
Abstract
Stem cell research has opened new and exciting possibilities in the biological and biomedical sciences, and holds great promise of impacting many areas of medicine. However, despite the rapid advancements of the last decade, the precise and efficient differentiation of stem cells into distinct cell types and tissues still remains a major challenge for the field. In an effort to reproduce biologically relevant differentiation niches, or to direct stem cell differentiation into specific cellular fates, many investigators have explored the effect of biomechanical stimulation on pluripotent cells. This review focuses on a particular type of biomechanical force, namely fluid shear stress, and our current knowledge on its ability to direct differentiation and modulate function of embryonic and somatic stem cells. Antioxid. Redox Signal. 15, 1463–1473.
Pluripotent and Somatic Stem Cells
Fluid Shear Stress
Fluid shear stress can be broadly defined as the frictional force generated by the movement of fluid on a surface. More specifically, fluid shear stress is defined as the stress (i.e., the force per unit area) that a moving fluid applies tangential to the solid boundary of its container (42). Fluid shear stress is measured as a force per unit area, and in biological systems is usually expressed in units of dynes/cm2 (Fig. 1).

A role for fluid shear stress in biology has been extensively investigated in the context of vascular biology where the continuous flow of blood constantly generates shear stress on the endothelial cells lining the inner surface of blood vessels. Initial studies between the 1960s and the early 1980s showed that the endothelium is responsive to shear stress (3, 10, 15 –17), opening a new area of investigation leading to the now recognized concept of fluid mechanical forces as critical regulators of vascular function (18, 41). Following these findings in the cardiovascular field, a role for fluid shear stress as regulator of cell function has been recognized in different biological systems where fluid flow is physiologically present, from the kidney tubules (12) to the canalicula in bones (6).
Fluid Shear Stress as a Tool for the Differentiation of Pluripotent Stem Cells
Fluid shear stress has been used as a tool to achieve differentiation of pluripotent cells (Fig. 2) into endothelial cells and, more recently, also into hematopoietic cell precursors. In most of the cases, shear stress has been applied on embryonic stem cells (ES) already subjected to initial differentiation steps. More specifically, the majority of the data has been gathered on murine embryonic stem cells differentiated to generate cells positive for Flk1, considered the earliest marker of precursors of both hematopoietic (28) and endothelial lineages (57). Nevertheless, shear stress has also been used as a tool to differentiate “naive” ES cells and in the context of other ES applications, which will be further discussed.

Effect of fluid shear stress on murine ES-derived Flk1+ cells
The first documented observations demonstrating that cell precursors derived from pluripotent cells can be further differentiated using fluid shear stress were published by Yamamoto et al. (55), documenting that murine ES (mES)-derived Flk1+ cells respond to shear stress upregulating markers of endothelial identity. In this study, Flk1+ cells were exposed to shear stress of 1.5 to 10 dynes/cm2 for 24 to 72 hours, demonstrating that mechanical stimulation increases Flk1+ cell proliferation and induces expression of the endothelial markers Flk1, Flt-1, VE-cadherin, and PECAM-1 both at the mRNA and protein level. Moreover, Flk1+ cells exposed to shear stress 1.5 dynes/cm2 for 24 hours showed increased ability to produce tube-like structures in collagen gels. While the molecular basis of the shear stress response was not fully investigated, the authors presented initial evidence suggesting mechano-activation of Flk1, as a key signaling event in this process (55). These results were expanded by the same research group in a subsequent study where exposure of Flk1+ cells to shear stress up to 20 dynes/cm2 was found to induce expression of the arterial endothelial marker EphrinB2 at the mRNA level and at the protein level. Concomitantly, the downregulation of the venous marker EphrinB4 was documented in these cells (34). In this second study, the authors presented a series of experiments with pharmacological inhibitors suggesting that the induction of this arterial marker is mediated by activation of the VEGF-Notch pathway. Interestingly, shear-stress-induced EphrinB2 upregulation obtained after exposure to mechanical stimulation for 24 hours was lost within 24 hours of culture under static conditions. These results suggest a role of shear stress in inducing and maintaining not only endothelial fate but also arterial endothelial identity.
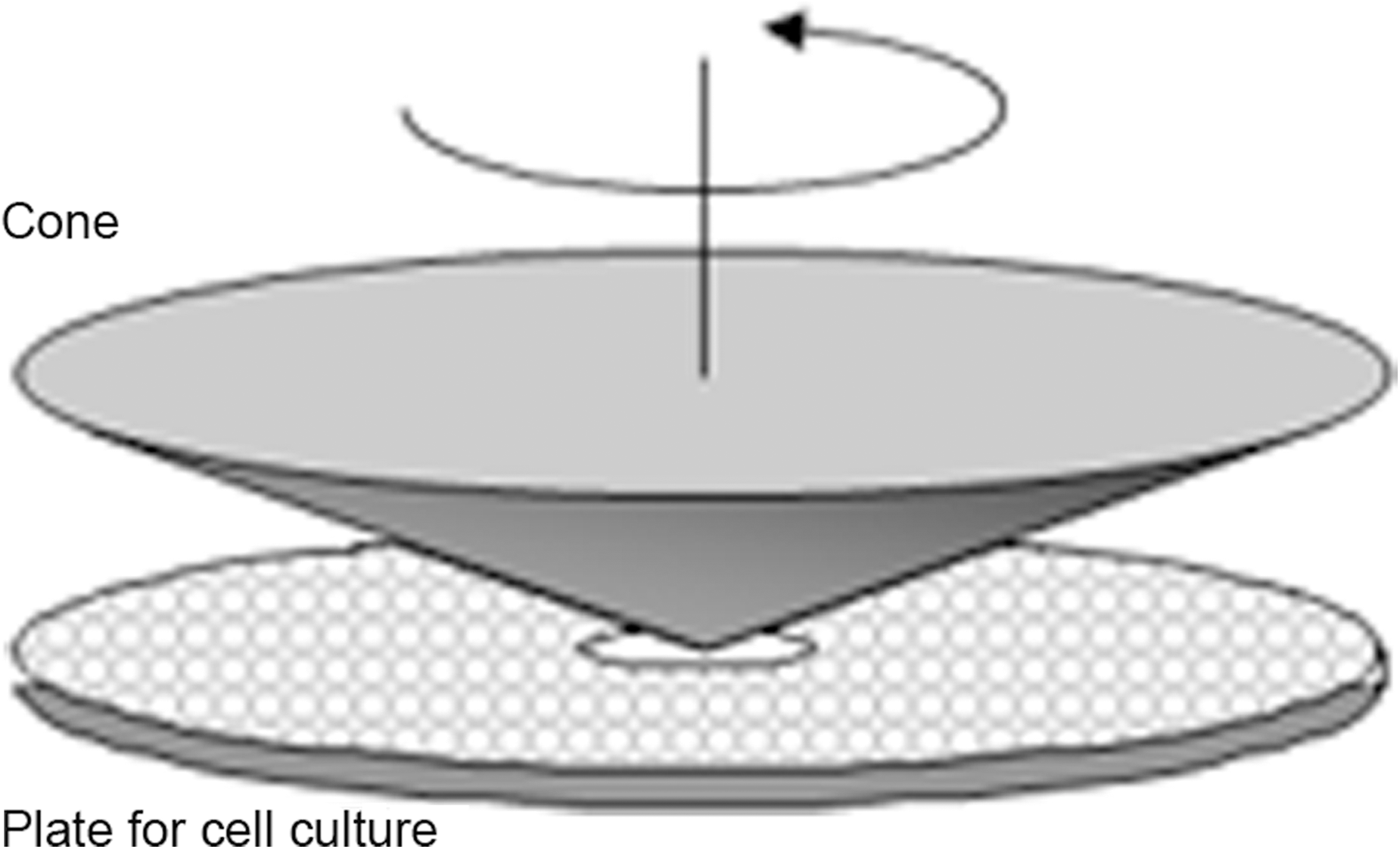
These two articles determined Flk1+ cells as shear stress-responsive cells and defined shear stress as a potential tool for the differentiation of mES-derived cells toward the endothelial phenotype. In contrast, Adamo et al. showed that under different culture conditions, shear stress can induce hematopoietic differentiation from Flk1+ cells (1). These authors cultured Flk1+ cells in the presence of hematopoiesis supporting cytokines and, taking advantage of a cone and plate fluid shear stress apparatus (Fig. 3), exposed them to a shear stress regimen similar to the one present in the mouse developing aorta soon after the initiation of heartbeat (ranging from 0 to 5 dynes/cm2 over 12 hours and stable at 5 dynes/cm2 for the following 36 hours). They found that, under these conditions, Flk1+ cells upregulate the expression of the transcription factor Runx1, a master regulator of hematopoiesis (38), and that these gene expression changes translate into an increase in the ability of Flk1+ cultured cells to grow hematopoietic colonies and to differentiate along the hematopoietic lineages in vitro (Fig. 4) (1). This observed cell differentiation recapitulates century-old observations showing that some of the earlier hematopoietic precursors originate from a specific kind of endothelium called “hemogenic endothelium”. The hemogenic endothelium is transiently present in specific sites of the vasculature of vertebrates only in a definite developmental time frame that coincides with the first hours after the establishment of heart beat. Thus, it is possible that the establishment of blood flow and the shear stress exerted on the hemogenic endothelium in the developing vertebrate embryo trigger its maturation. Interestingly, further work from the same authors on murine embryos (1) and observations in zebrafish embryos that fail to initiate the heartbeat (39) support this model. These results point at fluid shear stress as a potential tool for in vitro production of hematopoietic progenitors from ES derived cells.

The observations described above have used murine ES-derived cells. The use of fluid shear stress to differentiate human ES cells or iPS-derived cells has not been investigated with the exception of a study by Metallo et al. (37). In this study, the authors report that human ES cells differentiated into KDR+ endothelial cells respond to shear stress with change in morphology and upregulation of a number of genes including tPA, COX2, and SOD2. This response is similar to the one observed by the authors in postnatal endothelial cells (namely human umbilical vein endothelial cells and human microvascular endothelial cells). While the purpose of this study was to investigate the ability of ES-derived endothelial cells to respond to shear stress, these results suggest that fluid mechanical stimulation could be used to guide ES-derived endothelial cells toward the acquisition of gene expression patterns or functions typical of an endothelial phenotype.
Fluid shear stress in the context of other ES applications
In addition to being investigated as a tool to differentiate ES-derived precursors in the context of pluripotent stem cells research, shear stress has also been explored as a tool to affect the differentiation of “naive” undifferentiated ES cells (Fig. 5), and to test the effectiveness of methods for endothelial differentiation. Furthermore, the effects of shear stress have been documented in the context of differentiation protocols in which ES cells are exposed to multiple mechanical stimuli at the same time.
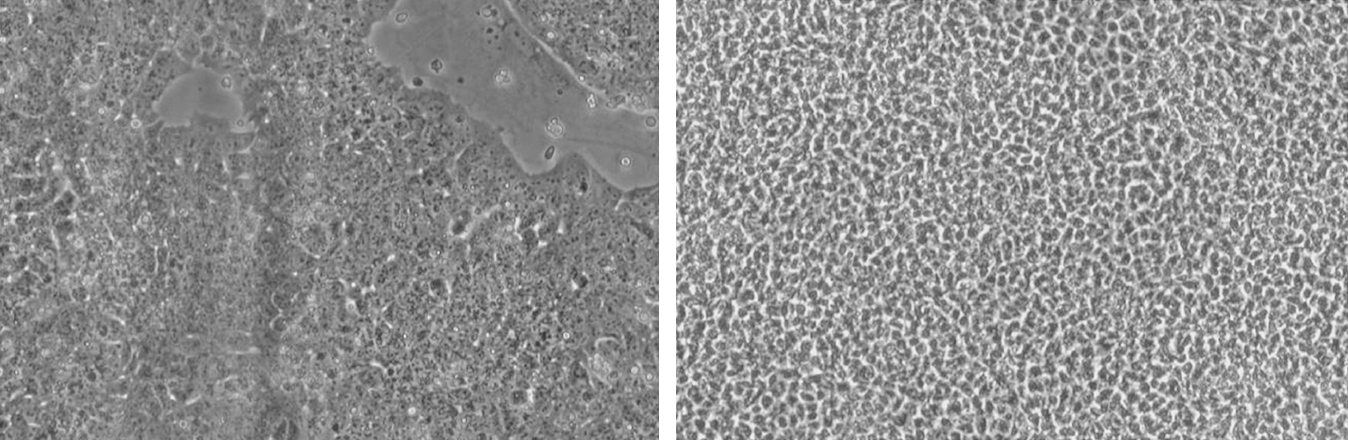
In order to study the effect of fluid shear stress on undifferentiated ES, Illi et al. exposed monolayers of murine embryonic stem cells to laminar shear stress 10 dynes/cm2 and showed that fluid mechanical forces induce epigenetic modification of histones and activation of expression of a small set of proteins typical of the cardiovascular tissue (27). Along these lines, Zeng L et al. found that undifferentiated ES cells exposed to laminar shear stress (LSS) 12 dynes/cm2 upregulate the expression of Flk1 and eNOS as well as the expression of surface markers compatible with endothelial identity (Fig. 6) and show an increased ability to generate tube-like structures in matrigel. In this latter study, the effect of shear stress was shown to be mediated via chromatin modification, with evidence suggesting that shear stress activated histone deacetylase 3 (HDAC3) through the Flk1-PI3K-Akt pathway (61). Importantly, at the time of exposure to shear stress, Zeng et al. plated undifferentiated cells on collagen type IV, a known potent inducer of vascular differentiation, and this condition may have played a role in the documented shear stress-mediated differentiation (57).

Kim et al. used shear stress on ES derived cells with a different approach. The authors in fact used fluid mechanical stimulation as a probe to characterize cells differentiated from ES cells without the use of mechanical stimuli. More specifically, Kim et al. differentiated endothelial cells from murine ESCs through the use of a drug resistance gene expressed under the control of endothelial promoters. They tested four different endothelial promoters: Flk1, Tie1, VE-cadherin, and CD31. They then used laminar shear stress to investigate whether the endothelial cells produced using each of the four different promoters had functional differences. Interestingly, when they challenged the different batches of ES derived endothelial cells with 16 dynes/cm2 for 48 hours, they found that only endothelial cells differentiated using Tie1 and CD31 as selective promoters were able to increase secretion of tissue plasminogen activator (tPA) upon biomechanical stimulation (29).
Shear stress has also been used on ES-derived cells in the context of 3D culture systems for ES differentiation as part of differentiation protocols based on the use of multiple types of biomechanical stimulation at the same time. Huang et al. exposed Flk1+ cells to both fluid shear stress and cyclic stretch in a microporous tube perfused with pulsatile flow and found that this complex biomechanical stimulation induces expression of the endothelial marker CD31 (26).
Fluid Shear Stress as a Tool for the Differentiation of Somatic Stem Cells
Mesenchymal stromal cells
Mesenchymal stromal cells (MSC) are multipotent cells that can be isolated from several human tissues. Initially derived from the bone marrow stroma, MSC have now been successfully isolated from a number of distinct tissues, including periosteum, muscle connective tissue, adipose tissue, amniotic fluid, and placenta. They usually account for a very low fraction of the tissue of origin and their functional role has been source of much deliberation. MSC isolation and identification is usually based on the absence of hematopoietic markers, together with ability to adhere and differentiate in at least three lineages, typically bone, adipose tissues, and cartilage (5).
Mesenchymal stromal cells have been shown to respond to shear stress, and mechanical stimulation has been used as a tool to direct their differentiation toward vascular and skeletal fates. Glossop et al. have shown that a rigorously defined MSC can respond to stimulation with shear stress of 1 dynes/cm2 for 1 hour and that this mechanical stimulation can induce gene expression changes in a number of members of the MAPK signaling pathway in as little as 10 minutes (20). Most of the other studies investigating the effect of shear stress on MSC, however, have been performed on loosely defined bone marrow-derived stromal cells (BMSC), a population expected to be enriched in proper MSC but not carefully characterized for expression of surface markers or multilineage differentiation potential.
Li et al. demonstrated that BMSC could respond to shear stress. Using a population of human bone marrow-derived stromal cells, not probed for surface marker expression and multilineage potential, they showed that oscillatory shear stress increases cell proliferation. The authors documented also a shear stress-mediated upregulation of the osteogenesis-related genes osteopontin and osteocalcin. Furthermore, they showed that fluid mechanical stimulation acutely increased intracellular calcium concentrations in BMSC (33). These investigators exposed cells to shear stress for 24 hours but did not define the frequency or the intensity of the laminar shear stress applied to the cultures. Some details about the specifics of fluid mechanical stimulation able to elicit a response in MSC were provided by Riddle et al., who exposed BMSC to oscillatory shear stress with frequency of 1 Hz and peak intensity of 5 to 20 dynes/cm2 for variable amount of times. The authors observed an increase in cell proliferation and acute increases in intracellular calcium proportional to the intensity of shear stress applied. They further reported a mechano-dependent activation of Erk1 and Erk2 as well as the activation of calcineurin, a calcium sensitive phosphatase (44).
Differentiation of MSC into osteoprogenitors
The results described in the previous section showed that MSC and bone marrow stromal cells, a cell population likely to be enriched in MSC, respond to shear stress. Could shear stress be used to differentiate these cells into osteoprogenitors? Initial investigations in this direction were performed by a number of different groups that cultured BMSC in the presence of shear stress and of one or more soluble factors that promote osteogenesis, namely dexamethasone, β-glycerophosphate, and ascorbate-2-phosphate. For example, Kreke et al. probed the effect of shear stress on culture of BMSC enriched only with the two latter pro-osteogenic factors. In their study, BMSC were exposed to 1.6 dynes/cm2 for 5 to 120 minutes every other day for up to 20 days. The authors did not have significant findings and reported that under these conditions shear stress induced only a modest increase in the production of osteopontin, a protein implicated in bone remodeling (31). In a follow-up study, the same group achieved a better outcome adding all three pro-differentiation factors to the cultures and stimulating them with shear stress at 2.3 dynes/cm2. In this work, the authors found that when cells were stimulated with fluid shear stress for 24 hours and allowed to differentiate in static conditions for 13 days, showed a 4- to 20-fold upregulation at the gene expression level of a number of markers of bone differentiation, including osteocalcin, osteopontin, and collagen1α; and moreover, showed a significant increase in osteopontin deposition at the protein level. Interestingly, shear stress was also found to increase PGE2, a suggested mediator of osteoblast proliferation and mineralization, and VEGF production as measured by ELISA on flow-conditioned media. No mechanism was provided for these shear-mediated changes but shear stress was shown to induce sustained phosphorylation of p38 and ERK (32). The same research group further extended these results showing a shear-mediated induction of secreted osteopontin and osteocalcin in BMSC. Importantly, the authors showed that pulsatile and continuous flow induced dramatically different changes in gene expression in stimulated cells when RNA was extracted right after the end of the mechanical stimulation and that the effects of shear stress were not transferable exposing cells to flow-conditioned media (46). These two pieces of evidence showed that BMSC respond differently to pulsatile and continuous flow and that the effect of shear stress is a direct effect on cells and it is not mediated by the production of soluble paracrine factors.
Other investigators have attempted to use different combinations of soluble factors and shear stress to induce BMSC osteogenic differentiation. Grellier et al. tested the effect of fluid shear stress on BMSC cultured with the addition of the only pro-osteogenetic factor dexamethasone. They found that exposure to laminar shear stress 12 dynes/cm2 for 90 minutes upregulates alkaline phosphatase activity and the expression of connexin 43, a gap junction protein previously shown to accumulate during osteoblastic differentiation. However, the use of these same experimental conditions led to the downregulation of collagen type 1 expression (22). In another set of experiments, Scaglione et al. (45) investigated the effect of shear stress on BMSC plated on different extracellular matrix. They found that when BMSC are stimulated with laminar shear stress (12 dynes/cm2 for 12 days) they deposit more mineralized matrix, but this effect, as well as shear stress-mediated changes in gene expression, are highly dependent on the type of surface coating used at the time of the initial plating. The results were particularly interesting for the osteopontin gene that was upregulated by shear stress in cells plated on a glass surface and downregulated in cells plated on calcium phosphate-coated glass. These results confirm the notion that shear stress can induce osteogenic differentiation of BMSC but also demonstrate that the effects of the mechanical stimulation are heavily dependent on culture conditions (45).
Taken together, these data document that fluid shear stress is capable of directing BMSC toward osteogenic differentiation.
Differentiation of MSC into vascular cells
Strictly defined, MSC have been previously shown to be able to differentiate into endothelial cells when exposed to VEGF (40). Building on this observation, Bai et al. (3a) isolated rat BMSC, which do not express surface markers of hematopoietic or endothelial lineage, and investigated whether exposure of these cells to shear stress in the presence or absence of VEGF could induce endothelial differentiation. The investigators exposed BMSC to shear stress ranging from 10 to 25 dynes/cm2 for up to 48 hours and showed that the lower level of shear stress tested (10 and 15 dynes/cm2) were more effective in inducing expression of the endothelial genes Flk1 and tPA at the 24 hour time point, and that this effect was additive to the gene expression increase induced by VEGF. However, the results were different at the 48 h time point. In fact, after 48 hours of shear stress stimulation, Flk1 and tPA expression levels went back down to static levels unless VEGF was present, suggesting that in these experimental condition VEGF might be necessary to maintain the shear stress mediated upregulation of VEGFR2.
In another set of experiments, Engelmayr et al. (13) investigated the ability of shear stress to differentiate BMSC into cardiac valve-like tissue. These investigators seeded BMSC onto thin strips of polymeric scaffold for tissue engineering (nonwoven polyglycolic acid and poly-L-lactic acid) and exposed cells to shear stress of approximately 1 dynes/cm2 in the presence or absence of cyclic stretch (circumferential strain) for up to 3 weeks. While shear stress alone simply induced cell proliferation, the combination of shear stress and cyclic stretch produced results suggestive of spatially organized expression of markers of differentiation of both endothelial and subendothelial tissues, including expression of CD31, vWF, and laminin (13).
Fluid shear stress in the context of 3D MSC differentiation models
While stem cells are commonly cultured on flat surfaces, cells have been cultured on three-dimensional scaffolds with the goal of building artificial tissues and organs (23). In the context of tissue engineering investigations, MSC have been seeded in three-dimensional scaffolds and then exposed to fluid shear stress inside this microenvironment with the ultimate goal of building vascular or bone structures.
As part of these efforts, Dong et al. (7) studied the response of MSC to shear stress in tissue engineered vascular grafts. To this end, they seeded canine-derived MSC cells on the inner aspect of tubular synthetic polymer scaffolds and then perfused the vascular structures with a peristaltic-pump-based system. The cells were exposed to shear stress slowly ramping from 1 to 15 dynes/cm2 over a 2-day period and then kept under constant 15 dynes/cm2 for additional 2 days. In this context, despite the absence of VEGF in the culture media, the authors observed an upregulation of the endothelial markers VE-cadherin and CD31 at the RNA level and an upregulation of the surface marker CD34 at the protein level. CD34 has been shown to be expressed on hematopoietic stem cells, endothelial progenitor cells, and endothelial cells, therefore the authors interpreted their result as a transition of MSC toward the endothelial phenotype (7).
In similar experiments, Zhao F et al. (62) and Stieheler M et al. (47) seeded human MSC in porous 3D scaffolds. The first group used a polyethylene trephthalate (PET) matrix and exposed the cells to osteogenic factors (dexamethasone, sodium-β-glycerophosphate, and ascorbic acid-2-phosphate), while the second group grew cells in a poly(D,L-lactide-co-glycolide) (PLGA) structure in the absence of osteogenic humoral stimuli. Both groups perfused the 3D porous scaffold with media and observed a perfusion-induced increase in cell number, alkaline phosphatase activity, and calcium content that they both interpreted as signs of flow-induced osteogenic differentiation. However, it is important to note that when porous 3D scaffolds are perfused with the flow rates used in these experiments, cells are exposed to minimal values of fluid shear stress that the authors themselves estimate to be in the order of 10−4 dynes/cm2. These values are several orders of magnitude lower that those found to be able to stimulate cells in 2D. Therefore, in the absence of further experimental evidence able to distinguish the relative importance of the different effects of perfusion within 3D porous structures (e.g., changes in gradients of grow factors and other extracellular molecules) the role of fluid mechanical stimulation in these experimental settings remains unclear.
Effect of fluid shear stress on mesenchymal stem cell lines
The studies addressing the effect of fluid shear stress on MSC previously reviewed here assessed the effect of this type of mechanical stimulation on primary MSC. However, a number of mesenchymal stem cell lines, or cell lines with behaviors similar to that of MSCs, have been recently developed. The effect of shear stress on two of these cell lines, C3H/10T1/2 and MC3T3-E1, has been investigated. The C3H/10T1/2 cell line is a cell line derived from whole mouse embryos that is characterized by fibroblast-like appearance, absence of chromosomal abnormalities, and ability to differentiate into adipocytes, chondrocytes, and myocytes (43). The MC3T3-E1 is a line derived from newborn mouse calvaria (upper portion of the skull) and appears to have a more restricted developmental potential. In fact, while this cell line has fibroblast appearance, it has been shown to be able to differentiate mainly into osteoblasts and osteocytes (48).
Wang et al. (52) exposed C3H/10T1/2 cells to laminar shear stress of 15 dynes/cm2 for 6 to 24 hours and then performed gene expression analysis. The authors found a shear-mediated downregulation of TGF-β signaling at the gene expression level as shown by the presence of shear-mediated downregulation of the mRNA for TGF-β1, TGF-β R1, and TGF-β R2, as well as of a number of other regulators of the TGF-β pathway such as Smad 2, 3, and 4 (52). Exposing the same cell line for 48 hours to oscillatory flow with peak intensity of 10 dynes/cm2, Arnsdorf et al. (2) found that fluid mechanical stimulation induces expression of the transcription factors Runx2, Sox9, and PPAR-γ accompanied with a slight increase in the activity RhoA as measured by an increase in GTP-bound RhoA and an increase in the activity of the RhoA substrate RockII. The authors interpreted these data as indicative of shear stress-mediated osteogenesis (2). McBride et al. followed a similar approach and studied shear stress-mediated gene expression changes in C3H/10T1/2 in order to obtain information on the role of shear stress in osteogenesis. They found that exposure of cells to a continuous shear stress 0.2 dynes/cm2 or 1 dynes/cm2 for 30 or 60 minutes induced upregulation of a number of ossification related genes, including collagen type I, collagen type II, Runx2, and Sox9. Interestingly, in this experiment the length of exposure to shear stress (30 or 60 minutes) was a stronger predictor of gene upregulation when compared to intensity of shear stress (35). Interestingly, exposing C3H/T101/2 to laminar shear stress (15 dynes/cm2) for 6 or 12 hours and culturing the cells on collagen type I, Wang et al. found a shear mediated upregulation of endothelial markers instead of bone differentiation markers. In particular, the authors described a shear stress-mediated upregulation of the endothelial markers CD31, von Willebrand factor, and VE-cadherin both as the mRNA and protein level, as well as a shear stress-mediated ability of these cells to perform characteristic endothelial functions such as uptake of acetylated-LDL and the formation of tubular structures in matrigel. Exposure to shear stress for 6 or 12 hours appeared to produce similar results with stronger biological effects observed at the 12 hour time point (53).
Experiments in a second mesenchymal cell line, the MC3T3-E1, have also been performed to assess the effects of shear stress on its differentiation. In a set of experiments, M. Thi et al. (50) exposed these cells to pulsatile shear stress, with a frequency 1 Hz, and average shear stress of 5 dynes/cm2 for 5 hours and then performed transcriptional profiling. In this context, shear stress was found to regulate ∼25% of the transcriptome. Analysis of gene expression showed a significant upregulation of VEGF and other pro-angiogenetic genes. However, none of these data were further validated with quantitative PCR, or any other technique, so it remains to be determined how the microarrays data translates into biologically significant events (50).
Endothelial progenitor cells
Endothelial progenitor cells (EPC) are somatic stem cells endowed with ability to differentiate into endothelial cells. Bone marrow and peripheral blood are considered among the major source of EPC (51). The surface markers pattern typical of EPC has been widely debated, and in the current literature EPC are often defined with a functional working definition as “blood-derived mononuclear cells that adhere to fibronectin and display typical endothelial functions, like uptake of acetylated low-density lipoproteins and binding of the Ulex lectin” (30).
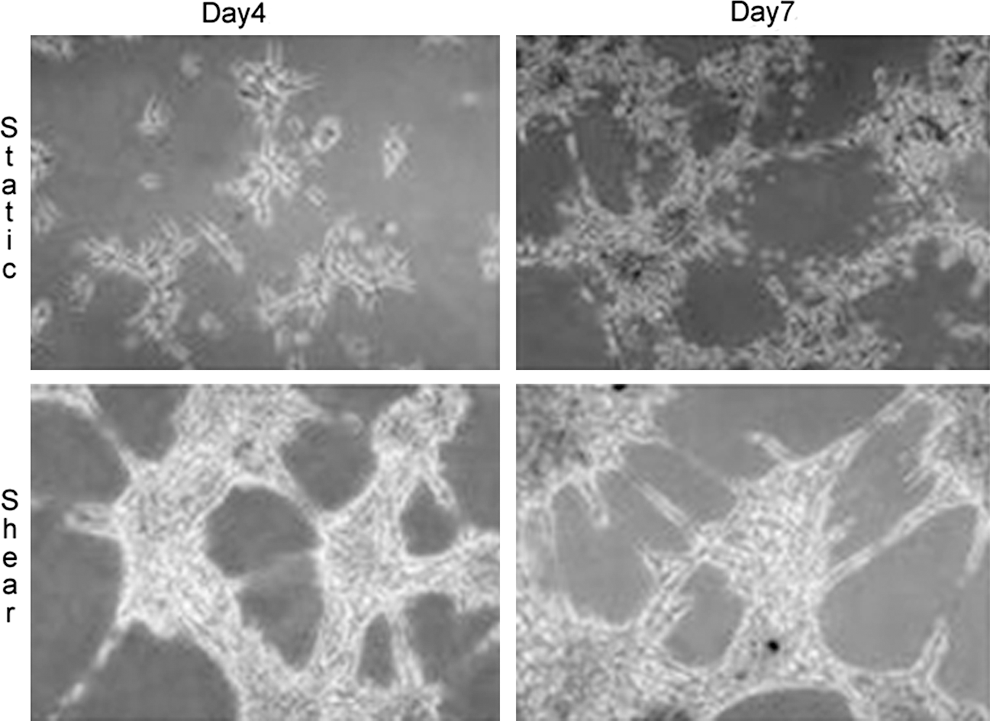
EPC have been shown to be able to respond to fluid shear stress, and fluid mechanical stimulation has been used to guide their differentiation towards a functional endothelial phenotype. One of the first groups to expose EPC to shear stress was Yamamoto et al. The authors isolated circulating EPC from human blood and exposed the cells to laminar shear stress with values ranging from 0.1 to 2.5 dynes/cm2 for 24 h. Interestingly, shear stress not only induced reversible changes in cell morphology, but it also induced cell proliferation and upregulation of the endothelial surface markers KDR (human analogue of the murine Flk1), Flt-1 and VE-cadherin, both at the mRNA and surface protein level. Moreover, shear stress significantly increased the ability of EPC to form tube like structures on collagen gels (Fig. 7). Critically, EPC exposed to shear stress were able to grow tube-like structures within 2 hours from seeding as opposed to the 4 day time frame required by EPC to grow similar structures when cultured under static (no flow) conditions (56). These results demonstrate that shear stress induces the maturation of EPC toward a fully differentiated endothelial cell phenotype.
This conclusion is now further supported by the results of three other groups who studied the effect of shear stress on the induction of specific endothelial functions in EPC, namely the anti-thrombogenic capacity, and the ability to react to oxidative stress challenges. Tao et al. (49) exposed EPC to shear stress from 5 to 25 dynes/cm2 for 25 hours in the context of porous polyurethane vascular grafts. These investigators showed that shear stress at levels of 10 dynes/cm2 or higher was able to induce secretion of the anti-thrombogenic protein tissue-type plasminogen activator (tPA) in a time-dependent manner. Importantly, Yang et al. (58) implanted the EPC-coated vascular grafts in the carotid of experimental dogs and showed that shear stress preconditioned grafts had a significantly higher patency rate at 2 months when compared with nonshear stress preconditioned conditioned controls (58). These observations were expanded by the same group in a subsequent study in which shear stress was found to slightly inhibit the secretion of the prothrombotic molecule plasminogen activator inhibitor-1 (PAI-1) and to increase the expression of the antithrombotic molecule prostacyclin (PGI2) (59). It was again this group of researchers that investigated the role of shear stress in modulating the ability of EPC-derived cells to modulate signaling pathways related to oxidative stress. The authors measured mRNA expression levels and activity of Cu/Zn superoxide dismutase in EPC exposed to shear stress and found that shear stress increases expression and activity of this important cyoprotective enzyme in a time- and intensity-dependent manner (49).
These experiments clearly showed that shear stress can induce endothelial differentiation of EPC. However, an important open question was whether shear stress is capable to direct the differentiation to arterial or venous endothelium. This question was elegantly investigated by Obi et al. (39a). These investigators exposed EPC to shear stress levels ranging from 0.1 to 5 dynes/cm2 and demonstrated that this type of mechanical stimulation induced the mRNA expression of markers of arterial endothelial identity (namely ephrinB2, Notch1, Notch3, Hey1, Hey2, and ALK1) while downregulated the marker of venous identity ephrinB4. The authors further showed that shear stress induced expression of ephrinB2 at the protein level.
Other type of somatic stem cells
As discussed in the previous sections of this review, most of the experiments performed assessing the effect of shear stress on somatic stem cells differentiation have focused on the effect of this biomechanical stimulus on the differentiation of mesenchymal stromal cells (MSC) or endothelial progenitor cells (EPC). However, fluid shear stress has been reported to be also a useful factor for the differentiation of a number of other somatic stem cells from a number of different tissues.
For example, Wu C et al. reported a role for fluid shear stress in the differentiation of human placenta-derived multipotent cells (PDMC) into endothelial cells. PDMC are multipotent cells with adipogenic, neurogenic, and osteogenic differerentiation potential that can be isolated from the human term placenta (60). By growing these cells in media enriched with endothelial grow factors and exposing them to laminar shear stress at 12 dynes/cm2 for 24 hours, the authors showed a slight induction of the endothelial markers vWF and PECAM1 at the mRNA and protein level. This combined mechanical and humoral treatment also induced a slight increase in the ability of the cells to perform typical endothelial activities (e.g., uptake of acetylated low density lipoproteins (Fig. 8) and formation of tube-like structures in matrigel). All these effects were suggestive of a potential role for fluid shear stress in the differentiation of PDMC toward vascular lineages (54). Importantly, similar results were obtained in experimental conditions used by Zhang P et al. that studied a different but “related” type of somatic stem cell: human amniotic fluid-derived stem cells (AFS). AFS are c-kit+ cells present in the amniotic fluid that have been shown to represent ∼1% of the amniotic fluid cells and to have the ability to differentiate into multiple lineages including adipogenic, osteogenic, neurogenic, and, in the presence of shear stress, the endothelial lineage (9, 14, 24).

Honda et al. have reported the use of shear stress in the context of tissue-engineered odontogenesis. Exposing cells harvested from the porcine molar tooth to shear stress in the context of 3D scaffolds, the authors reported a fluid shear stress-mediated increase in the expression of odontogenesis-related genes and increased size and ossification of fluid flow-stimulated grafts after in vivo implantation. Importantly, as in the case of the 3D systems discussed in the MSC section of this review, also in this case the expected shear stress values to which the cells were exposed in the context of the 3D scaffold used for seeding were extremely low and therefore, as previously discussed, it is difficult to assess the exact role of fluid mechanical stimulation in these observations (25).
Finally, shear stress has also been tested as a tool to differentiate adult adipose-derived stem cells (ASC). ASC are adherent multipotent cells found in the subcutaneous tissue (19). While fluid shear stress was not found to induce endothelial differentiation in this population (4), ASC were found to respond to shear stress both in the context of tubular scaffolds and when plated on flat surfaces. Progressively increasing magnitudes of shear stress were found to increase cell adhesion in tubular scaffolds (36), while stimulation of ASC on planar surfaces was found to induce VEGF and NO production (4).
Conclusions
Fluid shear stress has emerged as a strong regulator of multiple biological processes in many different cell types. Collectively, the studies reviewed here have documented that stem cells are not an exception to this principle. In fact, fluid shear stress elicits significant biological responses in both pluripotent stem cells and somatic stem cells. Moreover, it appears that this mechanical force induces differentiation by activating specific genetic programs and signaling pathways or that it promotes the acquisition of more mature cellular phenotypes in differentiating stem cells by modulating gene expression and complex cellular functions. Practically, these studies have uncovered the potential of using fluid mechanical forces as a “differentiation tool,” both per se and in conjunction with other extrinsic factors (e.g., growth factors and extracellular matrix) in order to modulate or significantly modify the biology of cultured stem cells.
Whereas several studies have investigated the effect of fluid shear stress on stem cells differentiation, many important questions remain unanswered. In fact, with few notable exceptions, including many of the studies on ES-derived Flk1+ cells and many of the studies on adult endothelial progenitor cells (EPC), most reports assessing the effect of shear stress on stem cells present limited sets of experiments that have not yet shed clear light on the biological significance of shear-stress-mediated differentiation of the studied cell population. In addition, there is a great variety of protocols for isolation/differentiation of stem cells and for the exposure to several types of shear stress (with variable magnitudes and times of exposure) that limits the ability to integrate in a systematic manner the outputs from the various studies. Collective efforts to reach consensus on the isolation and characterization of different types of stem cells, together with detailed definitions of the molecular, cellular, and functional effects of the shear-stress-mediated differentiation of stem cells are now necessary to fully develop the enormous potential of directing stem cell differentiation by mechanical forces.
Footnotes
Acknowledgments
The authors would like to thank William J. Adams for critical reading of this manuscript. Funding from NIH (RO1 HL-076686) supported this work.