
Abstract
Introduction
We recently demonstrated the presence of serum antibodies to the C-terminal subunit of RLIP76 (aaRLIP76) in a large percentage of patients with immune-mediated diseases characterized by endothelial dysfunction (14). In endothelial cells, aaRLIP76 increased intracellular levels of 4-HNE, decreased levels of GSH, and activated C-Jun NH2 kinase (JNK) signaling pathway, thus inducing oxidative stress-mediated endothelial cell apoptosis (14). As defects in the mechanisms of cell apoptosis have been associated with the pathogenesis and progression of vascular complications of several autoimmune diseases, it has been suggested that aaRLIP76 could play a key role in these diseases (10, 31).
Innovation
 RLIP76 is a Ral effector GTPase-activating protein expressed in endothelial and smooth muscle cells that catalyzes ATP-dependent transport and extrusion from the cell of anionic conjugates, leukotrienes, and weakly cationic compounds.
RLIP76 is a Ral effector GTPase-activating protein expressed in endothelial and smooth muscle cells that catalyzes ATP-dependent transport and extrusion from the cell of anionic conjugates, leukotrienes, and weakly cationic compounds.
 Autoantibodies to RLIP76 can impair the extruding function of this pump, thus leading to cell injury. This effect was exacerbated under mild oxidative stress, mimicking an inflammatory state.
Autoantibodies to RLIP76 can impair the extruding function of this pump, thus leading to cell injury. This effect was exacerbated under mild oxidative stress, mimicking an inflammatory state.
 Vascular cells from females appeared as more susceptible to the effects of these autoantibodies.
Vascular cells from females appeared as more susceptible to the effects of these autoantibodies.
 As serum autoantibodies to RLIP76 have been previously found in a large percentage of patients with immune-mediated diseases characterized by endothelial dysfunction, the impairment of RLIP76 by specific autoantibodies can play a role in the damage of vascular cells.
As serum autoantibodies to RLIP76 have been previously found in a large percentage of patients with immune-mediated diseases characterized by endothelial dysfunction, the impairment of RLIP76 by specific autoantibodies can play a role in the damage of vascular cells.
 This could be of major relevance in female subjects, contributing to the pathogenesis of immune-mediated vascular diseases.
This could be of major relevance in female subjects, contributing to the pathogenesis of immune-mediated vascular diseases.
 Findings reported in this work open a new perspective in the gender-dependent pathogenetic mechanisms of autoimmune diseases characterized by vascular dysfunction.
Findings reported in this work open a new perspective in the gender-dependent pathogenetic mechanisms of autoimmune diseases characterized by vascular dysfunction.
It is well known that several autoimmune diseases characterized by vascular dysfunctions, for example, systemic lupus erythematosus (SLE), systemic sclerosis (SS), and rheumatoid arthritis (RA), show a marked sex-associated difference, in terms of onset and progression, whose mechanisms remain unknown. In the present work, we analyzed the possible implication of “cell-sex” in this issue. In fact, although very few data are so far available in the literature regarding the possible role of “cell sex” (i.e., XX and XY cells) in determining cell fate, a gender difference in the apoptotic susceptibility, associated with differences in reactive oxygen species (ROS) production, has been recently suggested as concerns rat vascular smooth muscle cells (VSMCs), a key cell type in the maintenance of vascular integrity and function (12, 13, 15).
In this study, we investigated (i) the effects of aaRLIP76 on vascular cells, that is, rat VSMCs and human endothelial cells from males or females, and (ii) the implication and influence of estrogen in this system.
Results
RLIP76 expression and cell sex
We first investigated the surface expression of RLIP76 in freshly isolated VSMCs and human umbilical vein endothelial cells (HUVECs) from females (FVSMCs and FHUVECs) and males (MVSMCs and MHUVECs). Results concerning HUVECs, generally overlapping with those obtained in VSMCs, have been shown in Supplementary Figure S1A (Supplementary Data are available online at
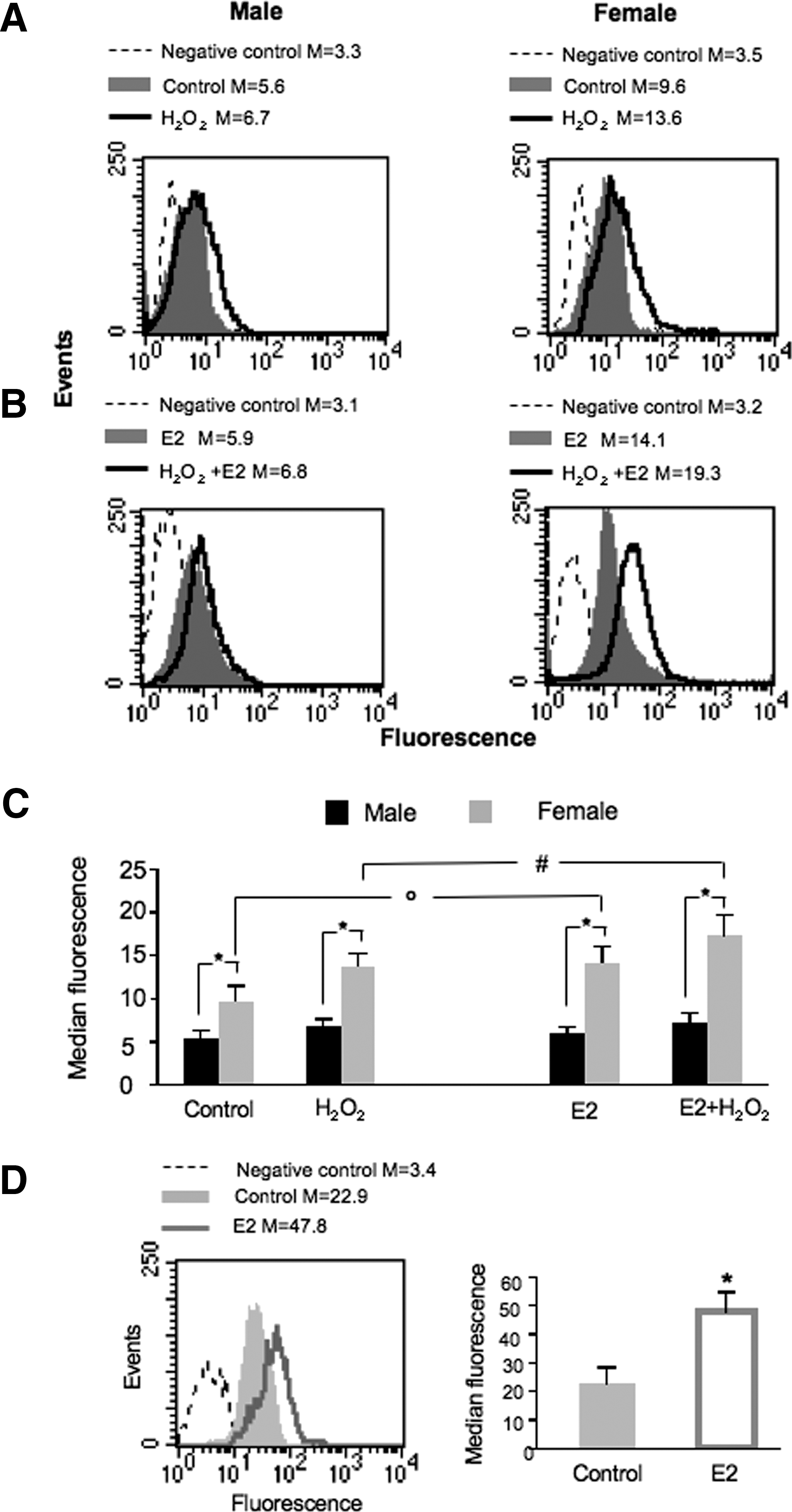
In consideration of the importance of RLIP76 expression and function in the maintenance of the redox balance inside the cells (14), we evaluated RLIP76 surface expression in response to mild oxidative stress, which normally occurs during the inflammatory response (3). Exposing cells from males and females to mild oxidative stress, using low concentrations of hydrogen peroxide, we found that mild oxidative stress induced an increase of RLIP76 membrane expression, as previously demonstrated (14) (Fig. 1A, C). As far as sexual hormone was concerned, we observed that E2 was able to induce a further significant increase of RLIP76 at the plasma membrane in cells from females only (e.g., about +30% vs. H2O2, p<0.01) (Fig. 1B, C).
aaRLIP76, cell sex, and oxidative stress
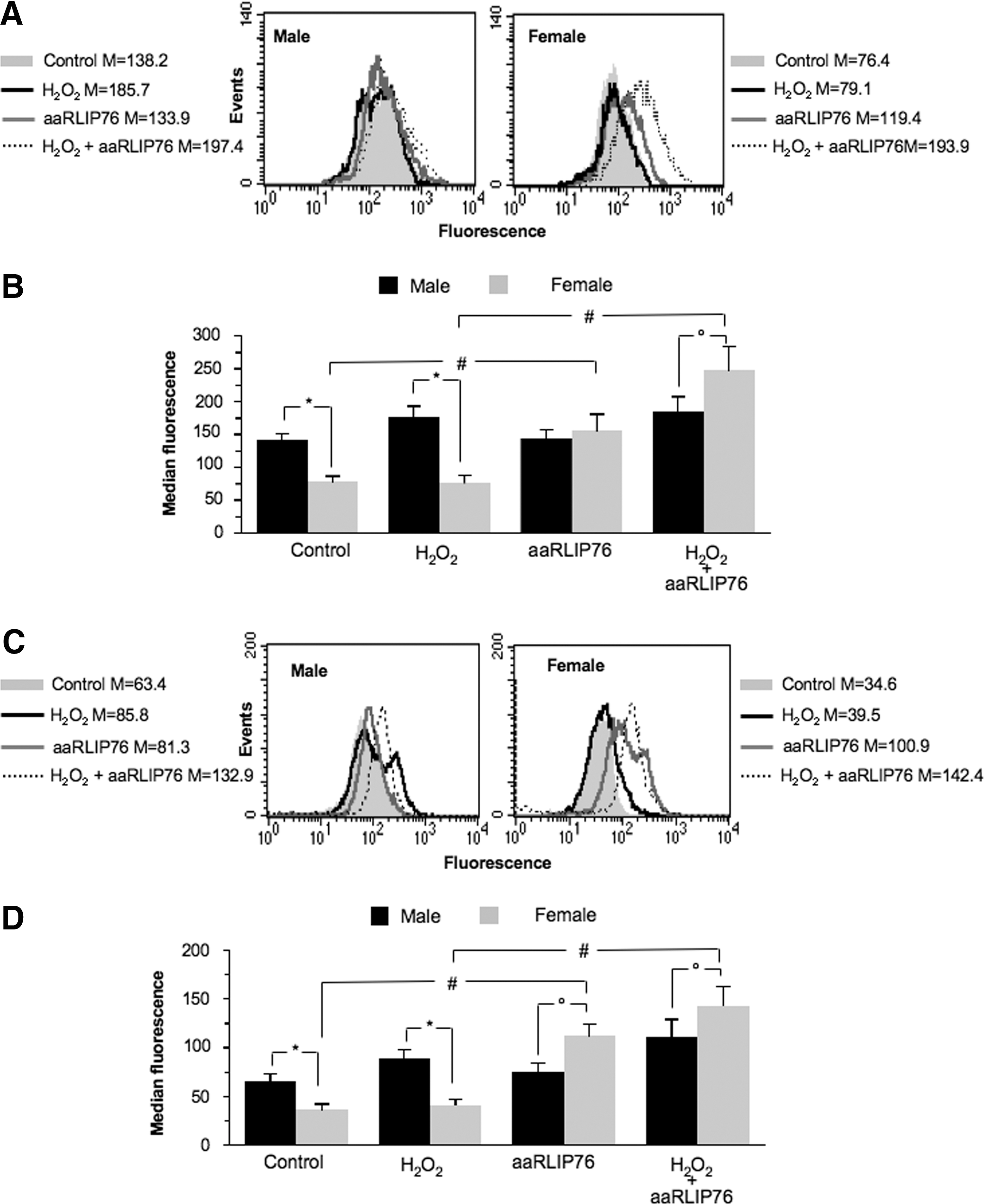
We also evaluated by flow cytometry the response to oxidative stress and to aaRLIP76-treatment by monitoring the ROS generation (Fig. 2). We observed the presence of “basal” differences between cells from males and females in terms of ROS production. In particular, even in the absence of any stimulation, either H2O2 (Fig. 2A, B) or O2 • (Fig. 2C, D) production was significantly higher in cells from males than in cells from females. In the same vein, after stress, MVSMCs displayed a higher production of ROS than FVSMCs (about +25% H2O2 and +26% O2 •) with respect to their basal parameters (Fig. 2B, D, left panels). In FVSMCs the formation of H2O2 and O2 after oxidative stress was similar to that found in untreated cells (Fig. 2B, D, right panels). Strikingly, when aaRLIP76 were added to the culture medium after oxidative stress, ROS were found to be significantly increased in FVSMCs (Fig. 2B, D, left panels; about +150% H2O2 and +240% O2 •, p<0.01) but not in MVSMCs (Fig. 2B, D, right panels; about +15% H2O2 and +60% O2 •).
RLIP76 participates to the extrusion from the cell of GSH conjugates, such as GS-HNE (19). We thus analyzed the formation of 4-HNE adducts with histidine by quantitative cytofluorimetric analysis (Fig. 3A, B) and fluorescence microscopy (Fig. 3C) with the aim to point out the mechanisms underlying the aforementioned effects. According to higher expression of surface RLIP76 in cells from female (Fig. 1), 4-HNE levels were significantly higher in MVSMCs than in FVSMCs either in physiological conditions or under mild oxidative stress (“*” in Fig. 3B). The incubation with aaRLIP76 induced a significant intracellular increase of 4-HNE levels in FVSMCs only (“#” in Fig. 3B). These results were thus in line with those reported in Figure 2, indicating a powerful activity of aaRLIP76 in hindering RLIP76 function, particularly in cells from females.
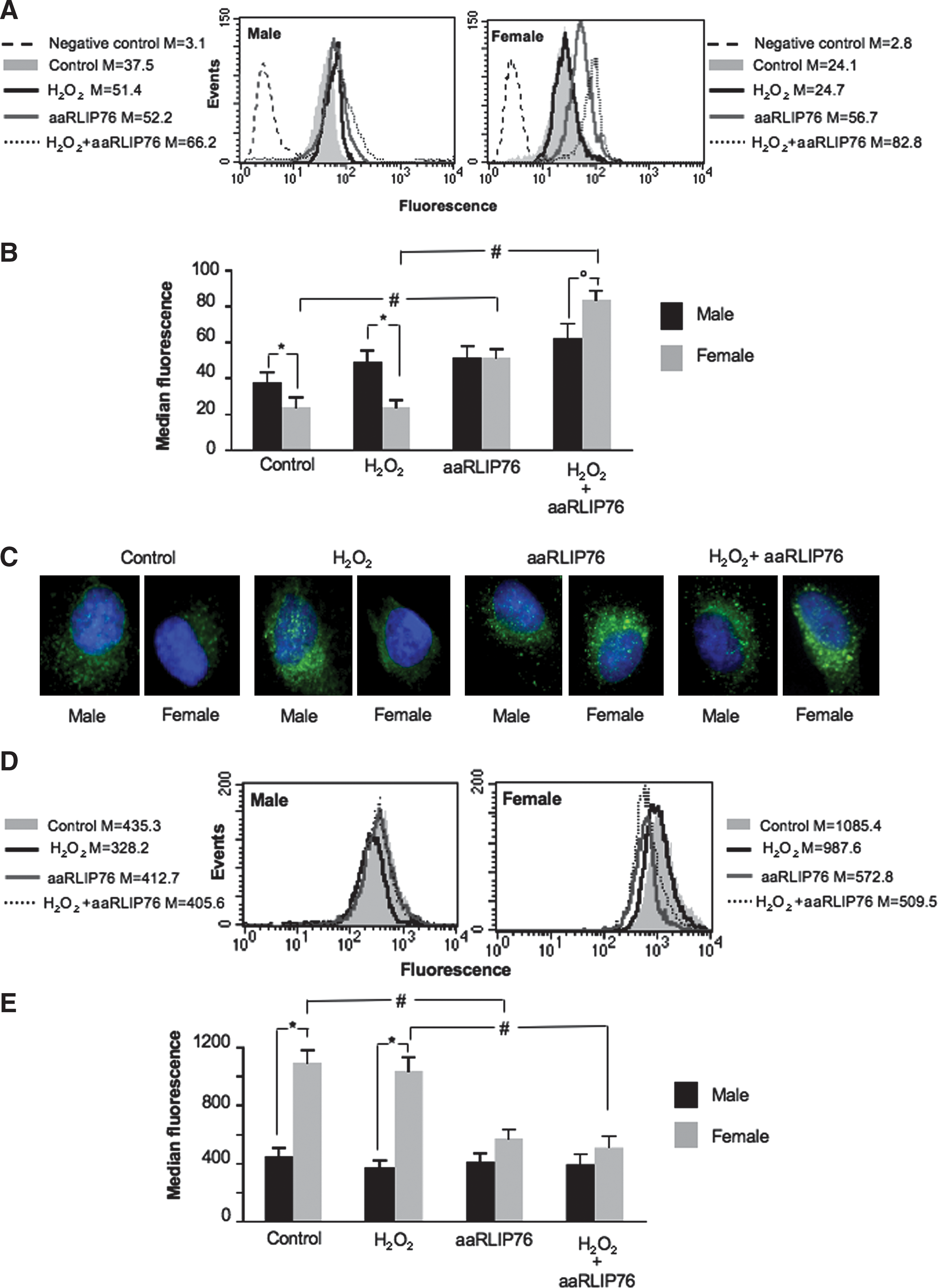
Because the detoxification of intracellular 4-HNE involves the most abundant cellular thiol-containing peptide, GSH, we analyzed intracellular GSH in the experimental conditions described earlier (Fig. 3D, E). We found that the basal levels of GSH were significantly higher in FVSMCs than in MVSMCs (+145%) and that, after mild oxidative stress, GSH levels remained substantially unchanged. Importantly, aaRLIP76 did not modify GSH intracellular level in MVSMCs, whereas they dramatically reduced the GSH content in FVSMCs either in physiological conditions or under mild oxidative stress (“#” in Fig. 3E). Hence, the homeostasis of cells from females seems to greatly depend upon RLIP76 pump function. Similar results were obtained in HUVECs as shown in Supplementary Table S1.
aaRLIP76, cell sex, and apoptosis
It has been reported that the reduction in endogenous GSH is associated with increased apoptosis (19). On these bases, apoptosis was evaluated in different experimental conditions (Fig. 4). We found a significantly higher spontaneously and oxidatively induced apoptosis in MVSMCs with respect to FVSMCs (*p<0.01) (Fig. 4A, C). The addition of aaRLIP76 to the medium did not significantly alter the percentage of apoptosis in cells from males (Fig. 4C). In contrast, aaRLIP76 were able to induce apoptosis in a significant percentage of FVSMCs either under physiological conditions or after mild oxidative stress (# p<0.01, Fig. 4C). Completely overlapping results were obtained in HUVECs as shown in Supplementary Figure S1 and summarized in Supplementary Table S1. Hence, vascular cells (either smooth muscle or endothelial cells) from females appeared as more susceptible to aaRLIP76-mediated apoptosis than vascular cells from males.
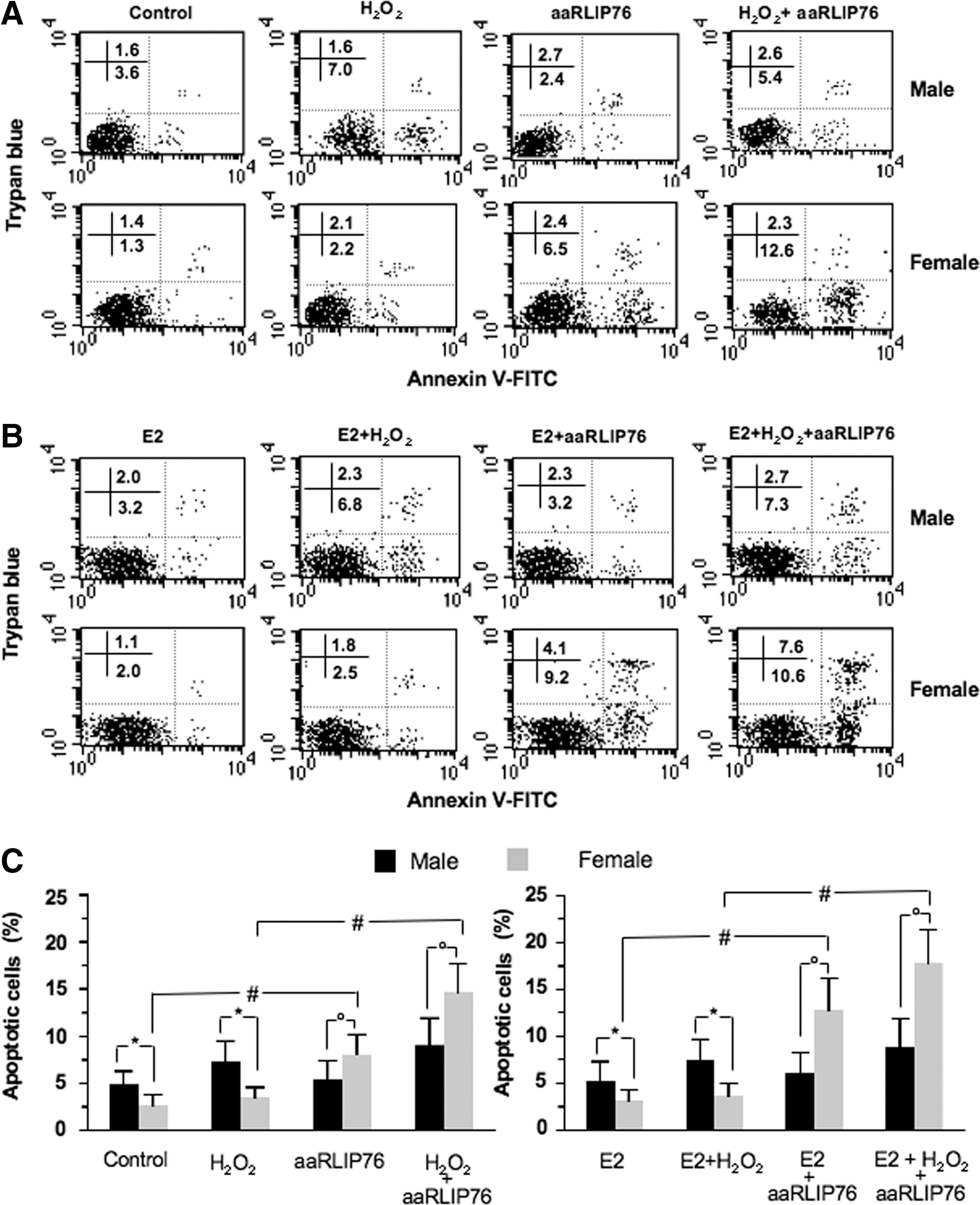
On the basis of the results reported in Figure 1, we also evaluated the effects of estrogens on aaRLIP76-induced cell death. Interestingly, we found that E2 significantly increased apoptosis induced by aaRLIP76, either in physiological or under oxidative stress conditions, but in cells from females only (representative experiment in Fig. 4B; Fig. 4C, right panel). The effect of E2 on aaRLIP76-induced apoptosis was also observed in HUVECs, as shown in Supplementary Figure S2.
aaRLIP76, cell sex, JNK, and caspase 3
To understand the mechanisms of apoptosis induced by aaRLIP76, we studied the role of JNK as signaling molecule in aaRLIP76-induced apoptosis in VSMCs. In fact, the typical signaling pathway induced by oxidative stress is associated with the activation of JNK, and in turn, 4-HNE plays an important role in oxidative stress–induced JNK activation (4, 36). As reported in Figure 5, treatment with aaRLIP76 leads to a significant phosphorylation of JNK in cells from females only, as demonstrated by quantitative flow cytometry analysis either in physiological conditions or after mild oxidative stress (Fig. 5B). In contrast, in MVSMCs no significant difference was detectable among the different experimental conditions. The same trend was observed in endothelial cells (HUVECs, Supplementary Table S1). Treatment with aaRLIP76 also significantly increased the activity of caspase 3 in FVSMCs, as clearly shown in Figure 5A and 5C.
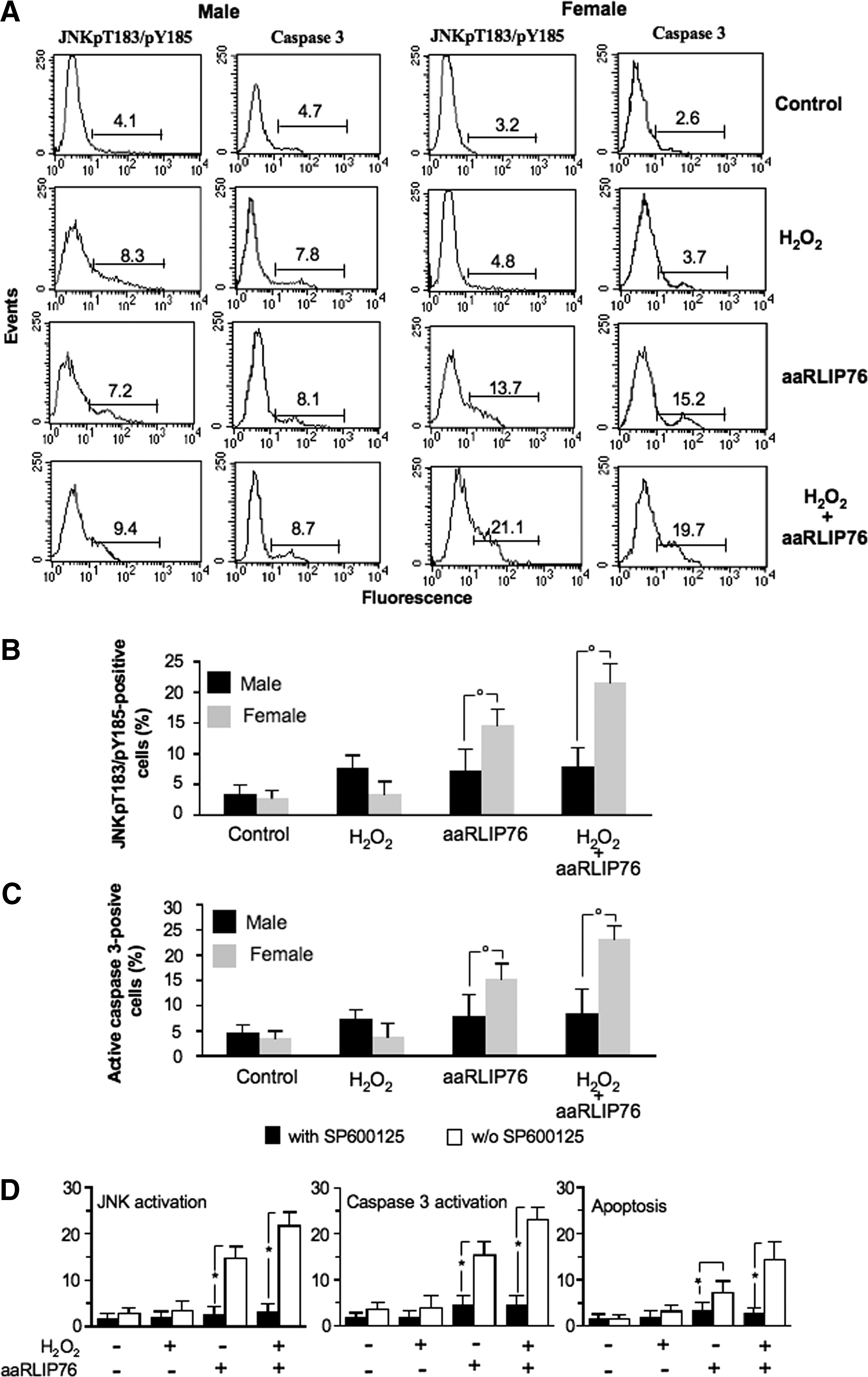
We also investigated the relationship between the JNK and caspase 3 signaling pathways in aaRLIP76-induced apoptosis by using the compound SP600125, a reversible ATP-competitive inhibitor of c-Jun phosphorylation, administered before the addition of aaRLIP76 (Fig. 5D). We found that pretreatment of cells from females with SP600125 significantly reduced, as expected, JNK activation, caspase 3 activation, and apoptotic rate either in physiological conditions or under conditions of mild oxidative stress (about −75% and −80%, respectively). In contrast, the compound SP600125 did not induce any considerable effect in cells from males (not shown).
Effect of RLIP76 silencing by siRNA
To clarify the role of RLIP76 expression level in aaRLIP76-mediated apoptosis, we transfected either VSMCs or HUVECs with RLIP76 siRNA before apoptosis triggering. In our experimental conditions, we found that 48 h after RLIP76 siRNA addition about 70% of both FVSMCs and MVSMCs (Fig. 6A, left panels) and 65% of FHUVECs and MHUVECs (not shown) were transfected. In RLIP76 siRNA-transfected cells, a significant reduction of RLIP76 surface expression, with respect to nonsilencing siRNA transfected cells, was detectable (Fig. 6A, right panels). Consequently, we also found that, after treatment with E2, either surface expression or total amount of RLIP76 was significantly decreased in RLIP76-silenced FVSMCs. In particular, after E2 treatment we found that (i) RLIP76 surface expression decreased (−86.5%±6.8%, p<0.01 for RLIP76 siRNA vs. nonsilencing siRNA), and similarly, (ii) the total amount of RLIP76 also decreased (−81.8%±7.3%, p<0.01 for RLIP76 siRNA vs. nonsilencing siRNA).

Further experiments were carried out in physiological conditions to reveal whether silencing of RLIP76 induced an alteration of cell homeostasis. We found a significant decrease of GSH in FVSMCs (−20.6%±4.3%, p<0.01 vs. nonsilencing siRNA transfected cells), but there was no increase of both apoptosis and 4-HNE levels (Fig. 6B, C). In contrast, under oxidative stress, the silencing of RLIP76 determined a significant increase of 4-HNE levels and apoptosis (p<0.01) in FVSMC (Fig. 6B, C). Notably, RLIP76 silencing significantly reduced susceptibility of FVSMC to aaRLIP76, either in physiological conditions or after mild oxidative stress, in terms of apoptosis (Fig. 6B). Conversely, RLIP76 silencing did not significantly influence 4-HNE production and apoptotic susceptibility of MVSMCs in any experimental condition analyzed here (Fig. 6B, C). Further experiments were carried out to evaluate the effects of RLIP76 silencing on JNK activation. We found a significantly reduced JNK activation in FVSMCs only (−68.2%±7.2% in physiological conditions and −73.0%±4.7% after mild oxidative stress). Similarly, in FHUVECs, RLIP76 silencing increased oxidative stress–induced apoptosis (of about 140%) and reduced aaRLIP76-induced apoptosis (of about 55%). Finally, control experiments carried out with E2 clearly indicated that this hormone did not affect cell susceptibility to apoptosis in RLIP76-silenced cells (Supplementary Fig. S2).
Gender differences in cell vulnerability to aaRLIP76
Statistical analysis pointed at a significant difference between cells from males and females, either VSMCs (Table 1) or HUVECs (Supplementary Table S1), for all parameters considered in this work. In particular, cells from females showed a higher susceptibility than cells from males to aaRLIP76 treatment, for example, in terms of apoptotic susceptibility, either when aaRLIP76 treatment was performed in physiological conditions or under a mild oxidative stress.
Statistical analysis of the differences between FVSMCs and MVSMCs in (i) surface expression level of RLIP76 either in physiological conditions or under oxidative stress and (ii) changes of different parameters in response to aaRLIP76 treatment performed under physiological conditions and after mild oxidative stress.
FVSMCs, vascular smooth muscle cells from female rats; GSH, reduced glutathione; JNK, C-Jun NH2 kinase; MVSMCs, vascular smooth muscle cells from male rats.
Discussion
It has been reported that oxidative stress can induce a rapid, transient induction of Ral-binding GTPase-activating protein (RLIP76), which catalyzes ATP-dependent transport of GS-HNE (14). In the present work, we analyzed the effects of aaRLIP76 in freshly isolated vascular smooth muscle cells and in endothelial cells from male and female. We found a different susceptibility in terms of ROS formation and a different propensity to undergo apoptosis, vascular cells from “females” being more susceptible to the effects of aaRLIP76. In addition, under mild oxidative stress, to some extent mimicking inflammatory microenvironment, these sex-associated differences become even more pronounced.
This different susceptibility could depend, at least partially, upon the expression of RLIP76 anionic pump at the cell surface. In fact, not only FVSMCs and FHUVECs express higher levels of this molecule, but also they are able, under stress, to modulate RLIP76 expression better than MVSMCs and MHUVECs. This could confer to the cells from female an advantage in the detoxification of 4-HNE with respect to cells from male, which express surface RLIP76 at lower levels. At the same time, the higher expression of surface RLIP76 could confer to vascular cells from females a higher vulnerability to the effects of aaRLIP76. As an anion exchanger such as RLIP76 could be associated with several cell functions, including homeostasis of negatively charged molecules and maintenance of the redox state (20, 29), we can hypothesize that the expression of RLIP76 at the cell surface could act as a homeostatic redox-related pumping unit that once impaired in its function, for example, by aaRLIP76, can hinder detoxification processes of the cells, thereby increasing ROS generation and apoptosis. A higher expression of RLIP76 at the cell surface may therefore correspond to a higher extrusion of GSH-conjugated products and a reduction of intracellular oxidative alterations. Accordingly, the block of this anionic pump is significantly detrimental for FVSMCs and FHUVECs, more than for MVSMCs and MHUVECs, where it is poorly expressed. However, we cannot rule out the possibility that further detoxification mechanisms already demonstrated to be differently expressed in cells from males and females, such as Cytochrome P450s (phase I related) (20) and glutathione-S-transferases (phase II related) (38) or vitamins with antioxidant activity (30), could contribute to the different susceptibilities reported here. Results obtained by knocking down RLIP76 by siRNA (see Fig. 6) confirmed our hypothesis that the pumping function of RLIP76 can represent an essential detoxification mechanism, primarily in vascular cells isolated from females.
As concerns JNK, a mitogen-activated protein kinase, we found that its phosphorylated form (P-JNK) was significantly increased in cells from females after aaRLIP76 treatment. JNK is responsive to stress stimuli such as changes in levels of ROS and, once phosphorylated, modifies the activity of numerous proteins that can contribute to apoptosis induction (24). We cannot rule out the possibility that therapeutic inhibition of JNK could provide clinical benefits in a number of human immune-mediated vascular diseases. In this context, our results obtained with the JNK inhibitor, SP600125, on aaRLIP76-treated FVSMCs could be of importance.
As a general rule, VSMCs represent important regulators of vessel integrity and function. As the formation of ROS can alter normal VSMC function (6, 25, 28), aaRLIP76 may be of great importance in changes of smooth muscle cell integrity and contractility, leading to vascular complications of some autoimmune diseases. Moreover, oxidative damage mediated by ROS resulting in the generation of deleterious byproducts, such as aldehydic products, lead to the formation of adducts with proteins that in turn make them highly immunogenic, thus inducing pathogenic antibodies in a variety of immunomediated diseases (17).
Interestingly, although the presence of aaRLIP76 appears as relevant in a significant percentage (about 40%) of patients (n=252) with immunomediated diseases characterized by vascular damage, such as SLE, SS, RA, Behcet's disease, and multiple sclerosis, our analyses did not demonstrate any significant difference between the two sexes (Supplementary Table S2). Differently from that observed in the general population, the risk of cardiovascular morbidity and mortality is significantly increased in women affected by some autoimmune diseases, for example, RA, SLE, diabetes, and SS, with a gender-dependent incidence (9, 11, 16, 35, 37). Compared with expected mortality rates in the normal population, women with these autoimmune diseases have a significantly more compromised life expectancy than men. Multiple factors are incriminated, including a higher prevalence of traditional cardiovascular risk factors, for example, C reactive protein, as well as specific factors such as autoantibodies. In this context, our results suggest that aaRLIP76, although present with the same frequency in males and females, might contribute to the progression of these diseases, for example, to vasculopathies, in female gender because of (i) the higher expression of RLIP76 anionic pump in their vascular cells; (ii) the fact that, under stress, cells from females appear capable of upregulating this pump at the cell surface better than those from males, (iii) the higher susceptibility of female vascular cells to the disturbances of this detoxification and pumping unit; and (iv) the higher susceptibility of vascular cells from females to estrogen activity. In fact, our experiments carried out in the presence of estrogens in the cell culture medium suggest that estrogens, E2 in particular, may contribute to vascular homeostasis by inducing an upregulation of RLIP76 (helpful effect) and by increasing apoptosis induced by aaRLIP76 (harmful effect). E2 regulated RLIP76 expression in cells from females probably by a “genomic” mechanism, increasing either cell surface expression or total amount of RLIP76. A limitation of the present study was that only in vitro cell culture analysis was conducted. Further studies in in vivo animal models could better clarify the role of aaRLIP76 and E2 in influencing cell response to oxidative stress.
Altogether these findings can contribute to the efforts of several groups to point out the pathogenic mechanisms of gender-associated physiologic or pathologic situations (18, 32, 33, 34) and could suggest new insights for the development of innovative diagnostic strategies and therapeutic approaches in vascular immune-mediated diseases.
Materials and Methods
Cell cultures and treatments
VSMCs were isolated from descendant aorta of both female (FVSMCs) and male rats (MVSMCs) as previously reported (5). VSMCs were maintained in DMEM (Gibco-Invitrogen) supplemented with 10% fetal calf serum, 1% nonessential amino acids, 100 units/ml penicillin, and 100 ng/ml streptomycin. The MHUVECs or FHUVECs (Provitro GmbH, Berlin, Germany) were grown in endothelial cell growth medium at 60%–70% confluence. Cells were seeded at a density of 2×105 cells in Petri dishes and maintained at 37°C. Each experiment was performed using distinct cell preparations. Only VSMCs and HUVECs at passages 2–5 were used in this study. Cells were treated as reported in Detailed Methods in Supplementary Data.
Antibody purification
Antibodies were purified as previously described (14). AaRLIP76 were purified from sera of patients with SLE. Antiendothelial cell autoantibodies (AECA) were purified from AECA-positive but aaRLIP76-negative sera of patients with SLE. Proteins (recombinant RLIP76 or endothelial cell surface proteins) were spotted onto a nitrocellulose filter and incubated with sera (see Detailed Methods in Supplementary Data).
Quantitative flow cytometry analysis of RLIP76 expression
An indirect immunofluorescence assay was performed. Cells fixed with 4% paraformaldehyde and permeabilized with 0.5% Triton X-100 (for total protein evaluation) or living unfixed cells (for surface expression) were incubated for 1 h on ice with human aaRLIP76 and with control human IgG (0.1 μg/μl) in phosphate-buffered saline (PBS) containing 1% bovine serum albumin. After washing in PBS, fluorescein isothiocyanate (FITC)–conjugated anti-human IgG (γ-chain specific; Sigma-Aldrich) were then added and incubated at 4°C for additional 30 min. After washing, samples were analyzed or counterstained with propidium iodide (1 μM) to discriminate dead cells and immediately analyzed with an LRS II cytometer (Becton & Dickinson) equipped with a 488-Argon laser and a UVB laser. Data obtained were analyzed by DIVA software (Becton & Dickinson).
4-HNE quantification
To evaluate the formation of 4-HNE adducts with histidine, cells fixed with 4% paraformaldehyde and permeabilized with 0.5% Triton X-100 (Sigma-Aldrich) were stained with specific monoclonal antibody against 4-HNE (10 μg/ml; R&D Systems, Inc.) for 1 h at 4°C. After washing, cells were incubated with an anti-mouse conjugated with Alexa-488 (Molecular Probes).
Staining for intracellular GSH
Intracellular GSH was detected by monochlorobimane (Molecular Probes) staining as previously described (21). Samples were analyzed with an LRS II cytometer (Becton & Dickinson) equipped with a UVB laser. Data obtained were analyzed by DIVA software (Becton & Dickinson).
Analysis of ROS production
Cells (5×105) were incubated with 1 μM of dihydroethidium (Molecular Probes) or 10 μM dihydrorhodamine 123 (Molecular Probes) for 15 min at 37°C for superoxide anion and hydrogen peroxide detection, respectively (8). After this time, samples were immediately analyzed on a flow cytometer.
Annexin V assay
Apoptosis was quantitatively evaluated by flow cytometry, after double staining with annexin V-FITC and Trypan blue (Eppendorf), which distinguishes early apoptotic (single annexin V positive), late apoptotic (double annexin V/Trypan blue positive), and necrotic cells (single Trypan blue positive).
Activation of caspase 3 in intact cells
Activation state of the caspase 3 was evaluated by using the CaspGLOW fluorescein active caspase staining kit (MBL). This kit provides a sensitive means for detecting activated caspases in living cells. The assay utilizes an FITC-conjugated specific caspase inhibitor (DEVD-FMK for caspase 3). These inhibitors are cell permeant and nontoxic and irreversibly bind to caspase active form. The FITC label allows detection of activated caspases in apoptotic cells directly by flow cytometry. Control and treated cells were incubated with FITC-DEVD-FMK for 1 h at 37°C, following manufacturer's instruction. Samples were thereafter washed three times and immediately analyzed on a cytometer by using FL-1 channel. Two additional experimental controls were also considered: (i) samples prepared by pretreating cells with specific caspase 3 inhibitor before cytotoxic treatment and (ii) unlabeled cells (negative control).
Activation of JNK
To evaluate the activation state of JNK by flow cytometry, we used a rabbit anti-JNK polyclonal antibody (BD/Pharmingen) able to recognize human JNK1 phosphorylated at T183 and Y185. Cells were fixed with paraformaldehyde (4% in PBS), permeabilized with Triton X-100 (0.05% in PBS), and then stained with anti-JNK (pT183/pY185) followed by addition of FITC-conjugated anti-rabbit for 45 min at 4°C. After washing, cells were resuspended in PBS and analyzed on a flow cytometer.
Knocking down RLIP76 by siRNA
VSMCs and HUVECs were seeded (2×104 cells/dish) in a 60-mm dish with DMEM or endothelial cell growth medium, respectively, containing serum and antibiotics. Twenty-four hours after seeding, cells were transfected with GeneSolution siRNA (QIAGEN Sciences), according to the manufacturer's instructions, using 5 nM Smart pool siRNA RLIP76 (see Detailed Methods in Supplementary Data).
Data and statistical analysis
Data are expressed as mean±SD of duplicate examinations of at least three independent experiments. Nonparametric Mann–Whitney unpaired test was used for statistical analysis of RLIP76 expression, 4-HNE and GSH intracellular levels, ROS production, and apoptosis. Data reported in Table 1 and Supplementary Table S1 were analyzed by Kruskal–Wallis nonparametric ANOVA test. Unless otherwise indicated, p-values <0.01 were considered significant.
Footnotes
Acknowledgments
This work was partially supported by grants from Ministero della Salute to W.M. and E.O. and from FIRB and AIRC9998 to W.M.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.