
Abstract
Introduction
Innovation
This study provides new insights into understanding of the innate immune and inflammatory response in the airway epithelia. In normal homeostasis with bacteria, hydrogen peroxide (H2O2) is continually released to the airway surface liquid to eliminate bacteria, whereas dual oxidase 2 (DUOX2)-mediated H2O2 generation regulates host defense by enhancing production of pro-inflammatory mediators (production of IL-8 and MUC5AC) and stimulates adaptive immune responses when the mucosa barrier is irritated by infection. Furthermore, the results from this study may prompt the development of pharmacological inhibitors of DUOX2 as a potential therapeutic approach to control inflammation.
Substantial evidence has indicated that transient reactive oxygen species (ROS) can be produced by receptor-mediated biochemical processes, although ROS such as superoxide anion and hydrogen peroxide (H2O2) have been thought to be by-products of aerobic respiration that could be damaging to DNA, proteins, and lipids. ROS generation for host defense has been extensively studied in terms of NADPH oxidase (NOX) in phagocytic cells. However, after identification of the homologs of gp91phox (NOX1, NOX3-5, and dual oxidase 1-2 [DUOX1-2]) from nonphagocytic cells, the function of the generated ROS has been extended into an understanding of various cellular events, including cell growth, differentiation, apoptosis, and inflammation responses (5, 26). NOX isozymes contain transmembrane domains harboring four heme-binding histidines and conserved NADPH- and FAD-binding sites in a long COOH-terminal region (26). DUOX isozyme contains an additional helix-loop-helix structural domain (EF)-hand that binds Ca2+ and extracellular peroxidase-like domain (25, 44). DUOX was originally identified as thyroid oxidase and important in thyroid hormone biosynthesis (5). However, recent studies have focused on a novel function of DUOX isozymes in host–microbe homeostasis for the mucosal immune response (5, 17). Human DUOX1 and DUOX2 play distinct roles in adaptive immunity of airway epithelial cells, although they have a quite similar structure. Th2 cytokines moderately increase Duox1 mRNA, whereas Th1 cytokines significantly increase Duox2 mRNA levels (13).
Recent studies have demonstrated a molecular link between TLR activation and NOX isozymes in innate immune response and host defense, suggesting that NOX activation is indispensable in TLR-mediated innate immune responses (5, 32, 35). We found that flagellin stimulates ROS generation through DUOX2 activation, which is dependent on Ca2+ mobilization in primary human nasal epithelial cells. Our results also indicate that flagellin-induced ROS generation through DUOX2 plays a role in TLR5-dependent MUC5AC and IL-8 expression in human airway epithelial cell and mouse mucosa. These results demonstrate that DUOX2-mediated ROS generation is essential for the inflammation process and, therefore, is a novel therapeutic target for infectious inflammatory diseases in the airway.
Results
Flagellin stimulates intracellular ROS production in normal human nasal epithelial cells
The molecular mechanism by which TLR-activated NOX leads to ROS production and triggers inflammation has not been clearly determined (7, 8, 24), although previous reports have reported that Nox is coupled with TLR in inflammation processes. To investigate the function of TLR-induced ROS production in the airway epithelial cells, which play an important role in host defense and inflammation, we stimulated primary normal human nasal epithelial (NHNE) cells with flagellin, a known TLR5 ligand, and then determined intracellular levels of ROS in terms of Nox acitivity by measuring fluorescence of 2′,7′-dichlorofluorescin diacetate (DCF-DA). ROS production increased rapidly within 5 min, reaching its peak at 30 min (3.7-fold over control), and then decreased after 30 min in NHNE cells in response to flagellin stimulation (Fig. 1A). Standard curve with exogenous H2O2 (1–1000 μM) in the DCF assay with NHNE cells were prepared and then the level of flagellin-induced H2O2 production was analyzed. It is found that the concentration of flagellin-induced H2O2 is around 150 μM. FliC, a structure subunit protein that polymerizes to form the filaments of bacterial flagella, is known to activate TLR5 (3, 38). To examine whether the FliC-TLR5 interaction is essential for ROS generation, we purified Salmonella typhimurium FliC and the N-terminal deleted mutant protein (FliCΔN; 31-118AA), which cannot interact with TLR5 (38). Stimulation of NHNE cells with WT FliC protein resulted in increased ROS production (4.2-fold over control), whereas mutant FliCΔN protein failed to induce ROS generation in NHNE cells (Fig. 1B). To confirm the involvement of TLR5 in this process, we explored the effect of neutralizing antibody against TLR5 (nAb/TLR5) on flagellin-induced ROS production in NHNE cells. Pretreatment of NHNE cells with nAb/TLR5 resulted in significantly decreased in ROS generation in response to flagellin (Fig. 1C). These results strongly indicate that interaction of TLR5 with flagellin is required for ROS generation in human airway epithelial cells.

DUOX2 is essential for flagellin-induced ROS generation in airway epithelial cells
We examined the expression level of NOX isozymes in NHNE cells to understand the ROS-generating system of flagellin-stimulated airway mucosa. Quantitative real-time polymerase chain reaction (RT-PCR) indicated that Duox1 and Duox2 were the predominant NOX isozymes in NHNE cells. Other NOX isozymes were minor or hardly detectable (Supplementary Fig. S1; Supplementary Data are available online at
To explore the roles of DUOX1 and DUOX2 in flagellin-induced H2O2 generation, we infected cells with lentiviruses expressing either DUOX1- or DUOX2-targeted shRNA. The infected cells exhibited a marked reduction by 70% in targeted mRNA (Fig. 1D, E). Next, we monitored H2O2 production using DCF-DA fluorescence. DUOX2-suppressed cells failed to generate H2O2 in response to flagellin, whereas scrambled shRNA virus-infected cells exhibited an increase in H2O2 (Fig. 1F). Interestingly, DUOX1 shRNA showed no inhibitory effect on H2O2 generation in response to flagellin stimulation. These results suggest that DUOX2 is responsible for the majority of flagellin-induced ROS generation in NHNE cells.
Flagellin-induced cytosolic [Ca2+] increase activates DUOX2
Human DUOX enzymes contain canonical calcium-binding EF-hands, suggesting that intracellular calcium ion plays a critical role in activation of DUOX. Furthermore, it has been reported that flagellin induces mobilization of intracellular calcium ion cells via ATP production in NCI-H292 cells (28, 29). To investigate flagellin-induced DUOX2 activation mechanism, we analyzed the intracellular calcium concentration ([Ca2+]i) and verified the effect of Ca2+ on H2O2 production. Stimulation of NHNE cells with flagellin resulted in increases in [Ca2+]i mobilization (Fig. 2A). Pretreatment of NHNE cells with 1,2-Bis(2-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid (BAPTA-AM), a Ca2+ scavenger, significantly reduced flagellin-induced ROS generation, compared with flagellin-treated control cells (Fig. 2B). This result indicates that flagellin-induced [Ca2+]i mobilization stimulates DUOX2, resulting in H2O2 generation.

Flagellin-TLR5 signaling axis stimulates ATP efflux, leading to activation of purinergic receptor (27, 28). The receptor activates Gαq protein-PLCβ cascade resulting in intracellular calcium mobilization. Suramin is an antagonist of purinergic receptor. We asked whether suramin can prevent H2O2 production in response to flagellin. Pretreatment of NHNE with suramin resulted in decreased flagellin-induced H2O2 production, suggesting that the activation of purinergic receptor is required for flagellin-dependent H2O2 production (Fig. 2C).
Based on the previous reports (7, 33) that a physical interaction between TLR and NOX isozyme plays a critical role in H2O2 generation, we performed a yeast two-hybrid assay using the toll/interleukin-1β receptor (TIR) domain of TLR5 as the bait and the COOH-terminal region of the human DUOX2 isozymes (DUOX2-C) as the prey. Yeast cells expressing pB42AD-TLR5 with pLexA-Duox2-C revealed a normal growth and produced blue colonies in the absence of leucine and in the presence of X-gal. The observation indicates that the TIR region of TLR5 interacted with the COOH terminal region of DUOX2 (Fig. 2D). We also performed an immunoprecipitation assay with HEK293 cells co-expressing TLR5 and DUOX2 to confirm the yeast two-hybrid results. The immunoprecipitation complex with antibody to DUOX2 showed an interaction with TLR5 in HEK293 cells (Fig. 2E). To evaluate the interaction of TLR5 with endogenous DUOX2, NHNE cell lysates were subjected to immunoprecipitation with antibodies to TLR5. Immunoblot analysis of the resulting precipitates with antibodies to DUOX2 or DUOX1 revealed that TLR5 interacts with endogenous DUOX2, not DUOX1 (Fig. 2F). However, the interaction was not increased in response to flagellin. These results suggest that DUOX2 localizes with the TLR5 signaling complex and efficiently activates DUOX2 followed by TLR5-induced calcium mobilization.
DUOX2 activation mediates MUC5AC and IL-8 expression induced by flagellin in human airway epithelial cells and the mouse nasal lavage fluid
The inflammatory response is initiated when airway epithelial cells respond to a bacterial or viral infection. Chemokines and inflammation-related molecules, such as IL-8 and MUC5AC, are involved in neutrophil recruitment to the inflamed region in the mucosal epithelium and they provide protection against pathogens (11, 41). We hypothesized that H2O2 generation is responsible for inflammatory reactions, such as MUC5AC and IL-8 expression, in response to flagellin stimulation. To elucidate the function of H2O2 in inflammation, we analyzed the effect of N-acetyl-
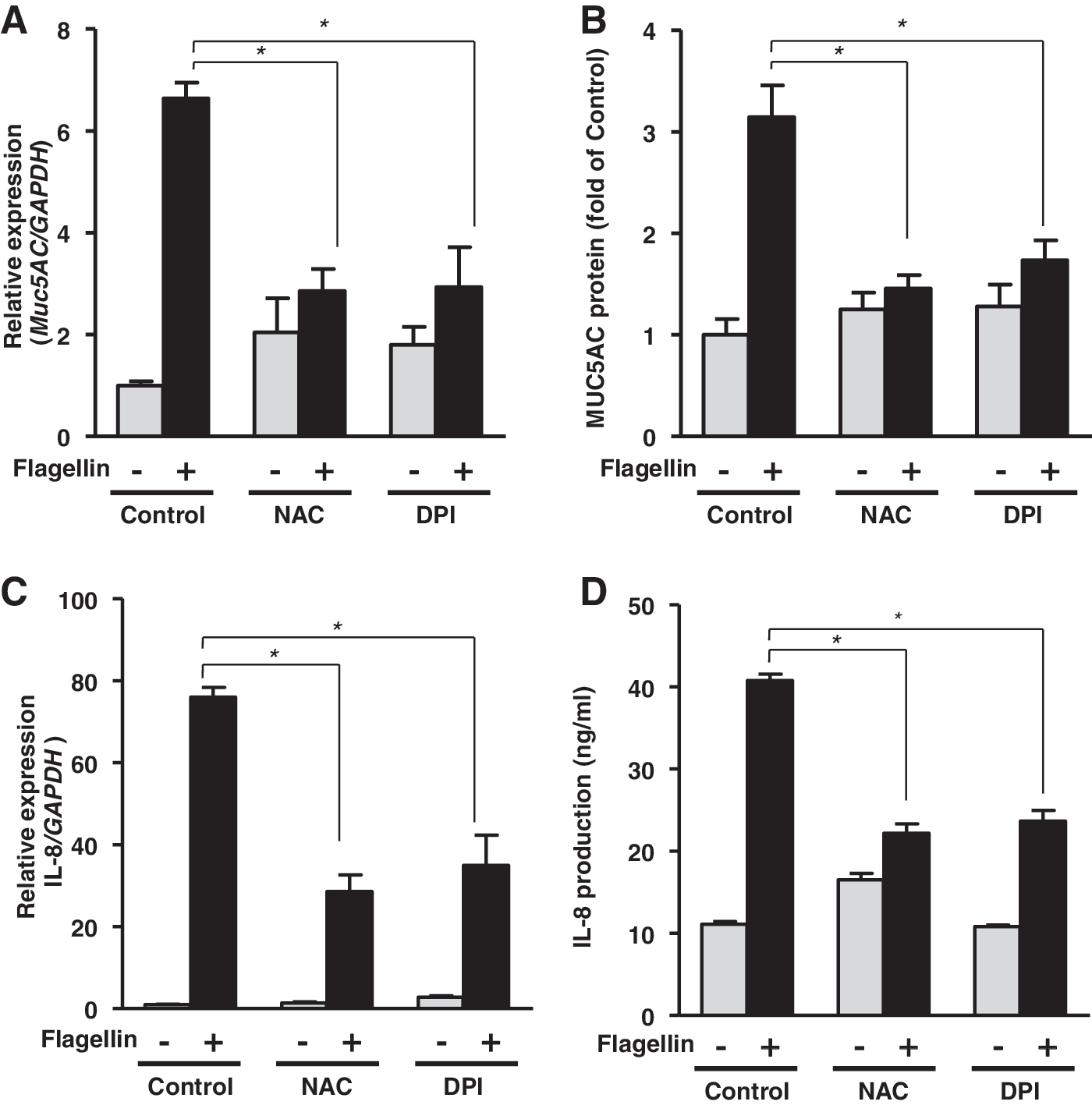
To identify the role of DUOX2 in the flagellin-induced inflammatory response in primary human airway epithelial cells, we explored the flagellin-induced inflammatory response in NHNE cells infected with a lentivirus encoding an shRNA specific to DUOX1 and DUOX2. Infection of NHNE cells with a lentivirus encoding a specific shRNA to DUOX1 and DUOX2 resulted in a significant reduction of Duox1 or Duox2, respectively (data not shown). Interestingly, the flagellin-mediated production of MUC5AC and IL-8 (Fig. 4A, B) was significantly reduced in DUOX2 shRNA-transfected cells compared with that in scrambled shRNA-transfected cells. However, knockdown of DUOX1 had no noticeable inhibitory effect on flagellin-induced MUC5AC or IL-8 production (Fig. 4A, B). Thus, these results strongly indicate that DUOX2 has an essential role in flagellin-induced H2O2 generation and inflammatory response in NHNE cells.
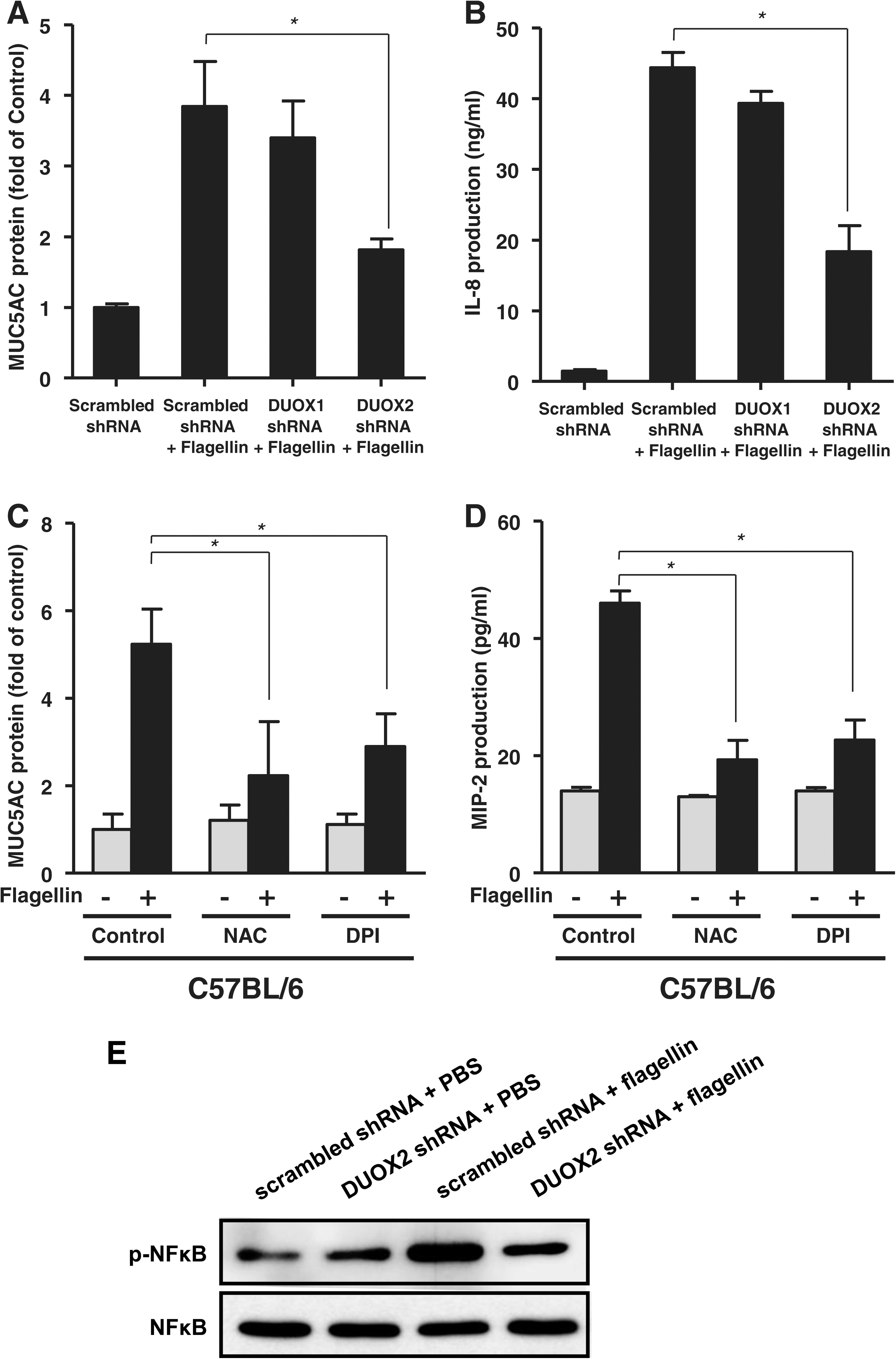
To determine the function of flagellin-induced H2O2 generation in the inflammatory response in vivo, we examined flagellin-induced MUC5AC and MIP-2α (IL-8 homolog in mouse) in nasal mucosa and nasal lavage fluid (NALF) of C57BL/6 mice in the presence of NAC or DPI. Mice pretreated with NAC or DPI had significantly inhibited the production of MUC5AC (Fig. 4C) and MIP-2α (Fig. 4D) in mouse NALF in response to flagellin compared to mice pretreated with flagellin alone. These results indicate that ROS generation by flagellin might be involved in the acute inflammatory response to bacterial infected in vivo. It has been reported that flagellin-TLR5 cascade activates nuclear factor kappa B (NF-κB) leading to immune responses (3). To explore the roles of DUOX2 in flagellin-induced NF-κB activation, NHNE cells were subjected into the knockdown of DUOX2 with lentivirus expressing DUOX2 shRNA. Scrambled shRNA virus-infected cells resulted in increased phosphorylation of p65 subunit, whereas DUOX2-suppressed cells showed a significantly decreased the phosphorylation, indicating that flagellin-mediated DUOX2 activation is essential for the activation of NF-κB (Fig. 4E). These results show that DUOX2 regulates NF-κB activity and production of chemoattractant such as IL-8 and MIP2α.
DUOX2 mutation downregulates the increased inflammation- and immune response- related genes expression induced by flagellin stimulation
We obtained Duox2 knockout mice from The Jackson Laboratory, which arose by spontaneous C to T transition in exon 16, causing a change from a highly conserved valine674 to glycine (21). Next, to evaluate the roles of DUOX2 in flagellin-induced inflammatory response in mouse nasal mucosa, we have performed transcriptional analysis using the Affymetrix GeneChip® Mouse Gene 1.0 ST Array made by Affimetrix, which comprised of more than 770,000 unique 25-mer oligonucleotide features constituting more than 28,000 gene and each gene is represented on the array by 27 probes. The data were processed using the robust multi-array analysis algorithm, which performs a background correction, a normalization step, and a probe-level summary.
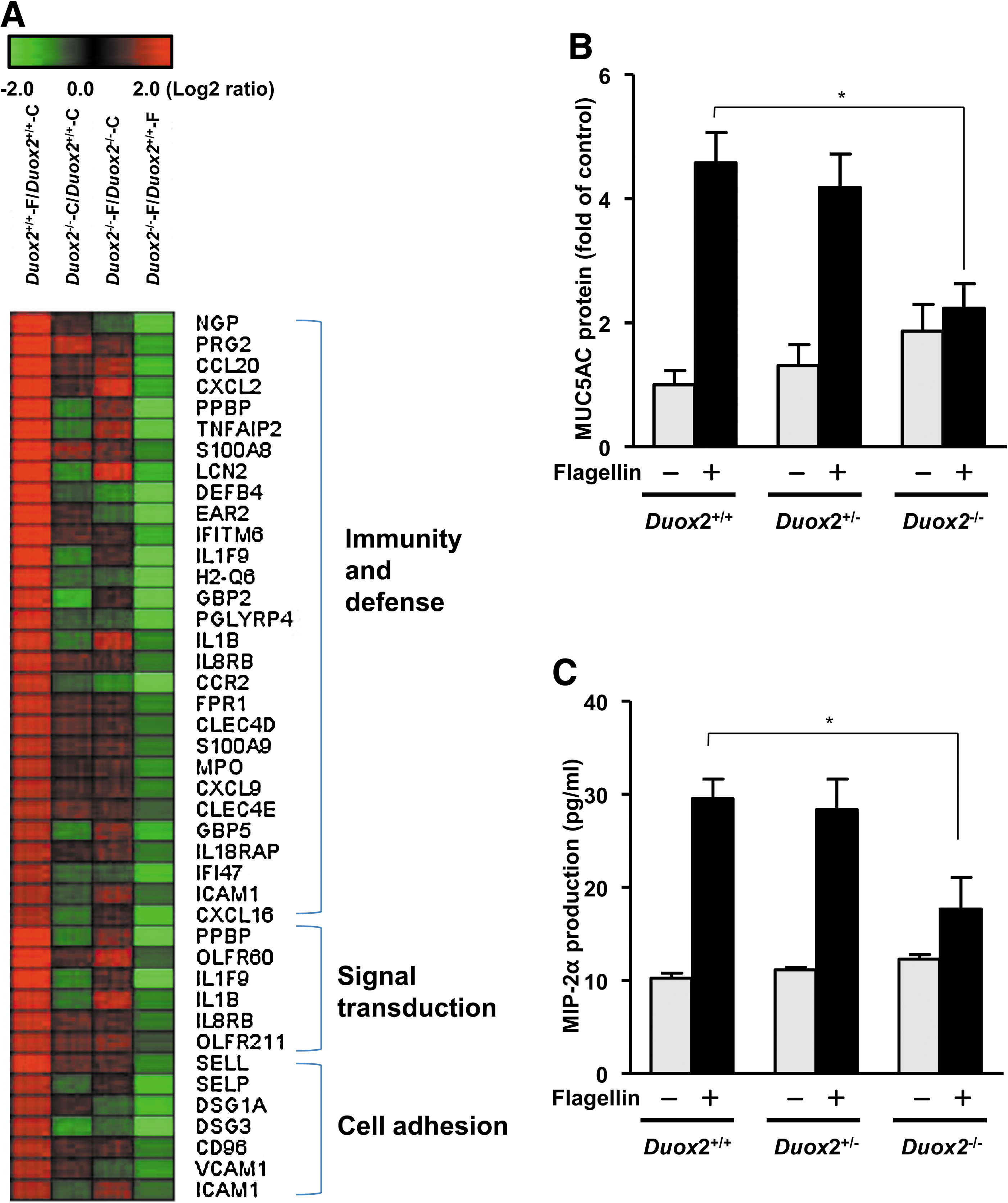
Wild type (Duox2 +/+) and Duox2 knockout (Duox2 −/−) mice were stimulated with 1 μg/ml of flagellin for 4 h. Two hundred three genes (greater than twofold) were upregulated in nasal mucosa of Duox2+/+ that was treated with flagellin and the full list of genes is presented in Supplementary Table S1. These genes included the following defense-related and immune response-related genes (Table 1 and Fig. 5A): cytokine/chemokine-related genes (CCL20, CCR2, CCR5, CXCL2, CXCL5, CXCL9, CXCL16, IL18RAP, TNFAIP2, IL1B EAR2, and FPR1), granulocyte-related genes (IL8RB, MPO, PRG2, PPBP, and PRG3), interferon-related genes (IFITM6 and IFI47), macrophage-related genes (IL1B and S100A9), and T-cell-mediated immune response-related genes (H2-Q6 and IL1F9). In addition, signal transduction (PPBP, OLFR60, and P2RY10) and cell adhesion (SELL, SELP, ICAM1, DSG1A, DSG3, and VCAM1) related genes were also increased by flagellin treatment. These genes were selected based on the biological processes and molecular functions of their gene ontology. However, the increase of inflammation and immune response related genes by flagellin treatment were diminished in the nasal mucosa of the Duox2−/− mice compared with that of Duox2 +/+ mice (Table 1 and Fig. 5A).
Flagellin-treated nasal mucosa of wild-type mice (C57BL/6).
PBS-treated nasal mucosa of wild-type mice (C57BL/6).
PBS-treated nasal mucosa of Duox2 knockout (Duox2−/− ) mice.
Flagellin-treated nasal mucosa of Duox2 knockout (Duox2−/− ) mice.
To confirm the microarray result, we measured flagellin-induced MUC5AC and MIP-2α (CXCL2) production in Duox2 knockout (Duox2 −/−) mice compared with that in Duox2 +/+ and Duox2 +/− mice. Although MUC5AC was not selected by twofold increased gene, flagellin-induced production of MUC5AC (Fig. 5B) and MIP-2α (Fig. 5C) were diminished in the nasal mucosa of the Duox2−/− mice compared with that of Duox2 +/+ and Duox2 +/− mice. However, the change in flagellin-induced MUC5AC and MIP-2α production was insignificant in heterozygous Duox2 +/− mice and in Duox2 +/+ mice.
Neutrophil infiltration and MUC5AC expression were reduced in the nasal epithelium and NALF of Duox2−/− mice
Increasing of neutrophil infiltration is hall mark in inflamed tissue. We performed histological analysis of neutrophil infiltration in nasal epithelium of Duox2 −/−, Duox2+/ −, and Duox2+/+ mouse. Mouse nasal epithelium was injected by challenging of flagellin and then subjected into immunohistochemical staining with antibodies to neutrophil and MUC5AC protein in mouse nasal mucosal epithelium. Neutrophil infiltration was significantly decreased in the nasal epithelium of Duox2 −/− mice and NALF of Duox2 −/− mice compared to Duox2+/ − and Duox2+/+ (Fig. 6A, B). Moreover, MUC5AC expression was abolished on the surface of the nasal mucosa of Duox2 −/− mice compared to Duox2+/ − and Duox2+/+ (Fig. 6C). Taken together, these results suggest that DUOX2 activation is essential for TLR5-induced inflammatory responses in nasal mucosal epithelium.

DUOX2 expression is upregulated in nasal mucosa of acute sinusitis
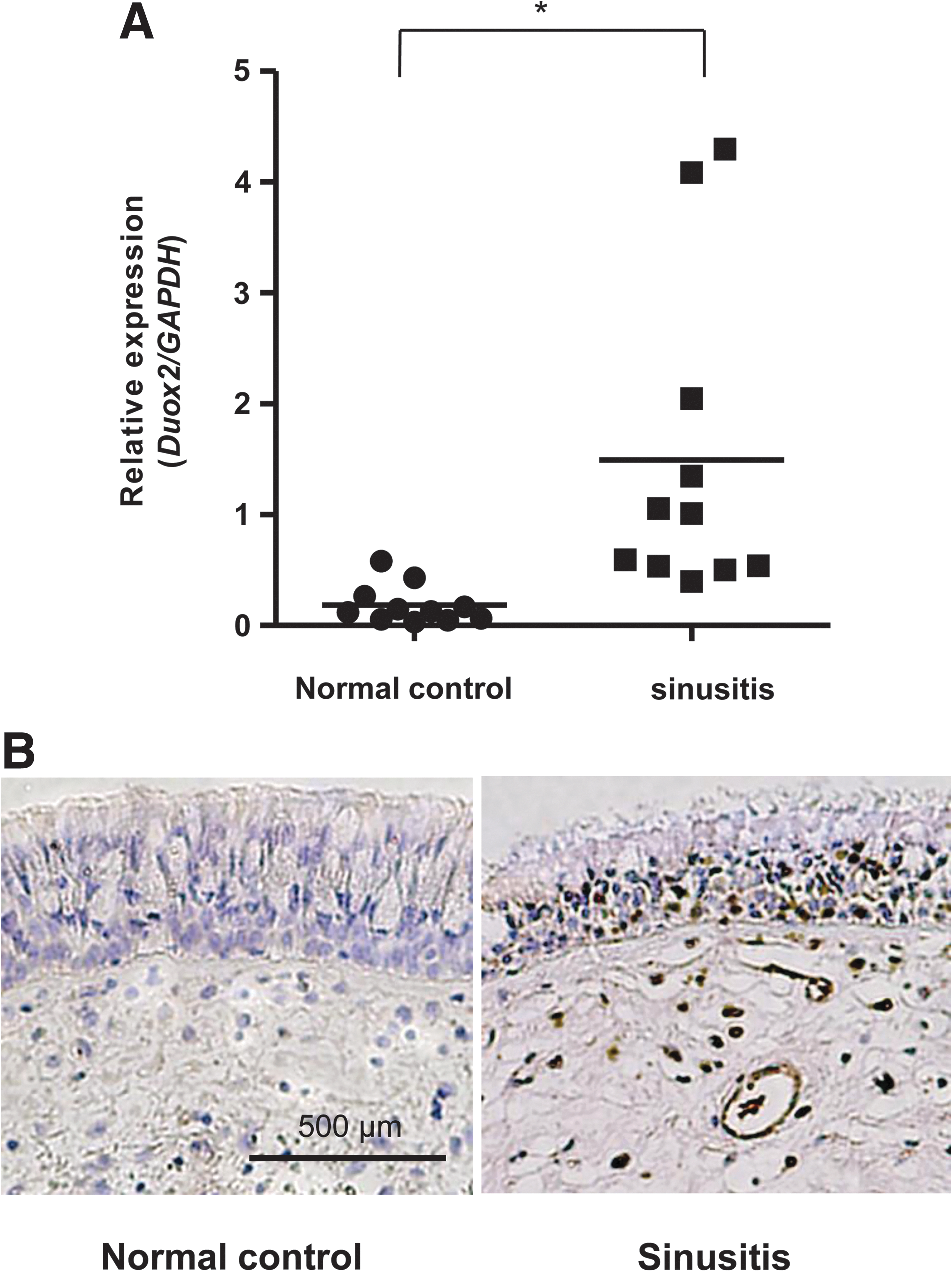
Consistent with critical role of DUOX2 in inflammation, we explored the level of Duox2 expression in nasal inflammatory biopsies of patients suffering from infectious sinusitis (nonallergic) in comparison to non-inflammatory tissue in nasal mucosa of healthy volunteers. Nasal mucosa from 11 adult patients with nonallergic sinusitis and 11 nonallergic normal adults were subjected into RT-PCR measuring the expression levels of Duox2 (Fig. 7A). Interestingly, we found a significant increase in the Duox2 gene expression level in sinusitis, compared to that in healthy adults. Moreover, neutrophil infiltration was increased in sinusitis (Fig. 7B). In conclusion, the upregulated expression of Duox2 seems to be linked to infectious inflammatory diseases in the nasal mucosa.
Discussion
The DUOXs so far are mainly known to be responsible for H2O2 generation, as seen in thyrocytes where they catalyze iodination of thyrogloubulin by stimulating thyroperoxidase (31). In addition to DUOX expression in the thyroid, DUOX1/2 isozymes are expressed in the ducts of the salivary gland, airway epithelia, and intestinal colon epithelium. Because DUOX has a peroxidase-like ectodomain in the NH3-terminal region, DUOX can directly generate H2O2 (10, 12). Substantial evidence indicates that both DUOX-dependent H2O2 and lactoperoxidase generate hypothiocyanate (OSCN−) from the oxidation of SCN−, providing a robust antimicrobial defense network in epithelial cells.
Although excess production of H2O2 by chronic infection and pathophysiological stress is detrimental to tissues, several recent reports have indicated that H2O2 is widely present as a key regulator in intracellular signaling related to immunity and inflammation (41, 43, 45). In this study, we first found that DUOX2-induced H2O2 is essential for the TLR5-dependent inflammatory response in primary NHNE cells and murine nasal mucosa; we tested DUOX1-knockdown NHNE cells and found no inhibitory effect on flagellin-induced H2O2 generation. However, when we silenced DUOX2 in the cells, H2O2 generation was inhibited in response to flagellin (Fig. 1F). Earlier studies have reported that DUOX1 is the major source of H2O2 in TLR-mediated cell signaling in the epithelial cells of the lower respiratory tract (9, 24, 41, 45). However, in NHNE cells, our results suggest that DUOX2 controls the flagellin-induced ROS generation and hence is involved in the inflammatory response. The discrepancy between those results and ours can be explained by a difference in experimental conditions. First, unlike the previous reports that used epithelial cells of the lower respiratory tract, we examined the flagellin-induced response in primary NHNE cells. Second, we focused on TLR5-mediated inflammatory responses induced by flagellin, whereas previous reports analyzed asialoGM1 function in epithelium (9, 45). The activating antibody against asialoGM1 (α-ASGM1) ligates the asialoGM1 receptor, not TLR5 (1). Our finding of the DUOX2 activation mechanism by flagellin stimulation is quite consistent with the recent researches for DUOX2-induced host defense mechanism in Drosophila gut immunity (17) and Helicobacter pylori–induced chronic inflammation of the gastric cavity (19). Third, DUOX1 and DUOX2 activities are mainly regulated by mobilization of intracellular calcium. However, additional mechanisms regulating their intrinsic activity are different; DUOX1 is positively regulated by the cAMP-dependent protein kinase A (PKA) cascade, whereas DUOX2 is highly induced by activation of PKC stimulated by PMA (39). It is well known that purinergic receptor stimulates PLCβ, leading to IP3 production for intracellular calcium mobilization and DAG generation for activation of PKC. Flagellin-TLR5 cascade may induce PLCβ-PKC pathway resulting in specifically DUOX2 activation in nasal epithelium. These recent results suggest that DUOX2 plays an important role in mucosal immunity in a wide range of species, from flies to humans.
Considering our data in conjunction with a previous study (14), we can hypothesize that a rapid and transient (5–10 min) increase in intracellular H2O2 may act as intracellular second messenger in TLR5-induced cell signaling, whereas an increase in H2O2, a result of a prolonged response to flagellin (>6 h), kills invasive microorganisms as part of the host defense mechanism. We also found that Duox2 expression was increased after 6-h treatment with flagellin (data not shown). However, increasing Duox2 expression could not explain the early and transient increase in H2O2 in response to flagellin stimulation. This disparity in ROS generation and Duox2 expression may indicate the existence of rapid and efficient regulation of DUOX2 activation. Several lines of evidence indicated that TLR5 induces intracellular calcium ([Ca2+]i) mobilization through ATP production as a purinergic agonist (9, 28). This observation explained why activation of DUOX containing EF hands can be regulated by ATP-dependent [Ca2+]i mobilization. These studies showed that H2O2 production in flagellin-stimulated cells is mediated by the sequential activation of TLR5, ATP release, activation of purinergic receptor, and an increase in intracellular calcium, which then binds to the EF hand of DUOX to stimulate its oxidase activity. Here we found that flagellin-stimulated [Ca2+]i mobilization and scavenging of [Ca2+]i in response to the addition of BAPTA and EGTA/thapsigargin suppresses flagellin-mediated H2O2 generation (Fig. 2, Supplementary Fig. S2). Yeast two-hybrid and immunoprecipitation analysis revealed that the COOH-terminal region of DUOX2 interacts with the COOH-terminal region of TLR5 (Fig. 2D–F). Moroever, immunoprecipitation analysis showed that an endogenous DUOX2 interacts with TLR5 (Fig. 2F). It is evident that DUOX2 and TLR5 constitute a multi-protein complex in a signaling cascade. Based on this observation, we propose that [Ca2+]i mobilization and DUOX2 localization in the TLR5 signaling complex serves proper activation of DUOX2 in response to flagellin stimulation.
In this study, we also evaluated expression of MUC5AC, an inflammation-related mucin and secretion of IL-8 as neutrophil attractant in flagellin-induced inflammatory responses to understand the roles of DUOX2-mediated ROS generation in NHNE cells. Pretreatment of NAC (an ROS scavenger), diphenyliodonium (a NOX inhibitor), or transfection of DUOX2-specific shRNA into NHNE cells resulted in inhibited expression of MUC5AC and secretion of IL-8 (MIP-2α in mice) in response to flagellin stimulation (Figs. 3 –5). We confirmed that flagellin failed to trigger an inflammatory response (MUC5AC expression and neutrophil infiltration) in the nasal mucosa of Duox2 −/− mice compared to that of wild-type and Duox2+/ − mice (Fig. 6). Interestingly, microarray results showed that many genes related to neutrophils were reduced in Duox2 −/− mice (Fig. 5). These results indicate that expression of inflammatory molecules was inhibited in Duox2 mutant mice resulting in the reduction of neutrophil infiltration.
Consistent with critical role of DUOX2 in inflammation, we explored the level of Duox2 expression in nasal inflammatory biopsies of patients suffering from infectious sinusitis (nonallergic) in comparison to noninflammatory tissue in nasal mucosa of healthy volunteers. Nasal mucosa from 11 adult patients with nonallergic sinusitis and 11 noninflamed normal adults (nonallergic) were subjected into RT-PCR measuring the expression levels of Duox2 (Fig. 7A). Interestingly, we found a significant increase in the Duox2 gene expression level in sinusitis, compared to that in healthy adults. The result suggested that the upregulated expression of Duox2 seems to be linked to infectious inflammatory diseases in the nasal mucosa. Moreover, our results are supported with the current study where exacerbation of chronic obstructive pulmonary disease is associated with overexpression of DUOX isozymes (40). Collectively, these results provide decisive evidence that oxidative stress is a critical factor in the development and progression of airway inflammation and DUOX2 plays a pivotal role in flagellin-induced inflammatory responses, including IL-8 production and MUC5AC expression in the airway epithelia.
Materials and Methods
Materials
NAC, allopurinol, dicumarol, and DPI were purchased from Sigma Aldrich. Purified flagellin, TLR5 antagonist (Anti-hTLR5-IgA), and its control antibody (control IgA1) were purchased from Invivogen. Unless otherwise stated, all other reagents were from Sigma.
Cell culture
After approval of the study protocol by the Institutional Review Board of the Yonsei University College of Medicine, human nasal middle turbinate specimens were obtained from healthy volunteers. The culture systems used for the NHNE cells have been described previously (23, 46). The air–liquid interface (ALI) was created on day 9 by removing the apical medium and feeding the cultures from only the basal compartment. The culture medium was changed daily after the ALI was initiated, and all experiments utilized NHNE cells 3 days after the creation of the ALI.
Mice
C57BL/6J (wild type) mice for the inhibitor experiment were purchased directly from Orient Bio, Inc. DUOX2 mutant mice were obtained from The Jackson Laboratory. The new, recessive thyd mutation arose spontaneously in a B6 (129)-Duox2 thyd/J mouse (Jackson Laboratory; Stock no. 005543). Molecular genotyping enabled direct identification of heterozygotes for simplified colony maintenance.
Methods
Quantitative RT-PCR
Quantitative RT-PCR was performed using TaqMan® universal PCR master mix. RT-PCR was performed using the PE Biosystems ABI PRISM® 7700 sequence detection system. Target-specific probe sets against Duox1 or Duox2 were purchased from Applied Biosystems. The relative quantity was obtained using the comparative threshold method and results were normalized against GAPDH as an endogenous control. Gene-specific primer and probe sets designed against the targeted molecules are listed in Supplementary Table S2.
Intracellular ROS assay by DCF-DA
ROS were measured by a method described previously (6). After the confluent cells were stimulated, they were washed with Hanks' balanced salt solution (HBSS) and incubated for 10 min in the dark at 37°Cin HBSS containing 10 μM DCF-DA (Molecular Probes). DCF fluorescence was measured using a Zeiss LSM 510 confocal microscope (Minneapolis, MN) at an excitation wavelength of 488 nm and an emission wavelength of 515–540 nm. Seven fields of each dish were randomly selected and the fluorescence intensity was measured with the Carl Zeiss vision system (KS400, version 3.0). The seven values were averaged to obtain the mean relative fluorescence intensity, and these means were used for comparisons. All experiments were repeated at least three times.
Preparation of recombinant FliC and mutant proteins
Salmonella enteritidis FliC flagellin gene (GenBank accession: No. M84980) was obtained by PCR. Wild-type FliC flagellin (AA 21-505) and deletion mutant form of FliC flagellin (AA 119-505) were cloned in the pMT/V5-His vector (Invitrogen) to generate C-terminal V5-His epitope-tagged recombinant protein. To secrete recombinant flagellin in the culture supernatant, the signal peptide of the Drosophila Spätzler gene (AA 1-22) was used by fusing directly to the wild-type or mutant FliC flagellin constructs. Drosophila Schneider cell line was used for the production of recombinant FliC. Transfection of these cells was conducted according to a standard protocol using CaPO4. Expression of FliC flagellin was induced in cells by the addition of CuSO4 to the culture medium at a final concentration of 500 μM for 48 h before harvesting. Recombinant proteins were purified from the culture supernatant by using Ni+-NTA resin according to the manufacturer's instructions. The recombinant fliC protein was eluted from the resin with 250 mM of imidazole, and dialyzed with 50 mM sodium phosphate buffer (pH 7, 1% glycerol, 48 h).
Knockdown of Duox1 and Duox2 using lentiviruses
For knockdown of Duox1 and Duox2, cells were infected with control lentivirus, Duox1 (sc-60550-V) or Duox2 (sc-60552-V) shRNA lentivirus (Santa Cruz, CA), according to the manufacturer's protocol.
Preparation of antibodies against Duox1 and Duox2
Three peptides (41-53 EHRWGSKGSRLQR, 364-373 NSYWSREHP, and 451-461 QDINPALSRSN) for Duox1 and two peptides (370-381 CNNYWIRENPNL and 421-432 SRTDYVASSIQR) are synthesized from Peptron. Polyclonal antibodies were generated from the injection of peptide mixture for Duox1 or Duox2 to rabbit (AbFrontier).
Preparation and analysis of NALF
Nasal passages were flushed with 0.5 ml of phosphate-buffered saline, of which an average of 0.45 ml was recovered into a 1.5 ml tube held to the nose of the mouse. Nasal wash fluid was centrifuged to remove cells and the supernatant was stored at−80°C until analysis. The cells collected from the NALF were counted in a hemacytometer. Two hundred microliters of cell suspension was dispersed onto slides using a cytospin machine set at 100 g for 5 min. Slides were stained using a Hemacolor stain set (Merck) according to the manufacturer's instructions.
Dot blot analysis for intracellular MUC5AC protein
MUC5AC protein was detected by a dot blot analysis as described previously (16). After treatment with flagellin, NHNE cells or mouse NALF were lysed with cell extraction buffer (Invitrogen) followed by sonication and centrifugation at 15,700 g for 10 min. Protein amounts were determined using the BCA protein assay (Pierce) and a dot blot analysis was performed.
Enzyme-linked immunosorbent assay of IL-8 secretion
The total level of IL-8 in cell culture supernatants was measured using a commercially available kit (R&D Systems) according to the manufacturer's instructions. Assays had a range of 10–2000 pg/mL.
Nonsurgical intranasal instillation for mice
Flagellin and other inhibitors were administered intranasally. For intranasal instillations, 25 μl of solution was applied drop-wise into each nostril.
Histology
The nasal tissue was prepared by removing the skin from the head before preserving in 10% formalin overnight, followed by decalcification for 5 h in Surgipath Decalcifier II solution. A coronal section immediately rostral to the eyes was embedded in paraffin, cut at 4 μm, and stained with hematoxylin and eosin by routine methods. For immunostaining, tissue sections were deparaffinized and rehydrated in graded alcohol. Nonspecific protein binding was blocked with 3% BSA and 2% goat serum in Tris-buffered saline (pH 8.0) with 0.2% Tween 20 (TBST) for 1 h. Primary antibodies were diluted in blocking buffer and incubated overnight at 4°C at a final concentration of 0.05 or 0.1 μg/ml for human and mouse tissue sections. For MUC5AC immunostaining, tissues were blocked with nonspecific background staining with blocking agent (Immunotech; 2391) and then incubated with mouse antibody to human MUC5AC (mouse monoclonal Ab 45M1, 2 μg/ml; Lab Vision Corp.). For neutrophil immunostaining, tissue sections were stained with the antibody (NIMP R14) Ly6g/Gr1 surface protein expressed in neutrophil (Abcam) or with control rat IgG, followed by detection with ABC-HRP kit and DAB substrate. Tissue sections were counterstained with hematoxylin, dehydrated in graded ethanol, and mounted for photomicrography.
Harvest of mouse nasal tissue
After euthanasia with a CO2 gas chamber and cervical dislocation, the skin at the nape of the neck was incised with fine dissecting scissors, and the incision rotated around the entire neck (4). This skin was then dissected interiorly and completely removed, leaving the anterior aspect of the skull base exposed. The remaining portion of the skull was removed to the posterior aspect of the nasal cavity. The scissor was then turned inferiorly to section the palate, and thus completely separated the septum and turbinate from the lateral nasal wall. Once harvested, the nasal septum and turbinate's mucosa from each side was gently elevated off the cartilage and bone and total RNA was isolated from these tissues using TRIzol (Invitrogen).
Patient samples
Normal, healthy biopsies were derived from patients who had undergone septoplasty without pathological conditions. Sinusitis samples were taken from the patients with chronic sinusitis who did not have histories of asthma, aspirin sensitivity, or cystic fibrosis. None of the patients had received immunosuppressive drugs within the previous 2 weeks. They all had negative results on the allergic skin-prick test. All patient-related procedures were approved by the Yonsei University, Severance Hospital's ethics committee. All patients agreed to participate by giving informed consent at least 24 h before the study.
Accession numbers
Array data are deposited at the Gene Expression omnibus. Accession number is GSE26135.
Statistic analysis
Significant differences between treatment groups were identified with a t-test. p-Values less than 0.05 were considered statistically significant.
Additional details on methods are provided in an online Supplementary Methods.
Footnotes
Acknowledgments
We thank Drs. Kyu-Bo Kim and Karl-Heinz Krause for critical reading of the article. This research was supported by the Basic Science Research Program of the National Research Foundation of Korea (2011-0001168 to J.H.Y.), by the National Core Research Center (grant R15-2006-020-00000-0 to Y.S.B), by World Class University program (R31-2008-000-10010-0 to Y.S.B), by National Research Laboratory program (grant number ROA-2007-000-20004-00 to Y.S.B) of the Ministry of Education, Science and Technology/Korea Science and Engineering Foundation, and by Korea Health 21 R&D program grant A084614 of Ministry of Health and Welfare. M.S.S and J.O.K are recipients of a BK21 scholarship.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.