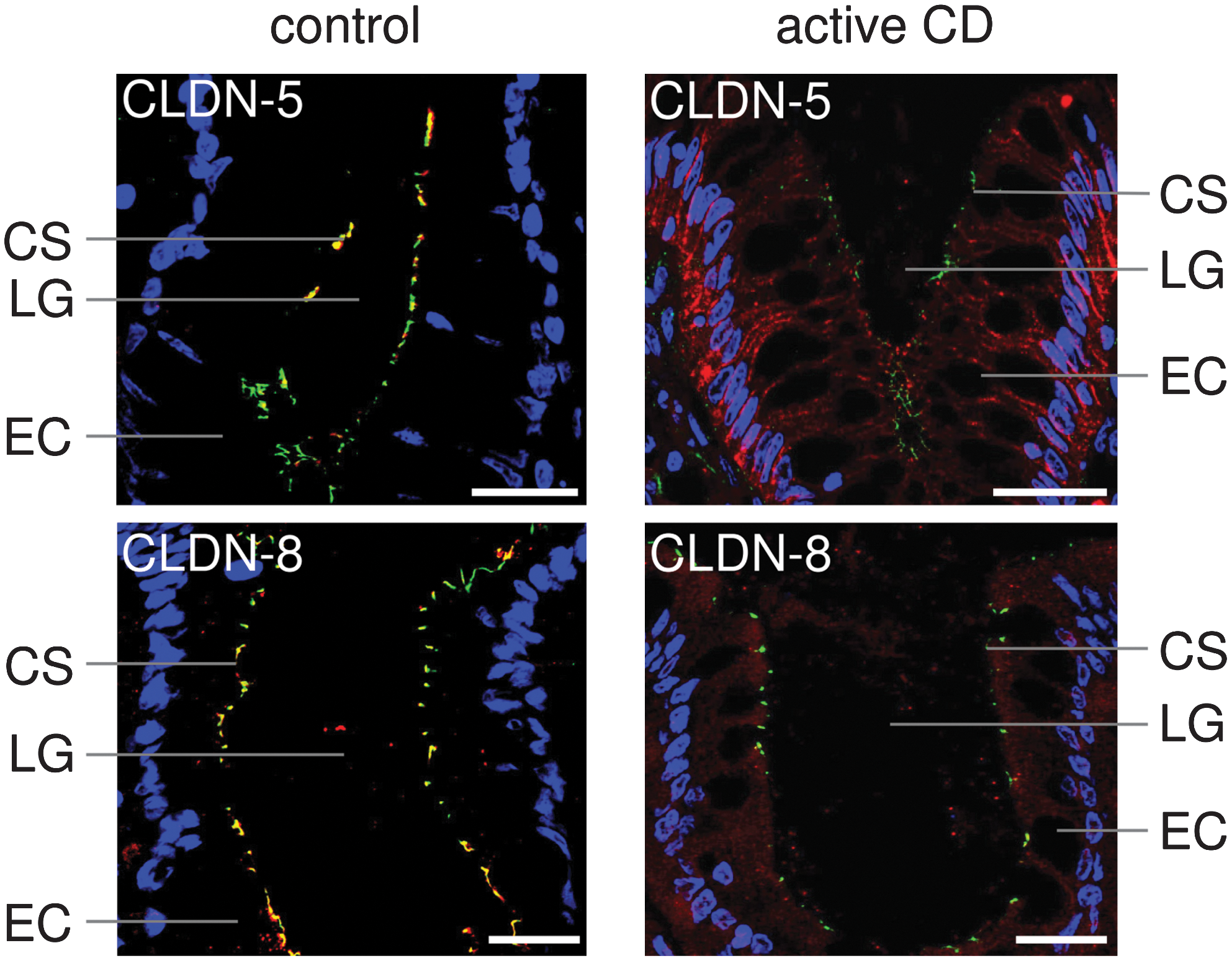
Besides alterations of the intestinal paracellular barrier for ions and small solutes, an increased permeability for macromolecules was reported which resulted from enhanced transcytotic transport. There is convincing evidence in the literature that the uptake of ovalbumin and horseradish peroxidase, as model antigens into IBD enterocytes, and their subsequent vesicular transport to the paracellular space contributes to the barrier defect in CD (105, 111).
Ulcerative colitis
Ulcerative colitis (UC) constitutes another main inflammatory bowel disease (IBD), but, in contrast to CD, is predominately restricted to the large intestine and, also in contrast to CD, is characterized by epithelial lesions. However, UC is comparable to CD in that the barrier dysfunction leads to an increased loss of solutes and water into the intestinal lumen resulting in leak-flux diarrhea, which is one of the predominant symptoms in UC.
The factors responsible for barrier dysfunction in UC are similar to those in CD, including an increase in epithelial antigen transcytosis and a change in TJ structure with a reduction in TJ strand count and in the depth of the TJ main meshwork; although, in contrast to CD, strand breaks are not as frequent as in UC (100, 105). Again, the upregulation of the pore-forming TJ protein claudin-2, together with a downregulation of occludin and several “tightening” TJ proteins like claudin-1 and -4 contribute to the barrier defect observed in UC (54) (Fig. 3). Upregulated epithelial apoptosis also plays an important role for the increase in intestinal permeability for molecules of up to 4 kDa (54, 112), which, together with an arrest in epithelial restitution, leads to the appearance of local gross leaks like apoptotic foci, (micro)erosions and even ulcer-type lesions (43, 44). Whereas local leaks can already be observed in mild colonic inflammation, microerosions and ulcer-type lesions are only found in moderately-to-severely inflamed tissues (44).

Microscopic colitis
Microscopic colitis (MC), comprising lymphocytic colitis (LC) and collagenous colitis (CC), ranks among the least severe inflammatory bowel diseases, and are typically accompanied by watery diarrhea. In contrast to CD and UC, the mucosa appears normal in MC, and epithelial lesions, such as ulcers and erosions, are usually lacking. In LC, the intraepithelial lymphocytes are increased to more than 20 per 100 epithelial cells, whereas CC is additionally characterized by subepithelial collagenous bands that can thicken the basal lamina up to almost 9-fold (126). The ability to absorb ions like sodium and chloride decreases in this situation due to these collagenous bands, as they represent a significant subepithelial barrier (19).
Similar to CD and UC, the permeability for tracer molecules of the mucosa is increased in patients with CC (74). Again, increased transcellular permeability may partly account for this loss of barrier function. However, hitherto there are no reports on TJ structure or freeze fracture electron microscopy data on TJ strand morphology.
Alterations in the expression of TJ proteins may resemble those observed in CD and UC, even if they are not as pronounced. The decrease of occludin and of the “tightening” claudin-4 is paralleled by an increase of the channel-forming claudin-2 in most of the patients, while claudin-1, -3, and -5 remain unchanged (19). Epithelial apoptotic rate is not increased in CC (19, 103), which is in sharp contrast to UC and CD, where the increase of the apoptotic rate is one of the main factors contributing to the increase in epithelial permeability, and this may in part explain the absence of gross epithelial lesions.
Celiac disease
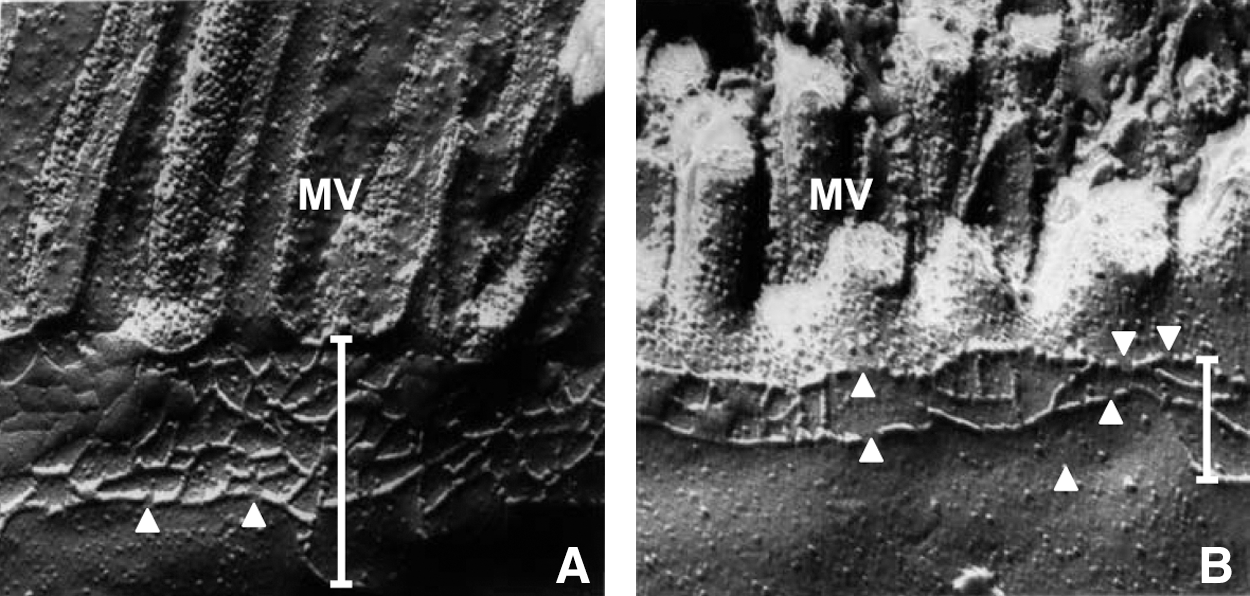
Celiac disease, which is characterized by gluten sensitivity, is an autoimmune disorder of the small intestine in individuals with the human leukocyte antigen haplotype DQ2. One of the most prominent clinical features is diarrhea. Functional alterations of the small intestine include a decline in barrier function, as indicated by an increase in permeability for tracer molecules of different sizes and a decrease in epithelial resistance (99, 104, 122). As far as epithelial TJs are concerned, a lot of structural and molecular changes have already been reported. Jejunal biopsies revealed a reduction of horizontal TJ strands and an increase of TJ strand discontinuities in freeze fracture electron microscopy (102) (Fig. 4).The exposure of the human epithelial colorectal adenocarcinoma cell line Caco-2 to gliadin, a prolamin that, together with glutenin, forms gluten, leads to a reorganization of actin filaments and a reduction of the expression of the TJ proteins occludin, claudin-3, -4, the TJ-associated protein ZO-1, and the adherens junction (AJ) protein E-cadherin (99). In duodenal biopsies from celiac disease patients, the level of tyrosine-phosphorylated ZO-1 and subsequently the level of ZO-1 bound to occludin were decreased (25). There are two additional studies confirming reduced ZO-1 levels in biopsies from celiac disease patients (73, 84).
Gastrointestinal infections
Gastrointestinal infectious diseases are characterized by a variety of symptoms including abdominal cramps, nausea, and diarrhea. In most of the cases, these symptoms are caused by an inflammation of the intestinal mucosa. Pathogenic bacteria, viruses, and protozoa often disrupt the intestinal barrier during gastrointestinal infections. Several pathogens have developed mechanisms to disrupt the TJs of intestinal epithelial cells which include the movement of TJ proteins off the TJ strands, for example, into the cytoplasm via alteration of the actinomyosin complex (8, 78), the dephosphorylation or degradation of TJ proteins (110, 127), and the disruption of the binding of TJ proteins like claudin-1, occludin, and ZO-1 to each other (75). Certain TJ proteins can function as receptors for bacterial toxins or viruses, which facilitates their entrance into the cells (28, 58).
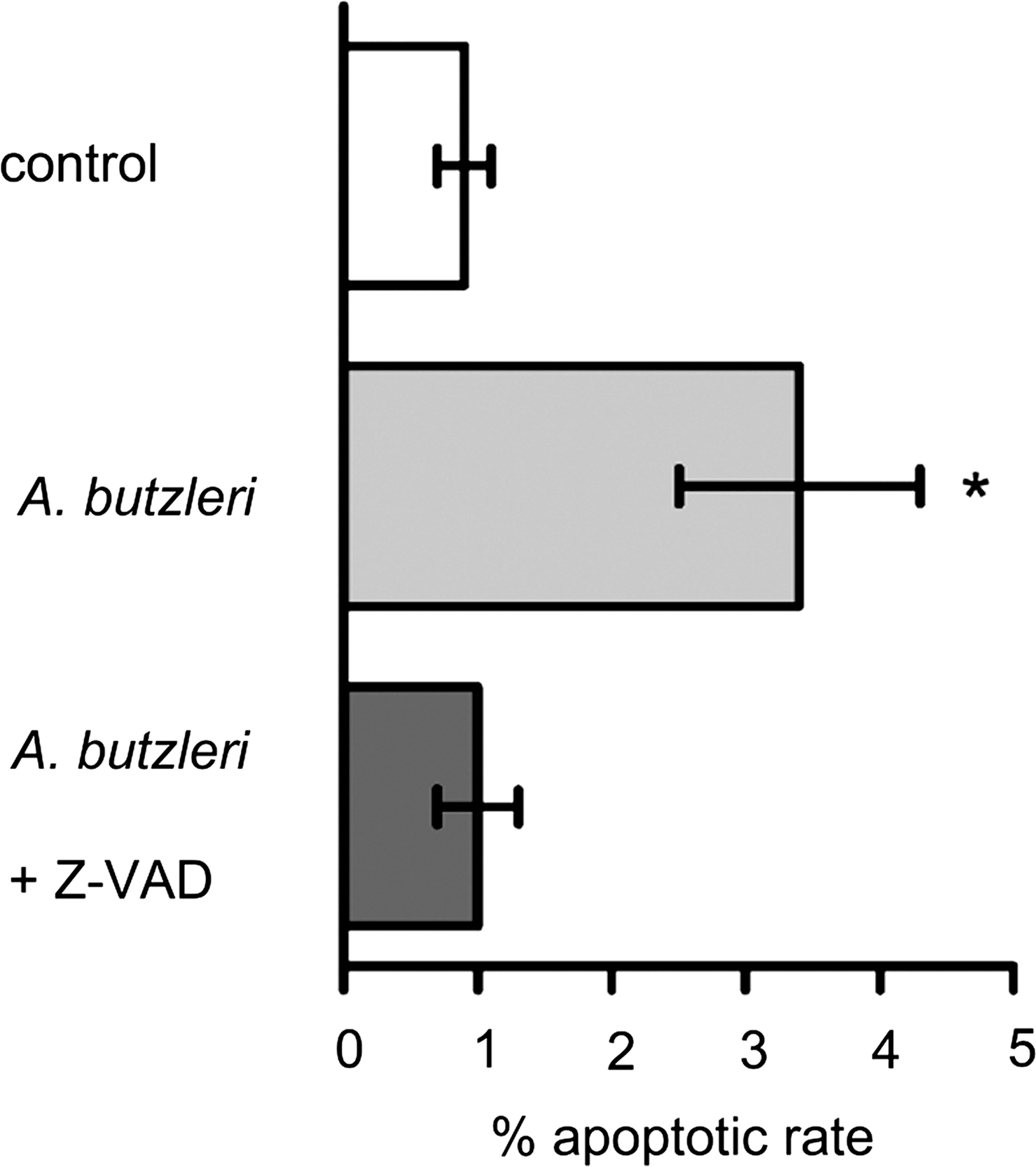
As an example, the epithelial barrier dysfunction caused by Escherichia coli can be due to the induction of focal leaks. For this purpose, bacteria use their α-hemolysin to facilitate their paracellular passage through focal leaks (119). In the cell line HT-29/B6, which is a subclone from the human colon adenocarcinoma cell line HT-29, Arcobacter butzleri causes similar TJ changes as observed in an inflamed epithelium in CD. The “tightening” TJ proteins claudin-1, -5, and -8 are downregulated, and claudin-1 and -8 are additionally redistributed off the TJs, whereas claudin-2, -3, and -4 remain unchanged. Further to these changes, apoptosis is increased (18) (Fig. 5). A downregulation of claudin-1 and an increase in epithelial apoptosis can also be observed in gastrointestinal infections with the protozoan Giardia lamblia (117).
Infections associated with the human immunodeficiency virus (HIV) cause an increase in epithelial apoptosis and an altered expression of several TJ proteins, leading to increased permeability and decreased epithelial resistance. The pore-forming claudin-2 is upregulated, whereas the “tightening” claudin-1 is downregulated. No changes in occludin and claudin-4 could be observed (32). A slightly different pattern of TJ protein changes was found in patients with norovirus infection. While claudin-1, -2, and -3 remained unaffected, occludin, claudin-4, and -5 were downregulated (118).
Oxidative Stress in Intestinal Inflammation
Functionality of the intestinal epithelium is continuously challenged by the preservation of tissue homeostasis and cellular integrity when faced with oxidants. These can either be endogenously generated by activation within epithelial cells or may be exogenous oxidants which can be in part the result of an impaired barrier function. Under physiological conditions, the release of oxidants, and on the other hand the production of antioxidants, is balanced; whereas under oxidative stress oxidants are in excess to antioxidants. Besides oxidant producing cells like neutrophils, several oxidants such as nitric oxide, hydrogen peroxide (H2O2), and the superoxide anion are found to be elevated within the epithelium during intestinal inflammation (60, 79). Through several mechanisms, including their mutual activation, these oxidants can also impair intestinal barrier function contributing to the typical disease pattern of intestinal inflammatory diseases. Intestinal defense mechanisms range from the maintenance of high antioxidant concentrations to the upregulation of antioxidant enzymes and the induction of apoptosis.
Reactive oxygen species
Reactive oxygen species (ROS) are molecules that interfere with several signaling pathways, thereby modulating the expression of different genes and the activity of various kinases and phosphatases (for review, see Ref. 52). ROS represent a group of reactive molecules that contain oxygen such as the superoxide anion, H2O2, and the hydroxyl radical. Superoxide originates from the electron transport chain, or is produced by enzymes such as NADPH oxidase and xanthine oxidase, and is converted to H2O2 in a reaction that is catalyzed by the superoxide dismutase (SOD). H2O2 can, by undergoing the Fenton reaction, react to the hydroxyl radical which is much more reactive than superoxide and H2O2. Under normal conditions, however, H2O2 underlies effects from antioxidant systems like catalase or glutathione peroxidases and becomes neutralized to water and oxygen (62). Further, sources of ROS in the intestinal lumen include xenobiotics, toxins, several bacteria, bile acids, desquamated mucosal cells, and activated neutrophils (9).
In inflammatory processes of the intestine, ROS are produced as part of the host response to kill bacteria or damage viruses. In IBD, however, ROS are produced in excessive levels and their deleterious effects are assumed to contribute to barrier dysfunction as indicated by a disruption of the composition of the TJs which accompany the functional changes.
It was shown that H2O2 induces the tyrosine phosphorylation of several TJ proteins such as ZO-1 and occludin and of adherence junction (AJ) proteins such as E-cadherin and β-catenin in Caco-2 cells which led to a disassembly of TJs and AJs with a subsequent disruption of the paracellular barrier formed by TJs and AJs (59, 91). In another study, claudin-4 was found to be translocated off the TJs after H2O2 treatment, which was due to activation of the p38 mitogen-activated protein (MAP) kinase (81). ROS can also affect the actin cytoskeleton, leading to a further disruption of the intestinal barrier (10, 60). However, the observed oxidation and nitration of the actin cytoskeleton is proposed to be mediated through nitric oxide (NO) (10). H2O2 has been shown to induce the inducible isoform of the enzyme nitric oxide synthase (iNOS) which in turn produces NO, leading to a disruption of the intestinal barrier (10). This induction of iNOS probably involves the transcription factor nuclear factor κB (NF-κB), as it is activated by H2O2 and involved in the expression of iNOS (12, 101). The activation of NF-κB by H2O2 also increases the production of proinflammatory cytokines such as tumor necrosis factor-α (TNF-α) in the mucosa. Conversely, TNF-α induces the production of ROS, thereby creating a vicious circle exacerbating the inflammatory process. Superoxide can react with NO, forming the highly noxious peroxynitrite which has been shown to promote epithelial barrier dysfunction (37).
ROS are also able to oxidize membrane phospholipids, leading to the formation of lipid radicals that can react with molecular oxygen to form peroxyl radicals. These oxidizing species can oxidize further lipids in a chain reaction called lipid peroxidation (reviewed in Ref. 34). In this manner, phospholipids that make up the plasma membranes of the epithelial cells are finally destroyed. Lipid peroxidation is elevated in both, UC and CD. However, the ROS responsible for the damage seem to differ. Lipid peroxidation in CD is likely to be induced by the superoxide anion and the hydroxyl radical, whereas H2O2 and hypochlorous acid seem to be involved in the lipid peroxidation in UC (61).
Nitric oxide
NO is a free radical messenger molecule, which derives from the amino acid L-arginine in a redox reaction catalyzed by the enzyme NOS. In the intestinal epithelium, NOS is available in two isoforms, the constitutive NOS which produces low levels of NO and the inducible NOS (iNOS) which produces high levels of NO. Under normal conditions, only the constitutive NOS is expressed, whereas iNOS can be induced after exposure to cytokines, microbes, and other activating stimuli (77). An important regulator in the induction of iNOS expression is the transcription factor NF-κB which has been found to be activated in CC, UC, CD, and in gastrointestinal infectious diseases (6, 12). The concentration of iNOS is elevated during these inflammatory processes. This elevation subsequently leads to an increase of NO in the colonic epithelium, with NO levels correlating to disease severity (60, 79) (Fig. 6). In several gastrointestinal infections, iNOS and the subsequent production of NO is induced by the pathogens or their lipopolysaccharides (LPS) (12).
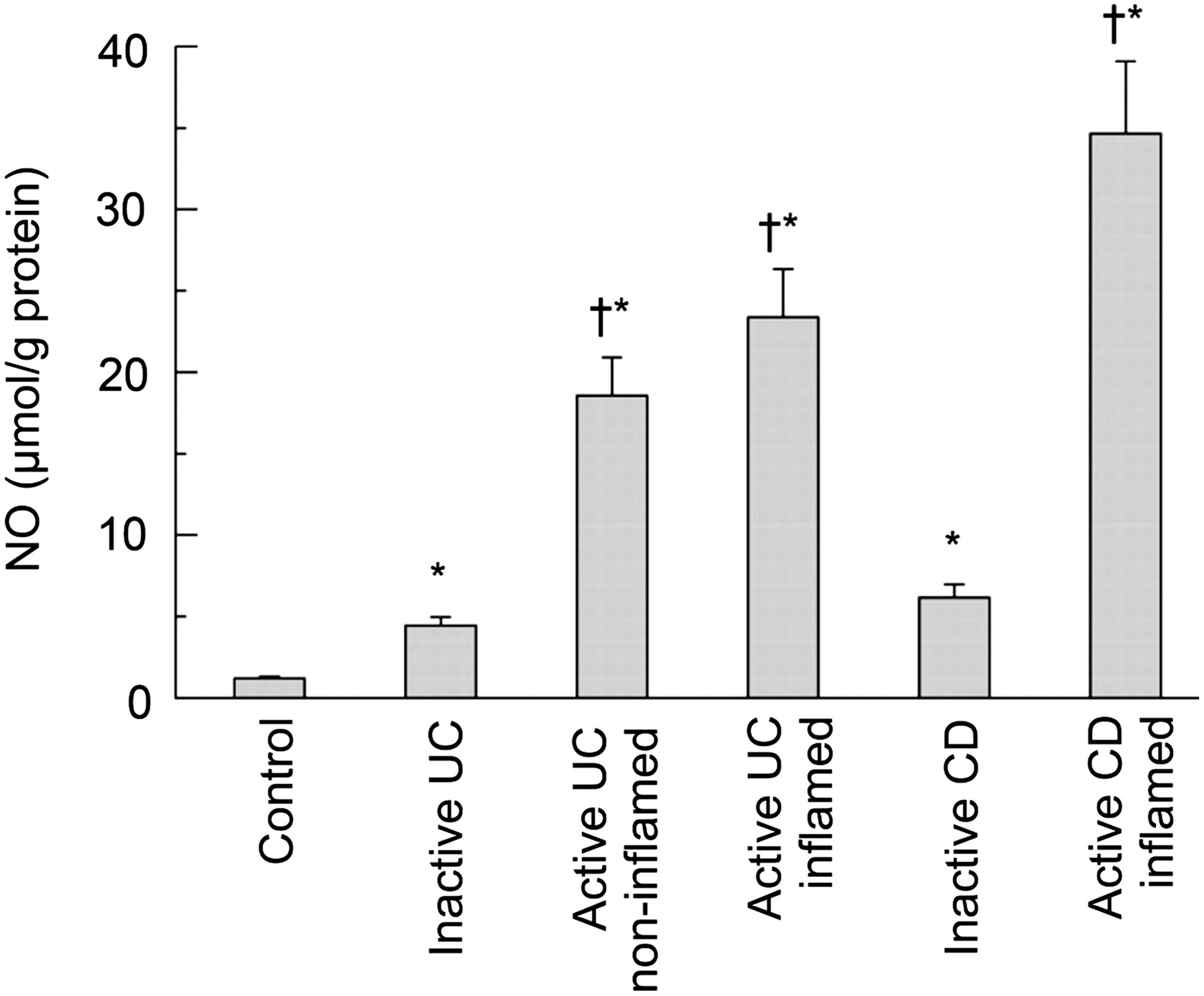
In the inflamed mucosa, NO has differential effects on intestinal barrier function depending on the respective concentration. Small concentrations of NO seem to have protective effects and to maintain mucosal integrity, whereas large concentrations have pro-inflammatory and damaging effects. In inactive as well as in active IBD, the NO levels are elevated when compared to healthy mucosa. The fact that NO levels are also slightly elevated in non-inflamed mucosal segments supports the assumption of NO-mediated damage to the epithelial barrier only in high concentrations (60).
So far, it is unknown which mechanisms underlie the cellular damage that is induced by NO under oxidative stress. However, it has been proposed that NO reacts with oxygen radicals like superoxide to peroxynitrite, which in turn mediates the toxic effects observed with NO, such as ATP depletion, de-arrangements of the cytoskeleton, and hyperpermeability (37). A subsequent reaction of peroxynitrite with tyrosine leads to nitrotyrosine, the level of which increases with NO levels in the intestinal mucosa and thus does the activity of the disease (60).
Several effects of NO on the intestinal barrier have been observed, such as the nitration of tubulin and actin in active CD and UC, which leads to a disruption of the actin cytoskeleton and a subsequent epithelial barrier dysfunction (60). It can be assumed that this barrier dysfunction is at least partly due to a disruption of TJs as the TJ proteins are tightly connected with the actin cytoskeleton via the ZO-proteins (reviewed in Ref. 46). In a study with Caco-2 monolayers, NO decreased epithelial barrier function via a decline in the expression of ZO-1, ZO-3, occludin, and claudin-1. Furthermore, a mislocalization of ZO-1 was observed in this model (51).
Neutrophils
Neutrophils transmigrate the inflamed mucosa in UC and CD after being attracted by chemotactic agents including cytokines such as interleukin-8 or soluble bacterial products. Despite their ability to destroy pathogens, they can also cause extensive tissue damage that can be observed in IBD. In UC, the activation of neutrophils even leads to the formation of crypt abscesses which are typical for this disease (76).
In a study with the human colon adenocarcinoma cell line T84, it was shown that the transmigration of neutrophils induces epithelial apoptosis (63), and this may contribute to tissue damage and the subsequent barrier dysfunction caused by neutrophils in intestinal inflammation. Aside from apoptosis, there are several ways by which neutrophils induce oxidative stress in the intestine and subsequently result in tissue damage. Once neutrophils are activated in order to destroy luminal pathogens, activation of the cell membrane located NADPH oxidase transfers electrons from cellular NADPH to oxygen to produce superoxide radicals (9). The impact of superoxide on intestinal barrier was already discussed in the section about reactive oxygen species.
Myeloperoxidase (MPO), an enzyme found in neutrophils, partly accounts for the oxidative stress by neutrophils (for review, see Ref. 76). In combination with H2O2, MPO can oxidize chloride to hypochlorous acid, which in turn can react with tyrosine to 3-chlorotyrosine. This molecule is thought to directly damage tissue and cause barrier dysfunction in inflammatory processes.
Another reaction catalyzed by MPO is the oxidation of nitrite to nitrogen dioxide, a reactive nitrogen species that can react with tyrosine to nitrotyrosine. In a third MPO-dependent pathway, nitrite is oxidized to nitryl chloride, a further reactive nitrogen species that can be metabolized to nitrotyrosine by the MPO-generated hypochlorous acid. In UC and CD, nitrotyrosine levels are elevated, as already mentioned above in this review. The formation of nitrotyrosine via the neutrophilic MPO is independent of peroxynitrite and thus of iNOS activity. The increase of nitrotyrosine in UC has been shown to be rather associated with MPO than with iNOS activity (61), indicating a pivotal role of neutrophils in the inflammatory process in UC. Neutrophils also contain iNOS (60), which catalyzes the reaction that produces NO, as already described earlier in this review. Besides the production of toxic agents and the induction of cell apoptosis, neutrophils stimulate the production of cytokines, some of which act as chemoattractants for neutrophils.
Antioxidant systems in the intestine
Glutathione (GSH) rates among the most important cellular antioxidants. In the intestine it is found in twice as high concentrations as in the liver (reviewed in Ref. 72). When reducing oxidants, GSH itself is oxidized to glutathione disulfide (GSSG), which can be regenerated by the NADPH-dependent GSH reductase. The maintenance of the GSH levels can also be achieved through synthesis from cysteine or the uptake of exogenous GSH. The preservation of the redox balance of GSH/GSSG in the intestine is of great importance, as it has been shown that an imbalance, triggered by the presence of higher amounts of GSSG, is linked to intestinal inflammation and disease activity in UC and CD (97). However, it remains unknown so far whether the decreased levels of GSH are due to an increased GSH oxidation or the result of a decreased biosynthesis and uptake of GSH.
Another important antioxidant enzyme is SOD, which catalyzes the dismutation of superoxide to oxygen and H2O2. H2O2 gets further converted to water and oxygen by catalase or glutathione peroxidases, so that excess reactive oxygen species (ROS) are eliminated. Under inflammatory conditions, the expression of Cu/Zn-SOD, one of the isoforms of the SOD found in the cytosol of the intestinal epithelium, is decreased. In contrast, Mn-SOD, a mitochondrial isoform in the intestinal epithelium, has been found to be increased, but the elevation remained limited to the enzymatically inactive form (62). This decrease of active SODs, together with an increase of ROS, leads to an imbalance between oxidants and antioxidants during intestinal inflammation, which is typical for oxidative stress.
Metallothioneins are small proteins that are able to reduce reactive oxygen and nitrogen species. During inflammation, a rapid upregulation of metallothioneins is induced by NO, glucocorticoids, and proinflammatory cytokines such as interleukin-1 (IL-1), TNF-α, and interferon-γ (IFN-γ) (reviewed in 124).
Cytokines
In intestinal inflammatory processes, cytokines such as TNF-α, IL-13, and IFN-γ play an important role in the regulation of the epithelial barrier function (54, 67, 133). On the one hand, the levels of these cytokines are elevated in CD, UC, MC, and gastrointestinal infectious diseases (12, 54, 66, 116) and the cytokine profiles are specific for the different disease processes. On the other hand, the respective alterations (e.g., in TJ structure and function) can be ascribed to these changes in cytokine profiles. In CD, for example, TNF-α and IFN-γ are important regulators, whereas IL-13 and TNF-α dominate the responses in UC. Furthermore, similar patterns of elevated cytokines lead to almost identical changes in epithelial TJ structure.
Interferon-γ and tumor necrosis factor-α
The pro-inflammatory cytokines IFN-γ and TNF-α were found to be elevated in UC, CD, and also in MC (54, 66, 116), where they promote several changes that contribute to the observed barrier dysfunction. IFN-γ and TNF-α decrease transepithelial resistance, increase paracellular permeability for Lucifer yellow, and increase bacterial translocation, which in turn exacerbates the inflammation of the intestinal mucosa (26, 27). IFN-γ and TNF-α induced a redistribution of occludin, JAM-A, claudin-1, and -4 off the TJs in T84 cells. This redistribution seems to depend on endocytosis of TJ proteins (16, 17). IFN-γ can also decrease the expression of ZO-1 and perturb the actin cytoskeleton in T84, leading to a disassembly of the TJ structure (130). In HT-29/B6 cells, the expression of occludin was decreased after treatment with IFN-γ, TNF-α, or both cytokines together, and these cytokines are thought to alter the TJ strand formation and thus to influence the barrier function of the epithelium. The effects on transepithelial resistance and the expression of occludin are even synergistically enhanced when both cytokines were applied together (68), which could at least in part be due to an IFN-γ-dependent induction of TNF receptor expression (1).
The increase in epithelial permeability for fluorescein sulfonic acid after IFN-γ treatment observed in Caco-2 cells is at least partly due to an increased expression of iNOS and thus due to an elevated production of NO, as the inhibition of iNOS after IFN-γ treatment diminishes this IFN-γ dependent permeability increase (24, 120). The treatment with a combination of IFN-γ, TNF-α, and IL-1β, referred to as cytomix, even exceeded the IFN-γ induced expression of iNOS, whereas the treatment with TNF-α or IL-1β alone had no effect on iNOS expression (24).
Interestingly, the rate of epithelial apoptosis was not altered after treatment with IFN-γ, whereas TNF-α treatment increased apoptosis (54, 103). TNF-α induces ROS, which inactivates MAP kinase phosphatases which usually suppress the activation of c-Jun N-terminal kinases (JNKs). Thus, the subsequent activation of JNKs could contribute to TNF-α-induced epithelial apoptosis (reviewed in Ref. 45). Furthermore, TNF-α intensified focal leak induction and thus facilitated bacterial translocation across HT-29/B6 cell monolayers (119). In addition, TNF-α, but not IFN-γ, led to an upregulation of the TJ protein claudin-2 in HT-29/B6 cells, indicating an essential role of TNF-α in those intestinal inflammatory processes, where the expression of claudin-2 is significantly elevated (67, 133).
Interleukin-13
IL-13 is increased in patients with UC, with IL-13 levels being much higher in UC than in CD, where it seems to be the key effector cytokine (54). It has been shown to decrease the resistance of the intestinal epithelium through an increase of the pore-forming TJ protein claudin-2 and an increase in epithelial apoptotic events in HT-29/B6 cells (54, 103, 133). IL-13 also impairs epithelial restitution, which underlines the important role of this cytokine in UC, since (micro)erosions and ulcer-type lesions can be observed much earlier and much more pronounced in UC than in CD. In this context, IL-13 has been shown to increase bacterial translocation across HT-29/B6 cell monolayers by intensifying focal leak induction (119).
Interleukin-8
IL-8 is elevated in many gastrointestinal infectious diseases (12) and is also increased in UC, where its level correlates with the level of the MPO, the neutrophil-derived enzyme that catalyzes several reactions that produce ROS (7). IL-8 is a chemoattractant and an activator for neutrophils, which in turn produce several cytokines and ROS. Thus, the increased production of IL-8 in several inflammatory processes of the intestine is linked to the production of ROS. Furthermore, because IL-8 rates among the cytokines produced by activated neutrophils, the induction of IL-8 results in a vicious circle of further neutrophil recruitment.
Transforming growth factor-β
TGF-β, an immunosuppressive cytokine, generally enhances the intestinal epithelial barrier properties (57, 86). It has been shown to increase the transepithelial resistance as well as the expression of the “tightening” TJ protein claudin-1 and is known to stabilize the epithelial barrier function after IFN-γ treatment or exposure to harmful microorganisms (57, 86). Howe and co-workers showed that TGF-β can prevent the decrease of claudin-2, occludin, and ZO-1, as well as the change in ZO-1 distribution caused by enterohemorrhagic E. coli O157:H7. However, the responsible mechanism remains unknown, as no direct upregulatory effects of TGF-β could be observed on claudin-2 or occludin (57). A further mechanism, by which epithelial barrier function is enhanced, is the TGF-β-dependent increase in the restitution velocity after injury, as shown in T84 monolayers (70). Interestingly, TGF-β also induces epithelial–mesenchymal transition (EMT), which plays an important role in the conversion of tumors from benign to invasive and metastatic. During EMT, genes are repressed that encode claudins, occludin, and E-cadherin and subsequently the epithelial barrier function is decreased (for review, see Ref. 42).
Therapeutic Approaches
A wide range of therapeutic options have been successfully applied in the treatment of intestinal inflammation and IBD, including compounds naturally occurring in the human organism, plant components as well as classical drugs. However, all of these therapeutic approaches are only able to induce or to maintain remission in IBD but not to cure the disease. The target molecules by which amelioration is mediated are very diverse, and in some cases the underlying mechanisms remain largely unknown. Reduction of oxidative stress, induction of mucosal healing, and prevention of TJ structure changes rate among the beneficial effects of many therapeutics used in the treatment of various inflammatory intestinal disorders including UC, CD, MC, celiac disease, and gastrointestinal infectious diseases. In the following section, therapeutic approaches are presented focusing on their antioxidative aspects.
Zinc
Zinc, an essential trace element, has antioxidant functions, both direct and indirect (reviewed in Ref. 88). It is an inhibitor of the NADPH oxidase which catalyzes the production of superoxide. Furthermore, it is a cofactor of the Cu/Zn-SOD that catalyzes the dismutation of superoxide to H2O2, which under physiological conditions is decomposed (e.g., by catalase) to water and oxygen. Zinc is also involved in the regulation of metallothioneins, a protein family known to reduce the amount of NO and ROS. Zinc supplementation in healthy individuals was shown to decrease lipid peroxidation and to inhibit the induction of the expression of proinflammatory cytokines, such as, for example, TNF-α and IL-1β which generate ROS. In this zinc-induced inhibition of TNF-α and IL-1β expression, NF-κB is inhibited by the zinc-finger protein A20 (88). Taking all these findings into account, zinc deficiency in intestinal inflammation does not only result in increased oxidative stress and subsequent damage but even in the maintenance of chronic inflammation through pro-inflammatory cytokines.
Zinc deprivation in Caco-2 cells decreased the transepithelial electrical resistance and altered the molecular composition of the TJs and AJs in several ways. Occludin, ZO-1, β-catenin, and E-cadherin were shown to be delocalized, and F-actin and β-tubulin were disorganized. Moreover, dephosphorylation of occludin and hyperphosphorylation of β-catenin and ZO-1, as well as a decrease in ZO-1, occludin, and β-tubulin were observed. These disruptions of TJs and AJs facilitated the migration of neutrophils through the paracellular space (36).
Zinc supplementation as a therapy has been shown to improve intestinal barrier function in patients with CD in remission. The inflammatorily elevated intestinal permeability was significantly reduced by a daily oral uptake of zinc (113). However, this improvement could also be simply due to the recovery of zinc deficiency which is known to occur in IBD patients, as well as in patients with a Helicobater pylori infection (55, 107). Nevertheless, improved barrier function of the intestinal epithelium may contribute to a reduced risk of relapse in CD (113). In the rat small intestinal epithelial cell line IEC-6, cell restitution was enhanced after treatment with zinc, indicating an improved epithelial repair (20). Studies on the effect of zinc supplementation on TJ and AJ proteins in humans are lacking. However, a study in piglets showed that the decrease in intestinal lactulose permeability was accompanied by an upregulated expression of occludin and ZO-1 after zinc supplementation (134). Hence, zinc is a promising therapeutic approach, even if the direct mechanisms behind this therapy remain in part unknown.
Budenoside
Budenoside is a glucocorticoid which, due to a pronounced first pass effect in the liver, has predominant topical effects when applied to the intestine. Therapeutically, it is used in the treatment of most IBDs, including UC, CD, and CC, where it can result in clinical and histological improvement in a significant portion of the patients (14, 48, 65). In response to budesonide, the inflammatory infiltrate in the intestinal mucosa was found to be decreased in patients with CC. Furthermore, it was proposed that the improvement is for the most part based on a reduction of the iNOS mRNA content (14, 15). Hence, the amount of iNOS mRNA seems to be correlated to the grade of inflammation and the symptoms in patients suffering from CC. The decrease of iNOS mRNA accompanied by a reduced production of NO after budenoside treatment was also observed in biopsies from patients with UC. The underlying mechanism, however, remains unknown, albeit there are indications for an involvement of soluble mediators that are produced by the mononuclear cell system that includes neutrophils (65). In CD, budenoside treatment induced and prolonged remission, but the remission usually did not exceed one year (48, 49). For CC, it was shown that after stopping the budesonide treatment there is a high risk of relapse (14).
Nitric oxide
Besides the damaging effects of NO in high concentrations, NO seems to be advantageous in small amounts, since it was shown to have a protective effect on barrier function in Caco-2 cell monolayers during oxidative stress induced by H2O2 (Fig. 7). However, the exact mechanism of this protection remained unknown (59). NO can act as a scavenger of free radicals, thereby terminating chain reactions such as lipid peroxidation. As an inhibitor of the Fenton reaction, it can prevent the generation of the highly reactive hydroxyl radicals (21). Furthermore, NO can reduce tyrosine phosphorylation of cellular proteins such as TJ and AJ proteins, which has been shown to be increased by oxidative stress (59, 91). Another likely mechanism of NO to mediate protection of epithelial barrier function is through interference with intracellular signaling, since NO has been shown to interact with signaling proteins (for review, see Ref. 47).

5-Aminosalicylic acid
5-Aminosalicylic acid (5-ASA), also known as mesalazine, is a drug with anti-inflammatory and antioxidant activity, which is used in the therapy of IBD. The decomposition of superoxide rates among the antioxidant properties of 5-ASA, as well as the chelation of iron, which leads to an inhibition of the Fenton reaction and subsequently to a decrease in the production of the highly reactive hydroxyl radical. 5-ASA is also able to scavenge peroxyl radicals, which are formed during lipid peroxidation, and thereby inhibits the chain reaction that destroys membrane lipids. Furthermore, it can act as an alternative substrate for MPO, thereby inhibiting the reactions which are catalyzed by MPO and thus reducing oxidative stress (for review, see Ref. 128).
5-ASA was shown to maintain remission in UC over a test period that lasted 48 weeks (114). In a dextran sodium sulfate (DSS)-induced colitis model of ulcerative colitis in the rat, inflammation and permeability were reduced by mesalazine. Furthermore, changes in the DSS-induced irregular distribution of occludin were prevented (53). Since 5-ASA is known to inhibit the NF-κB and several kinase pathways as well as the response to IL-8, it may be hypothesized that these mechanisms are involved in the mesalazine-induced amelioration of the intestinal barrier. However, the exact mechanism(s) are hitherto unknown.
Wip1
Wip1 is a serine/threonine phosphatase that selectively inactivates the p38 MAP kinase, an important signal transducer in cellular responses to external stress signals. The expression is induced by the transcription factor p53 (38). In Caco-2 cells where p53 is mutated, the treatment with H2O2 results in a decrease of the transepithelial resistance and an increase of the permeability for fluorescein isothiocyanate labeled dextran (FITC-dextran). These changes were accompanied by a translocation of claudin-4 off the TJs. Exogenous Wip1 was able to prevent the H2O2-induced claudin-4 dislocalization and to reverse the decrease in transepithelial resistance, as well as the increase in epithelial permeability (81). Since the p38 MAP kinase is activated in IBD (123), Wip1 could be a new therapeutic option to treat IBD.
TNF-α antibodies
Therapeutics based on TNF-α antibodies like Remicade® (infliximab) or Humira® (adalimumab) seem to be very promising. In addition, TNF-α antibody treatment in patients with CD improved the epithelial barrier function measured as epithelial resistance in impedance spectroscopy. This improvement was based on a normalization of the inflammatorily elevated apoptotic rate, which reached the normal range in almost all patients 2 weeks after a single infusion of infliximab (132). Another study showed mucosal healing after 4 weeks of a single infliximab infusion. It is reasonable to assume that the decrease in epithelial apoptosis induced by infliximab is the first step towards mucosal repair (31). In the aforementioned study by Zeissig and coworkers, the TJ proteins occludin, claudin-1, and -4 were also investigated, but no changes after TNF-α antibody treatment could be observed within this time frame (132). However, since TNF-α induced a rapid upregulation of claudin-2 in HT-29/B6 cells (67, 133), an inhibition of this increase by TNF-α antibodies is assumable, but this was not investigated.
Two studies with 364 patients suffering from UC showed that treatment with infliximab induced mucosal healing and remission in significantly more patients than in a control group receiving placebo treatment (98). Hitherto, there are no studies showing whether the decrease of epithelial apoptosis is also involved in the effect of the infliximab treatment in UC.
Berberine
Berberine is an isoquinoline alkaloid which has been used in Chinese medicine for many centuries. Antidiarrheal, anti-inflammatory and antimicrobial activities are proposed.
Berberine reduced intestinal inflammation in rats with trinitrobenzene sulfonic acid (TNBS)-induced colitis. Furthermore, the production of IL-8, a chemoattractant for neutrophils, was inhibited (135). In a study with Caco-2 cells, berberine inhibited the changes induced by the proinflammatory cytokines TNF-α and IFN-γ. The decrease in transepithelial resistance was reversed and the redistribution of occludin off the epithelial TJ was inhibited. Furthermore, the cytokine-induced increase in permeability for FITC-dextran was prevented (64). However, the underlying mechanisms remain to be elucidated. So far, it is known that berberine can inhibit the protein kinase A as well as the NF-κB signaling pathway (83, 136), both of which are important regulators of inflammation and TJ assembly, respectively.
In vitro studies revealed additional antioxidant properties of berberine. It was able to scavenge free radicals such as NO and superoxide, to chelate iron, and to terminate lipid peroxidation (109). Taking these effects of berberine into account, it seems to be a promising therapeutic agent for multimodal treatment concepts of intestinal inflammation in the future.
Quercetin
The flavonoid quercetin that can be found in different fruits like apples and onions was shown to enhance barrier function in Caco-2 cell monolayers. Here, it upregulated the expression of the TJ protein claudin-4 that is subsequently inserted into the TJ strands. Hence, as claudin-4 rates among the “tightening” TJ proteins, transepithelial resistance was increased and the permeability of Na+ was decreased. The expression of occludin, claudin-1, -3, and -7 remained unchanged (3) (Fig. 8). The mechanisms underlying the quercetin induced upregulation of claudin-4 could not be completely resolved, although protein kinase C was inhibited and it was speculated that quercetin may directly interact with the DNA or could affect transcription factors (80, 131).

Quercetin, like all flavonoids, is able to scavenge free radicals like superoxide and the hydroxyl radical as well as to terminate lipid peroxidation by scavenging the lipid peroxyl radical (93). Despite these antioxidant and pronounced barrier-preserving functions of quercetin, hitherto there are no studies on its role in IBD.
Butyrate
Butyrate is a short-chain fatty acid (SCFA) produced in significant amounts within the intestinal lumen through bacterial fermentation of dietary carbohydrates. It is an important source of fuel for colonocytes, and a diminished oxidation of butyrate, meaning an energy deficiency, has been associated with UC (22). In a study with healthy humans, it was shown that butyrate increases the amount of the antioxidant GSH, the role of which has already been described in detail. This effect was based on an increase of the heavy catalytic subunit of the glutamate-cysteine ligase (GCLC), a rate-limiting enzyme involved in the production of GSH (50). Additionally, a study with rats after induction of TNBS-colitis also showed that supplementation with dietary fiber resulted in elevated GSH concentrations. The ameliorating changes found in this study were ascribed to butyrate and propionate, which were shown to be the mainly produced SCFAs after fiber administration. Further changes observed in this study were a decrease in neutrophil infiltration and the amelioration of mucosal damage (94). In another study, the maintenance of remission in UC by administration of the same dietary fibers was as effective as mesalazine over a 12-month test period (35).
The pro-inflammatory cytokine IL-8 was inhibited by butyrate in HT-29 cells, which could account for the decrease in neutrophil infiltration, since IL-8 functions as a chemoattractant for neutrophils. Butyrate also inhibited iNOS activity leading to a reduced production of NO. These inhibitory effects seem to be due to an inhibition of the transcription factor NF-κB which plays an important role in the regulation of iNOS expression and the release of several cytokines. In this context, butyrate has also been shown to decrease the production of TNF-α in intestinal biopsies (106). The inhibition of NF-κB is probably due to the ability of butyrate to suppress the histone deacetylase (85). In a study with HT-29 cells and human colonocytes, butyrate reduced H2O2-induced DNA damage (96). It seems that butyrate can decrease the damage caused by oxidative stress by the aforementioned increase in antioxidant systems and by affecting DNA repair systems.
Probiotics
Probiotics are microorganisms which promote healthy intestinal function. In several studies, an ameliorating effect of probiotics on barrier function has been shown (71, 108, 129).
In a study examining the effect of Lactobacillus rhamnosus GG (LGG), probiotic bacteria occurring in the human intestine, on H2O2-treated Caco-2 cells, LGG as well as LGG supernatant and soluble proteins produced by LGG were able to decrease paracellular permeability for FITC-inulin and to attenuate the disruption of the intestinal barrier. LGG did not change the level of H2O2, excluding a direct antioxidant effect. H2O2 induced a re-distribution off the intercellular junctions of the TJ protein occludin, the TJ-associated protein ZO-1, and the AJ proteins E-cadherin and β-catenin. This redistribution was reduced by the LGG-produced proteins p40 and p75 (108). An earlier study had previously shown that LGG prevented apoptosis induced by the cytokines TNF-α, IFN-γ, and IL-1α in HT-29 cells. In this former study, LGG activated the anti-apoptotic Akt/protein kinase B and inhibited the activation of the pro-apoptotic p38 MAP kinase (129). However, the mechanisms leading to an amelioration in H2O2-treated cells did not involve Akt, whereas isoforms of the protein kinase C and the MAP kinases ERK1 and 2 were found to be activated (108).
TJ proteins are also affected by probiotics as shown in mice with DSS-induced colitis. The decreased expression and redistribution of the TJ proteins occludin, claudin-1, -3, -4, and -5 was prevented by a therapy with VSL#3, a mixture of eight probiotic bacterial strains. It was further shown that the DSS-induced increase in epithelial apoptosis and permeability for the marker Evans blue was inhibited by VSL#3 (71). The underlying mechanisms were not elucidated in this study. In other studies with the same probiotic mixture, as well as with the nonpathogenic E. coli strain Nissle 1917, remission could be retained in patients with UC (92, 121). An improvement of the disrupted epithelial barrier function in T84 cells infected with enteropathogenic E. coli could be obtained by the addition of Saccharomyces boulardii, including changes in transepithelial resistance, permeability for 3H inulin, ZO-1 distribution, and apoptosis. The translocation of EPEC into the cells was diminished in an ERK 1/2-dependent manner (30). In CD, S. boulardii was able to reduce disease activity, but again the underlying mechanisms remained elusive (87).
Conclusions and Future Perspectives
During intestinal inflammation, the impairment of the epithelial barrier is a widespread feature found in various diseases including UC, CD, MC, and gastrointestinal infectious diseases. Barrier dysfunction is mainly caused by changes in the TJs, but other features like induction of epithelial apoptosis, gross lesions and leaks, as well as epithelial restitution arrest have been found to contribute. Oxidative stress is strongly correlated with several of these features, thus being a main contributor to the barrier dysfunction in intestinal inflammation. Several cytokines are elevated during intestinal inflammation, and the resulting cytokine profiles are specific for the respective diseases and are connected to the changes in TJ protein expression. Furthermore, oxidants and cytokines promote each others production, leading to an exacerbation of inflammatory processes in the intestine. In this manner, inflammation and barrier dysfunction trigger each other in a vicious circle (discussed as leaky gut concept). Similar influences have been assumed in regard to the post-infectious irritable bowel syndrome. Above this, it has even been discussed that systemic inflammation derived from barrier disturbances could contribute to cardiovascular risk. In order to overcome the disruption of the intestinal barrier, therapeutic agents use a variety of target molecules and function through different mechanisms to exert their beneficial effects, including mucosal healing, prevention of the TJ structure, and reduction of oxidative stress.
During the last decade the interest in TJ research has grown immensely, as well as the knowledge about the importance of TJs for the maintenance of the intestinal homeostasis, which was proven by various experimental findings. For example, the TJ protein claudin-2 was found to act as a water channel, which is functionally of great relevance. Nevertheless, a lack of knowledge in various fields of TJ research still remains. Further experimental exploration will be needed to elucidate the functional meaning of a variety of individual TJ proteins, as well as the mechanisms as to how permeability and barrier properties are performed by single TJ proteins. Moreover, the signaling properties of TJ proteins within cell contacts remain a central field of investigation, which seem to be of great importance for gastrointestinal diseases and tumorigenesis. A further interesting and important field of future research concerns the therapy of gastrointestinal diseases. Several therapeutic approaches use the TJs as targets. However, the underlying pathways remain elusive and whether they can support each other in multimodal therapy approaches.