
Abstract
Introduction
Innovation
CO-RMs have been originally designed as a tool to deliver precise amounts of CO to cells and tissues to better understand the signaling and physiological effects of this important endogenous gas. Two major classes of CO-RMs have been so far characterized and tested for their pharmacological properties: transition metal carbonyls and boranocarbonates. The emerging chemical diversity, versatility, and reactivity of CO-RMs with biological systems have clearly opened the possibility of exploiting this novel class of compounds as potential therapeutics. The study presented here reports on the comparative and differential antibacterial activities of these two major classes of CO-RMs, highlighting the importance of certain transition metal carbonyls in rendering CO more efficacious as bactericidal agent.
A recent fundamental advance in the field of CO research has been the discovery of CO-releasing molecules (CO-RMs), a group of chemicals that serve as carriers for the delivery of controlled amounts of CO in biological systems (25, 9). These molecules provide a useful pharmacological tool to exploit the bioactive properties of CO and at the same time minimize the inherent toxicity of this gas (17, 26, 24, 23). Numerous CO-RMs have been recently developed possessing different chemical structure and properties as well as different rates of CO release in cells and tissues (36, 15, 3, 19, 21). Essentially, two major groups of CO-RMs have been extensively studied so far: (i) metal carbonyls containing different types of transition metals (26, 24); (ii) boranocarbonates that spontaneously release CO under physiological conditions (27, 1, 32).
In the past 3 years some unprecedented studies revealed that CO and CO-RMs possess potent antimicrobial properties (29, 11, 14, 28). This type of research is of paramount importance because of the major public health problem posed by the increased bacterial resistance to antibiotics currently used to treat infections (16). Among transition metal carbonyls, the best characterized CO-RMs are the lipid-soluble tricarbonyldichlororuthenium(II) dimer (CORM-2) and the water-soluble tricarbonylchloro(glycinato)ruthenium(II) (Ru(CO)3Cl(glycinate) [CORM-3]) (25, 9). In addition, sodium boranocarbonate (CORM-A1) has been shown so far to be the only nonmetal compound capable of releasing CO in aqueous solutions (27). Davidge et al. showed that in Escherichia coli under aerobic conditions CO liberated by CORM-3 binds to the terminal oxidases of the respiratory electron transport chain and inhibits the expression of genes involved in respiration (11). Similarly, Nobre et al. demonstrated that CORM-2 and CORM-3 inhibit the growth of E. coli and Staphyloccocus aureus under both aerobic and anaerobic conditions (29). In a previous work, our group showed that CORM-3 is bactericidal in vitro against Pseudomonas aeruginosa by targeting the respiratory electron transport chain (14). In addition, when administered in vivo, CORM-3 improves the survival of mice subjected to a lethal infection with P. aeruginosa (14). The recently developed CO-RMs have been synthesized based on the presence or absence of a metal center, the type of transition metal being used, their solubility in water, and the coordination of ligands of different chemical nature that might modulate the rate of CO release. Each one of these properties may clearly contribute to the overall reactivity of CO liberated by CO-RMs toward biological targets. Studying the comparative effects of these different compounds should increase our understanding on the mechanisms of action of CO-RMs on cell function. The aim of the present work was to investigate the effects of CO-RMs on bacterial oxygen consumption and growth of P. aeruginosa. Specifically, we used two well-characterized ruthenium-containing CO-RMs (CORM-2 and CORM-3), which release CO with a fast kinetic and CORM-A1, which lacks a metal center and liberates CO with a slow rate. We then compared the effect of these CO-RMs in vitro and in vivo with a newly synthesized CO-releaser, [Me4N][Mn(CO)4(thioacetate)2] (CORM-371), which contains manganese as transition metal but exhibits a slow kinetic of CO release similar to CORM-A1.
Results
Detection of CO release from CO-RMs
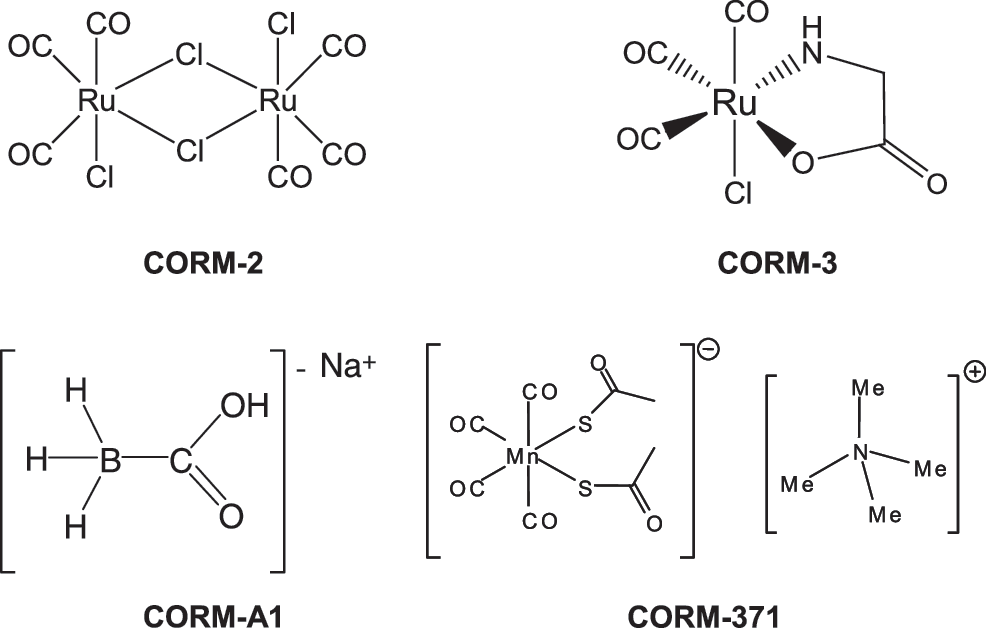
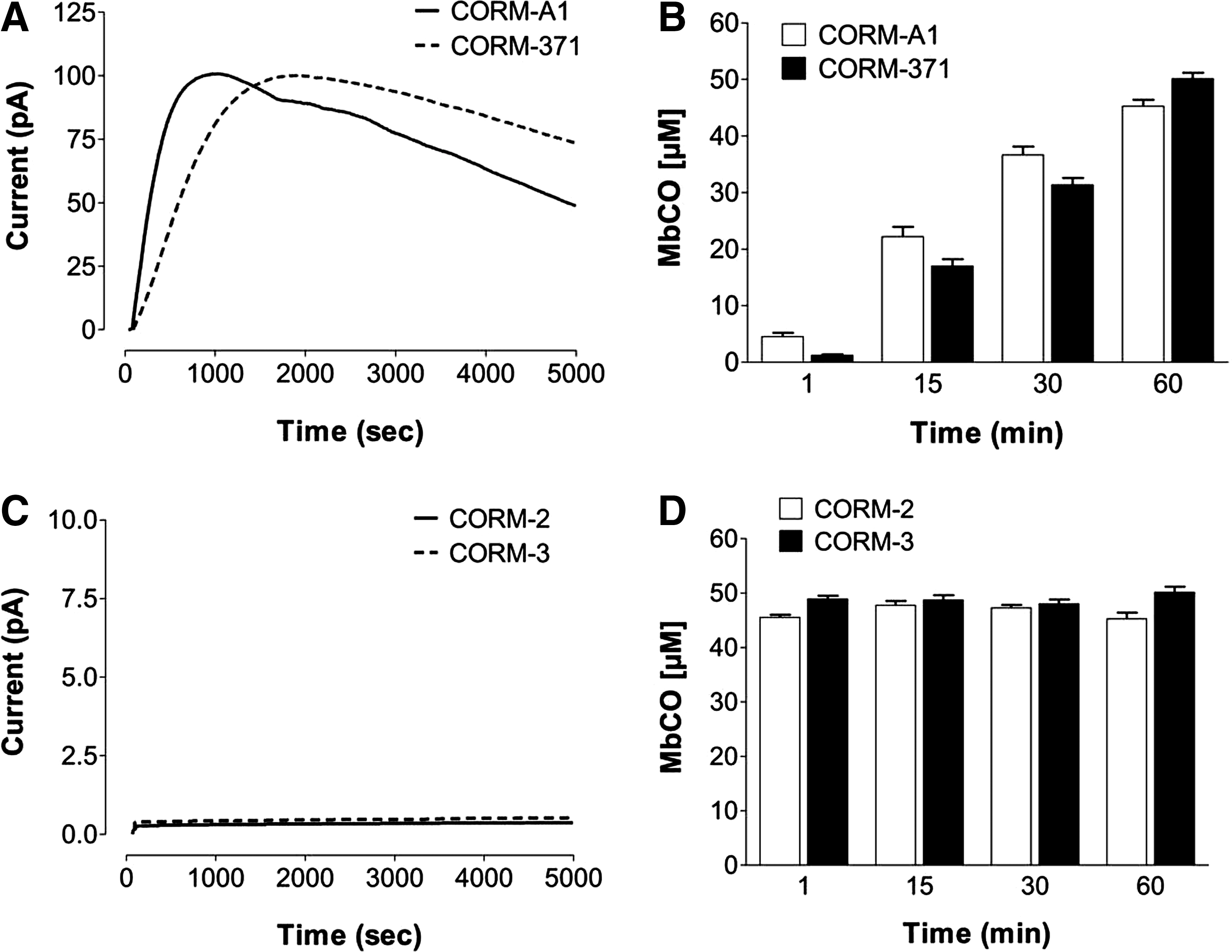
The chemical structure of all CO-RMs used in this study are shown in Figure 1. As a first step in studying the antimicrobial action of different CO-RMs, we carefully examined the pattern of CO release from these compounds using two different techniques: the direct measurement of CO liberation with an amperometric CO sensor (Fig. 2A, C) and the conversion of deoxymyoglobin (deoxy-Mb) to carbonmonoxy myoglobin (MbCO) (Fig. 2B, D). Depending on the CO-RM examined and the method used for detecting CO release two patterns were identified. On one hand, both the CO electrode (Fig. 2A) and the Mb-CO assay (Fig. 2B) showed that CORM-A1 (the boron-based compound) and CORM 371 (the Mn-based compound) liberated CO with a similar slow rate. The half-life for CORM-A1 and CORM-371 calculated from the formation of Mb-CO over time was 16.5 and 20.2 min, respectively. On the other hand, the Mb-CO assay confirmed that CORM-2 and CORM-3, both ruthenium-based compounds, released CO with a very fast rate (Fig. 2D), as previously published by our group (25, 9). However, and of major interest, no CO was detected from CORM-2 and CORM-3 when using the CO electrode (Fig. 2C). These results indicate that certain CO-RMs spontaneously release CO in solution, whereas others need an interaction with biological components to trigger the release of CO (30).
Metal-containing CO-RMs but not CORM-A1 exert persistent inhibition of bacterial growth
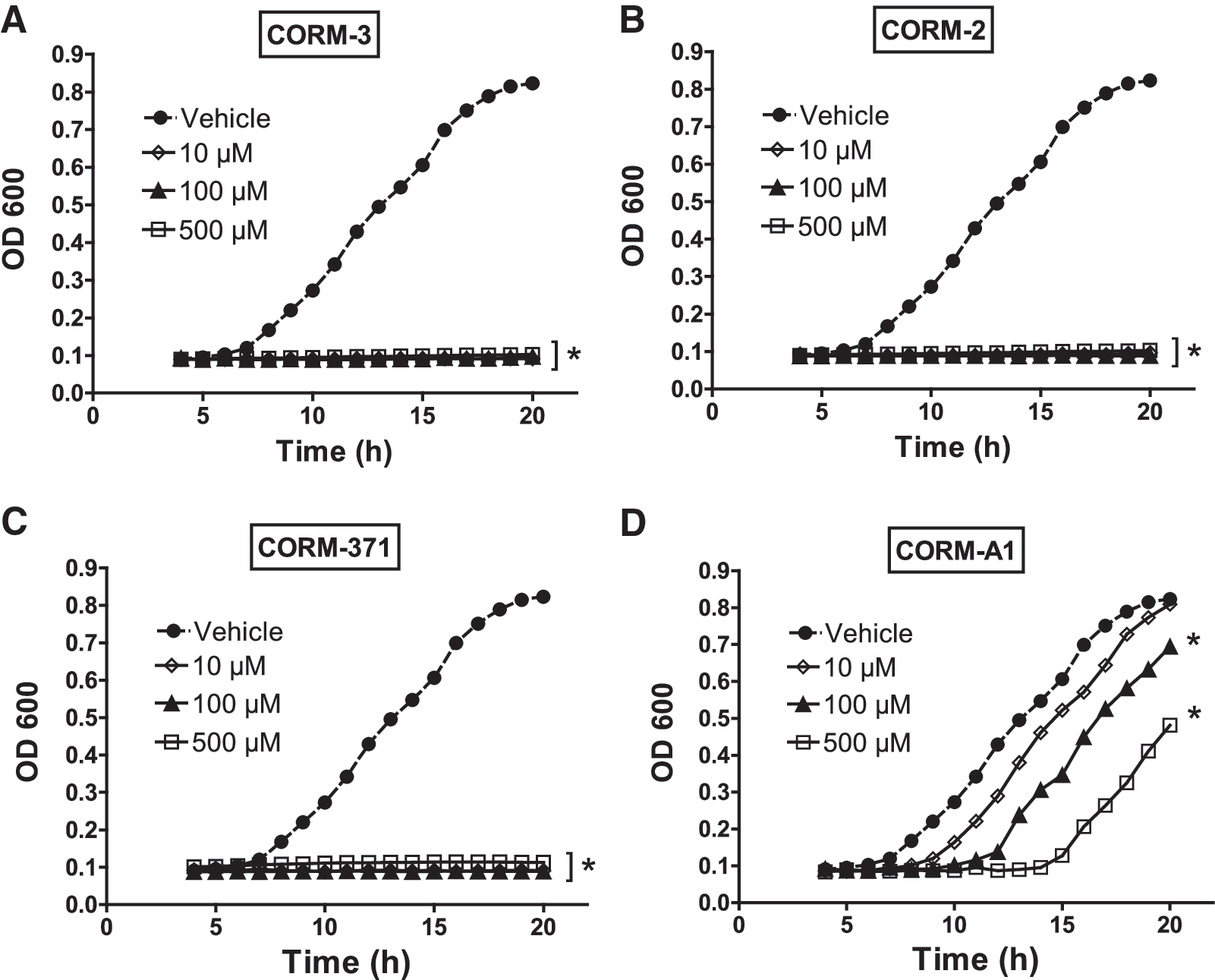
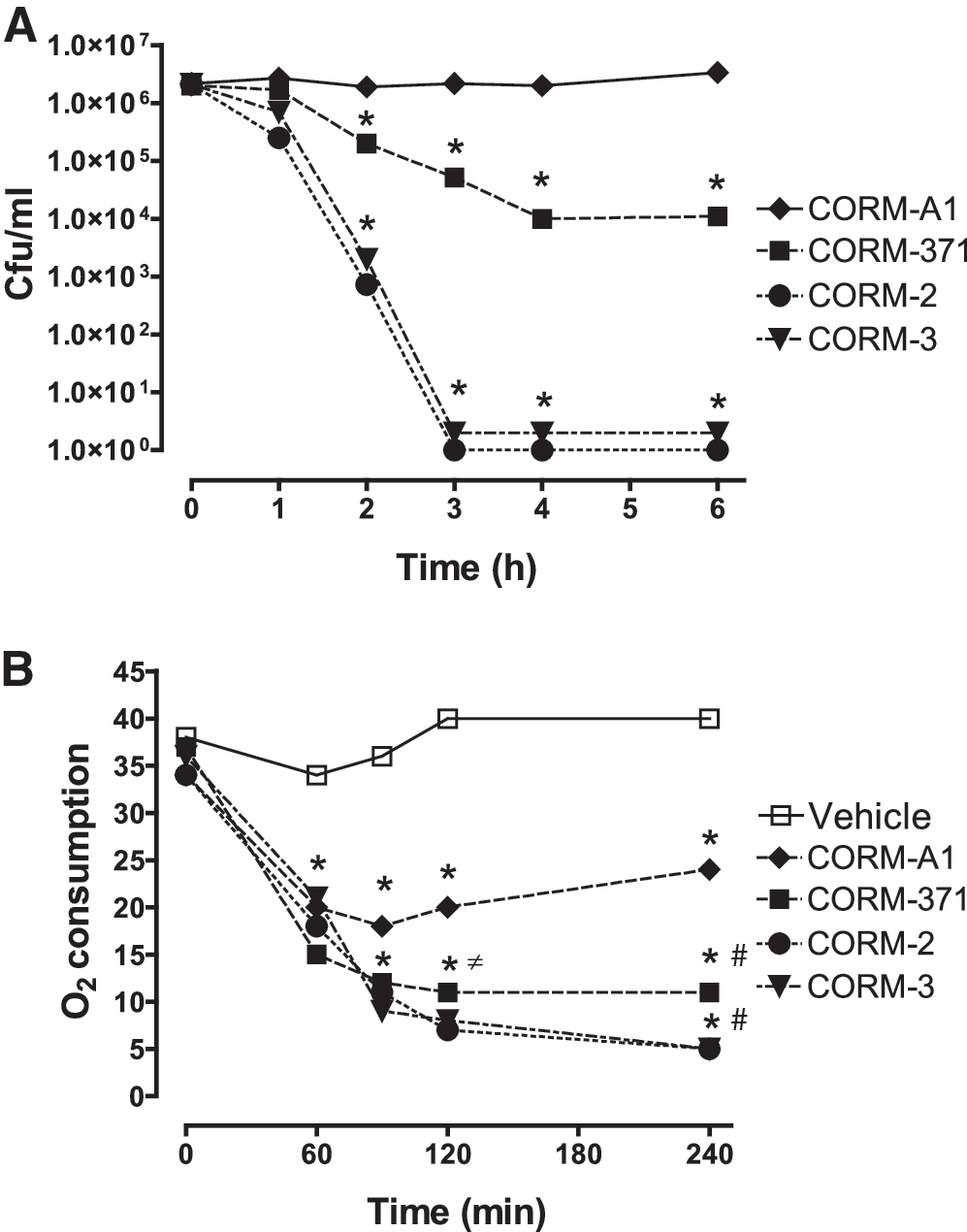
Based on previous data showing that CO mediated the antibacterial effect of CORM-3 (14), we hypothesized that differences in the rates of CO release could impact the effect of different CO-RMs on bacterial growth. Interestingly, CORM-2, CORM-3, and CORM-371 (10–500 μM) inhibited bacterial growth during a 20 h period with substantial inhibition observed already at the lowest concentration examined (p<0.01 for each concentration vs. vehicle; Fig. 3A–C). Conversely, the inhibition of bacterial growth by CORM-A1 (10–500 μM) was reversible and the duration of this inhibition was concentration dependent. Treatment with CORM-A1 at 10, 100, and 500 μM delayed bacterial growth for 120, 240, and 470 min, respectively (Fig. 3D). Repeated additions of 100 μM CORM-A1 to the culture medium every 3 h during a 6 h period provided a longer inhibition of bacterial growth than a single initial addition of 300 μM, but bacterial growth inhibition was still reversible (data not shown). In a previous study we showed that the bactericidal effect of CORM-3 was mediated by binding of CO to P. aeruginosa (PAO1) respiratory chain (14). Thus, we analyzed the effect of CORM-2, CORM-3, CORM-371, and CORM-A1 on PAO1 viability (Fig. 4A) and oxygen (O2) consumption (Fig. 4B). These experiments required a high bacterial load (5×108 cfu/ml) to reach a level of O2 consumption detectable by the electrode. The results obtained from these experiments are very interesting and informative. First, treatment with CORM-2 and CORM-3 (100 μM) induced a similar and significant decrease in O2 consumption and a fast bactericidal effect with bacterial count decreasing from 2×106 to 0 within 3 h. Second, although CORM-371 (100 μM) decreased O2 consumption in a manner similar to CORM-2 and CORM-3, bacterial count diminished significantly after 2 h but stopped after 4 h. Third, CORM-A1 (100 μM) decreased O2 consumption only temporarily reaching a steady state after 60 min of treatment; moreover, CORM-A1 did not diminish bacterial count. Collectively, these data show that the metal-based CORM-2, CORM-3, and CORM-371 produce a comparable inhibition of bacterial growth and O2 consumption, but the manganese compound (CORM-371) decreases bacterial count less potently than the ones containing ruthenium. CORM-A1, which does not contain a transition metal, elicits a decrease in O2 consumption and bacterial growth that are less pronounced than other CO-RMs and are only temporary. These results suggest that the early inhibition of O2 consumption by CO released from CO-RMs is not the only factor influencing their bactericidal effect.
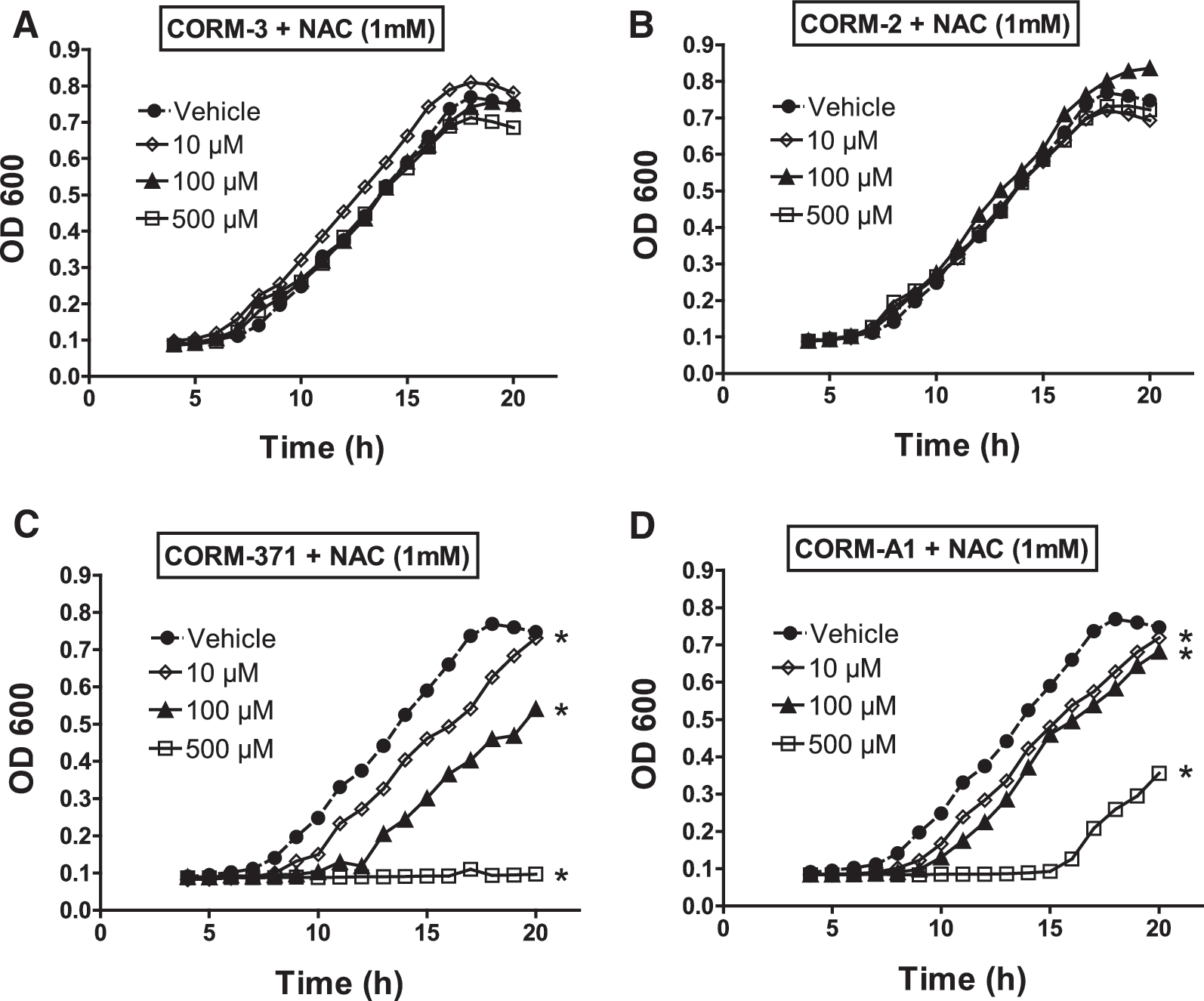
Effect of ROS production and N-acetylcysteine on CO-RM-mediated bacterial growth
To investigate whether oxidative stress is involved in the antibacterial effect mediated by CO-RMs, we measured the oxidation of 2′,7′-dichlorofluorescin diacetate (DCFH-DA) as an index of the production of ROS by PAO1 after treatment with the various compounds. We found that none of the CO-RMs changed ROS production after 30 or 60 min (see Fig. 5), confirming previous results obtained with CORM-3 (14). The results also exclude oxidative stress as a potential mechanism of bactericidal activity caused by CO-RMs. In the context of oxidative stress, we have recently shown that N-acetylcysteine (NAC) and other thiol compounds reverse the effect elicited by CORM-3 on bacterial growth and O2 consumption (14), leading us to hypothesize that NAC interferes with the interaction of CO liberated from CORM-3 and the cytochromes of the bacterial respiratory chain. Since our present data show that CO-RMs with different chemical properties differentially influence bacterial growth and O2 consumption, we examined the effect of NAC on the modification of bacterial growth induced by CORM-2, CORM-3, CORM-371 and CORM-A1. We found that NAC (1 mM) completely inhibited the anti-bacterial effect of CORM-2 and CORM-3 (Fig. 6A, B), whereas its effect in the presence of CORM-371 was more complex. In fact, NAC did not inhibit bacterial growth induced by 500 μM of CORM-371 but partially reversed the inhibition induced by 10 and 100 μM (Fig. 6C). Finally, NAC did not modify CORM-A1-induced delay in bacterial growth (Fig. 6D).
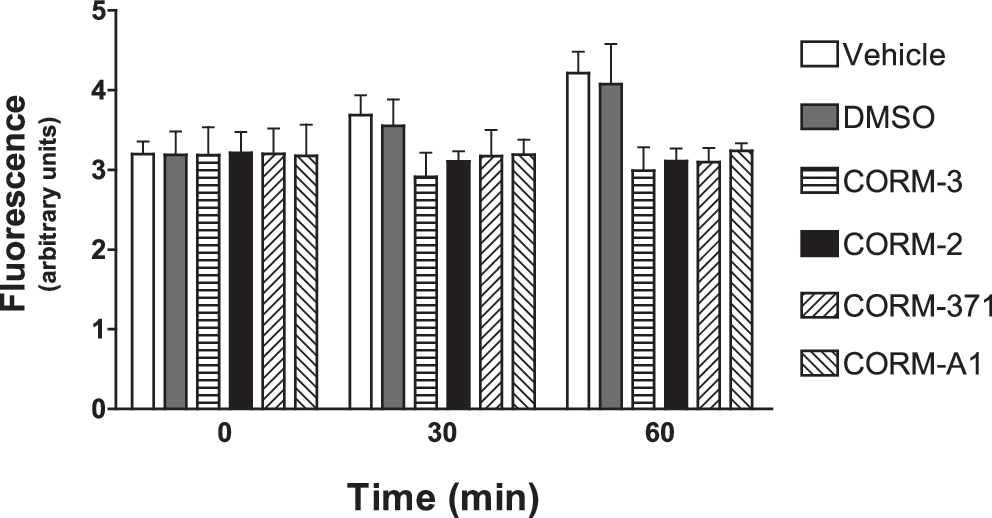
Concentrations of CO-RMs that are bactericidal and anti-inflammatory do not cause toxicity in eukaryotic cells
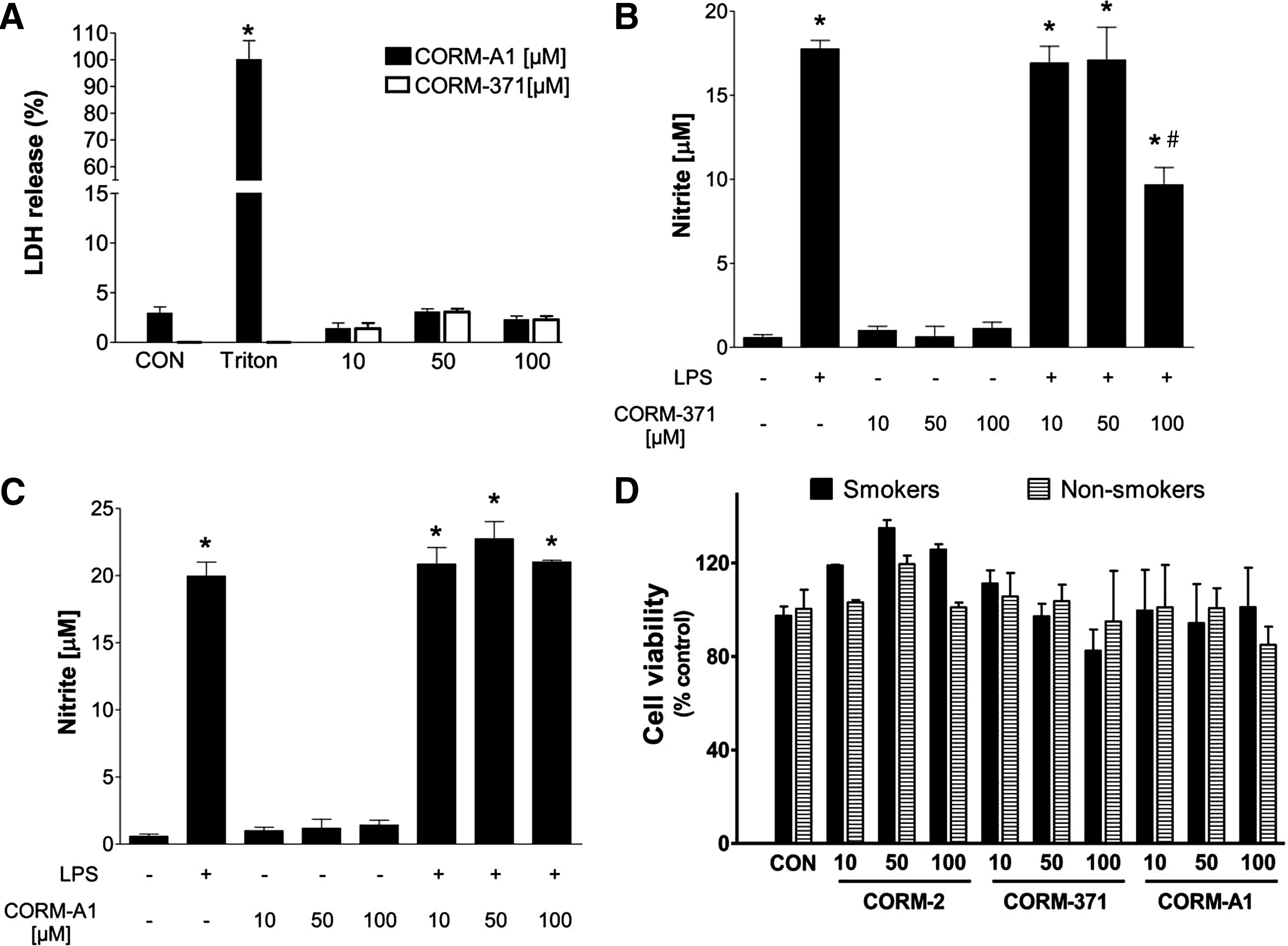
Having demonstrated that CO-RMs exert anti-bacterial activities, we evaluated their anti-inflammatory and toxicity profiles in eukaryotic cells at the concentrations that exhibit bactericidal properties. As previously shown for CORM-2 and CORM-3 (35, 37), we found that CORM-371 and CORM-A1 at concentrations between 10 and 100 μM are not toxic to macrophages as indicated by preserved cell viability (lactate dehydrogenase [LDH] release) after 24 h incubation (Fig. 7A). Interestingly, whereas CORM-371 significantly decreased nitrite production in lipopolysaccharide (LPS)-stimualted macrophages, CORM-A1 did not show any anti-inflammatory effect (Fig. 7B, C). In addition, no significant cell toxicity was detected in human lung fibroblasts from both smokers and nonsmokers treated with 10–100 μM CO-RMs (Fig. 7D). Thus, it appears that CO-RMs are not cytotoxic in murine and human cells at the range of concentrations that exert anti-bacterial action. Moreover, metal-containing CO-RMs are more efficacious in their anti-inflammatory and anti-bacterial activity in vitro than nonmetal CO-RMs. To provide readers with a concise picture of the results collected in this study, we summarized in Table 1 the characteristics of CO-RMs, their rates of CO-release, and their effects on bacterial O2 consumption and growth.
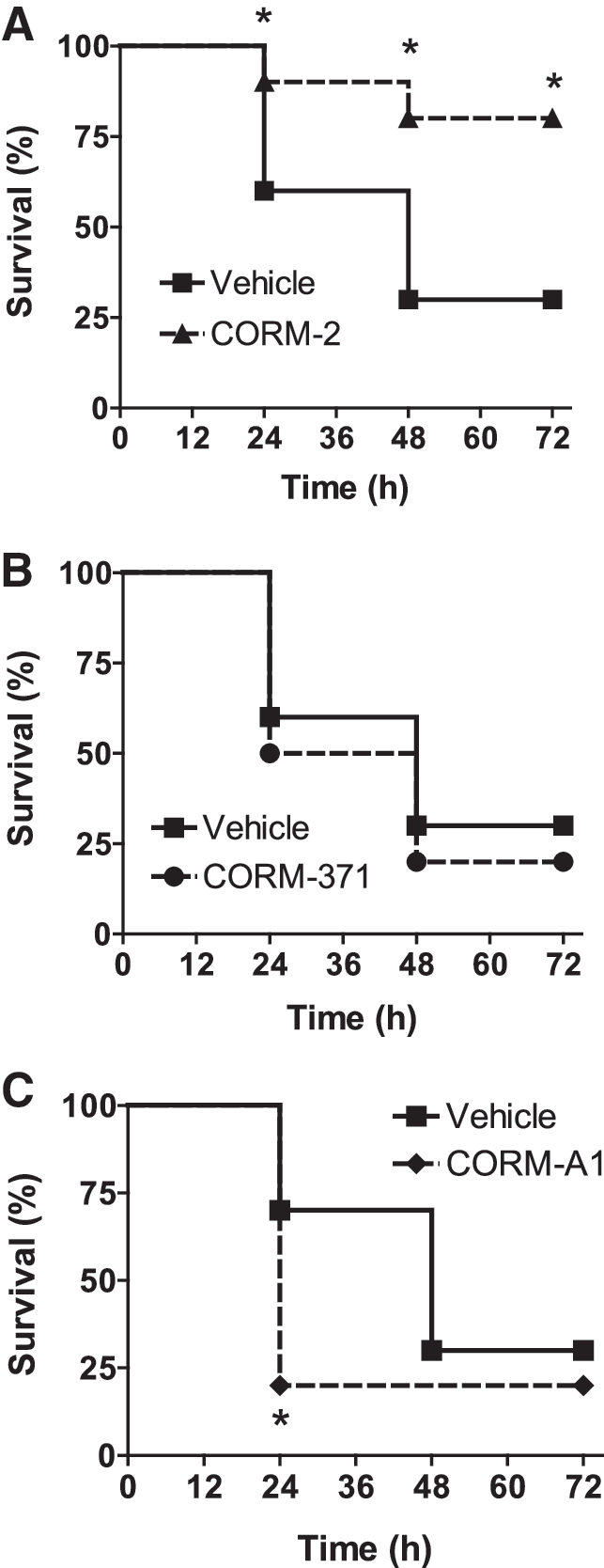
Effects of CO-RMs on bacterial infection in vivo
Among the three CO-RMs tested, only CORM-2 displayed a significant efficacy in a model of bacterial infection. As shown in Figure 8, administration of CORM-2 (25 μmol/kg) significantly prolonged the survival of mice infected with PAO1 at 24, 48, and 72 h (Fig. 8A), whereas CORM-371 was without effect (Fig. 8B) and CORM-A1 worsen the survival rate already at 24 h (Fig. 8C). These data support previous results obtained by our group showing that another ruthenium-based carbonyl complex (CORM-3) exerts bactericidal activity against P. aeruginosa and improves survival in an animal model of bacteraemia (14).
Discussion
The results of the present study reveal a differential antibacterial effect mediated by CO-RMs which depends on the specific chemical nature of the compounds used. Indeed, the CO-RMs tested not only exhibited a different chemical structure but were also selected based on the rate of CO release and the presence or absence of specific transition metals. CORM-2 and CORM-3 contain ruthenium and are defined as fast CO releasers since CO liberation occurs within 1 min after their addition to biological systems [present results and (25, 9, 24)]. These two compounds only differ by their solubility, in DMSO for CORM-2 and in water for CORM-3. In contrast, CORM-A1 does not contain any transition metal but boron, and spontaneously releases CO in aqueous solution with a half-life of approximately 21 min under physiological conditions (37°C, pH 7.4) (27). In this study we also tested a new compound, CORM-371, which is instrumental for comparison with the others as it contains manganese (therefore falling into the category of transition metal carbonyls) but releases CO with a kinetic very similar to CORM-A1. We found that CORM-2 and CORM-3 exerted inhibition of bacterial growth, which was accompanied by an intense bactericidal effect and a persistent decrease in O2 consumption. These findings confirm our previously published results on the antimicrobial effects of CORM-3 on P. aeuroginosa (14) and reports by others on the effects of CORM-2 and CORM-3 on E. coli and S. aureus (11, 29). We also observed an anti-bacterial activity of CORM-371 and CORM-A1, although with a different profile. CORM-A1 inhibited O2 consumption less profoundly than CORM-2 and CORM-3 and only delayed the initiation of bacterial growth. In addition, CORM-A1 had no bactericidal properties but acted more like a bacteriostatic agent. Interestingly, this effect is akin to that elicited by CO gas as reported in a previous work published by our group (14). Of note is that the novel manganese-containing CORM-371 is a slow CO releaser with a profile comparable to that of CORM-A1. However, the antibacterial potency of CORM-371 was found to be intermediate between the ruthenium-based compounds (CORM-2 and CORM-3) and CORM-A1 since CORM-371 caused a profound inhibition of O2 consumption and bacterial growth but exerted a bactericidal effect less intense than CORM-2 or CORM-3.
Thus, the fact that CORM-371 and CORM-A1 have a similar rate of CO release but CORM-A1 only exerts a bacteriostatic effect strongly suggests that the kinetic of CO liberation from CO-RMs is not crucial to manifest their antibacterial effects. Rather, combined with the results obtained with the ruthenium based CORM-2 and CORM-3, which rapidly abolished bacterial growth in vitro, it appears that the presence of a metal center and the nature of transition metal in general play a more important role in the antibacterial activity exerted by CO-RMs. In our precedent work, we established that ruthenium per se did not posses any proper anti-bacterial activity (14). However, our data suggest also that the nature of the metal center cannot totally explain the effects of CO-RMs on O2 consumption. Although we recently hypothesized that the bactericidal effect of CORM-3 was mainly due to its ability to decrease O2 consumption (14), the current expanded study suggests otherwise. In fact, we found that the ruthenium-based compounds and the manganese-based CORM-371 similarly decreased bacterial O2 consumption but differentially affected bacterial count. Thus, the direct inhibition of bacterial O2 consumption by CO-RMs does not justify the antibacterial effect of these agents. This hypothesis is also supported by previous works showing that both CORM-2 and CORM-3 inhibit the growth of E. coli under anaerobic condition (11, 29). Other interesting mechanisms have been proposed to explain the antibacterial effect of CO-RMs. Davidge et al. showed that in E. coli treated with CORM-3 transcriptor regulators other than the ones involved in O2 consumption or sensing (ArcA, CRP, Fis, FNR, Fur, BaeR, CpxR, and IHF) could be a target of CO (11). Another report provided data on a microarray analysis of E. coli treated with CORM-2 and showed up-regulation of genes involved in sulfur metabolism, such as tauABC, ssuAD, cysWA, and sbp, and in methionine metabolism, like the gene clusters metNI and metBLF (28). In the same study it was demonstrated that E. coli mutants lacking genes involved in methionine biosynthesis had an increased sensitivity to CORM-2. This particular result suggests that the thiol-containing amino-acid methionine could play a protective role in bacteria against the toxic activities of CORM-2. This intriguing finding is in line with the results of our previous work showing that supplementation of growth medium with thiol-containing NAC or reduced glutathione inhibited the antibacterial effect mediated by CORM-3 but had no effect on the conversion of deoxy-Mb to MbCO (14). In the present study we confirm that the anti-bacterial effects of metal-containing CO-RMs (CORM-2, CORM-3, and CORM-371) but not CORM-A1 was reversed by the presence of NAC in the growth medium. These data point toward an interplay between the metal center of a given CO-RM, the inhibitory capacities of metal carbonyls on bacterial growth and the presence of thiol-containing aminoacids in bacteria. Although little is known about the role of thiols in bacterial targets that are responsive to CO, the importance of cysteine residues in modulating a thiol/disulfide redox switch that controls the affinity of heme and CO for large conductance calcium-activated potassium channels (BKCa) has recently been proposed (44). Apart from altering the interaction of sensitive cysteines with iron-heme centers, the presence of high concentrations of thiols as in our experimental setting may favor a very rapid release of CO from transition-metal carbonyls, thus preventing the gas to reach the intracellular milieu and evoke its effect intracellularly (19). Nevertheless, the nature of this important interplay between intracellular and extracellular thiols and CO in their interaction with critical metal centers in functional proteins sensitive to CO deserves further deep investigation.
Thus, the results presented here strongly suggest that the rate of CO liberation by CO-RMs is not the only component affecting the growth of Pseudomonas, but a more important role is played by the metal center in maximizing the antibacterial effect of CO. We emphasize that ruthenium-based compounds (CORM-2 and CORM-3), which similarly affect bacterial growth, cell viability, and O2 consumption, elicit a very fast carbonylation of myoglobin, but no CO release is detected using a CO electrode. This suggests either a direct transfer of CO from CO-RM to its potential target and/or the need of specific aminoacid in proteins (in this case myoglobin), which favors the liberation of CO. We did not observe any toxicity of CORM-371 and CORM-A1 against eukaryotic cells and this is in line with previous works showing similar results using CORM-2 and CORM-3 (14, 37). The high sensitivity of P. aeruginosa toward CO-RMs as compared with macrophages may be due to a greater accessibility of CO to targets in bacteria. The data obtained in vivo provide more instructive information as among the CO-RMs tested only CORM-2 was significantly effective in prolonging the survival rate of mice infected with P. aeruginosa. First of all, this is in line with previous results showing that CORM-2 reduces endotoxemia and increases survival in infected HO-1-deficient mice (40). Moreover, our present results supports our previous findings on the in vivo antibacterial effects of CORM-3 (14), which profile of CO release is the same as CORM-2 when tested with the CO electrode and the myoglobin assay. We must remember that in vivo both CORM-2 and CORM-3 do not lead to an increase in carbonmonoxy hemoglobin but their ability to transfer CO more effectively to targeted tissues and elicit a pharmacological effect toward a given pathological state is now supported by several studies using in vivo models of disease (41, 22, 12, 42). In the case of CORM-2, liposolubility of this compound may also affect its efficacy compared with the water-soluble CORM-A1 and CORM-371 (3716, 4189). Thus, the fact that CORM-2 is the only CO-RM to be effective in vivo against bacterial infection indicate that the specific design of CO-RMs capable of transferring CO directly to a target within a cell (trans-carbonylation) may lead to more effective therapeutic agents based on CO delivery in mammals. In addition, the type of metal that carries CO to the target may also be crucial in eliciting a specific biological action. Indeed, it is interesting to note that terpyridine ruthenium(II) complexes have been shown to exhibit a very good activity against P. aeruginosa and other pathogens (4) indicating that a concerted action between this metal and CO in the bactericidal effect of CORM-2 and CORM-3 observed in vivo cannot be excluded a priori.
In conclusion, we report here that certain transition metal carbonyls are effective in suppressing P. aeruginosa growth and could represent an interesting new chemical scaffold in anti-infective research. This study is the first to indicate that the anti-microbial activities of CO-RMs, particularly CORM-2 and CORM-3, cannot be explained solely by their CO releasing properties since the effects are mediated by a complex network involving CO liberation, the presence of a metal center that favors CO targeting bacteria and the interaction between CO-RMs and their intracellular target(s).
Materials and Methods
Reagents
NAC was obtained from Sigma-Aldrich. DCFH-DA was obtained from Molecular Probes. CORM-2 was purchased from Sigma Aldrich. CORM-3 and Na[H3BCO2H] or CORM-A1 were synthesized as previously described (9, 27). The novel manganese-containing compound, CORM-371, was prepared as reported below.
Preparation of CORM-371
One hundred fifty milligrams (0.436 mmol) of Mn(CO)5(SO3CF3) (12) and 128 mg (0.857 mmol) of [Me4N][thioacetate] were stirred in 8 ml of dry tetrahydrofurane and 2 ml of methanol, under argon at 50–55°C for 4.5–5 h. During this time the color of the solution went a little darker yellow/orange. After this, the solvent was removed on rotary evaporator to give a yellow/orange semi-solid residue. This was crystallized from dichloromethane/diethyl ether at −18°C to give 91 mg of a yellow crystalline product (yield=55.7%).
Bacterial strain and growth conditions
P. aeruginosa (PAO1, American Tissue Culture Collection 15692) was grown aerobically for 24 h at 37°C in Luria-Bertani (LB) medium and then washed twice in NaCl 0.9% before the experiments. The procedures of growth were the same as previously published by our group (14). Experiments were performed aerobically at 37°C in minimal M9 medium (Na2HPO4 6 g/l, KH2PO4 3 g/l, NaCl 0.5 g/l, NH4Cl 1 g/l, and MgSO4.7H2O 0.25 g/l) containing 2 g/l glucose. We used an initial bacterial concentration of 4×106 cfu/ml. PAO1 growth was determined by measuring the optical density at 600 nm each 10 min for 20 h in 96-well plates using 300 μl of medium for each condition. The number of viable cells (bacterial count) was evaluated by measuring the colony-forming unit (cfu/ml) after plating serial dilutions of the various cultures onto LB agar plates. CO-RMs were added to the growth medium at the concentrations indicated and viability assessed at different times of growth.
Measurement of oxygen consumption
Oxygen consumption was measured at 37°C under agitation using a Clark-type electrode (14). Bacteria (5×108 cfu/ml) were incubated in 1 ml of a respiration buffer containing 100 mM KCl, 50 mM sucrose, 10 mM HEPES, and 5 mM KH2PO4, pH 7.4, at 37°C. Respiration was initiated by addition of glucose (2 g/l).
Measurement of ROS production by the DCFH-DA assay
The formation of ROS was measured by oxidation of DCFH-DA into 2′,7′-dichlorofluorescin (DCF). Bacteria were incubated for 60 min with 20 μM DCFH-DA, then centrifuged, and washed twice in Hanks' balanced salt solution (HBSS). Bacteria (2×107cfu/ml) re-suspended in HBSS were treated with the different CO-RMs (100 μM) or vehicle. ROS produced in bacteria lead to oxidation of DCFH, yielding the fluorescent product DCF measured fluorometrically at 520 nm after excitation at 480 nm (34).
Detection of CO release from CO-RMs using a myoglobin assay
The release of CO from CO-RMs was assessed spectrophotometrically by measuring the conversion of deoxy-Mb to MbCO as previously reported (25, 9). A small aliquot of concentrated CO-RM solution was added to 1 ml deoxy-Mb solution in phosphate buffer (final concentrations: CO-RM=50 μM; deoxy-Mb=53 μM), and changes in the Mb spectra were recorded over time. The amount of MbCO formed was quantified by measuring the absorbance at 540 nm. Before the addition of CO-RMs, 500 μl of mineral oil (Sigma Aldrich) was added on top of the aqueous solution containing Mb to prevent CO escaping and the myoglobin becoming oxygenated.
Detection of CO release from CO-RMs using an amperometric CO sensor
The spontaneous release of CO from the selected CO-RMs was measured using a CO-sensitive electrode as previously described (27). This CO electrode (World Precision Instrument) is a membrane-covered amperometric sensor that has been designed on a basic operating principle similar to the nitric oxide (NO) sensor. The CO sensor can be connected to the WPI ISO-NO Mark II meter for detection of the current signals providing that the poise potential is set to a different value (900 mV for CO as opposed to 860 mV for NO). Briefly, CO diffuses through the gas permeable membrane and is then oxidized to CO2 on the working electrode. This oxidation will create a current whose magnitude can be related directly to the concentration of CO in solution. The electrode was immersed into distilled water containing 0.1 M phosphate buffer (pH=7.4) and equilibrated for 30 min before addition of CO-RMs (100 μM). The experiments were conducted at 37°C and the solutions were maintained at the desired temperature using a Grant W6 thermostat.
Assessment of cell viability
Murine RAW264.7 monocyte macrophages were purchased from the European Collection of Cell Cultures (Salisbury) and cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100 units ml−1 penicillin, and 0.1 mg ml−1 streptomycin. Cultures were maintained at 37°C in a 5% CO2 humidified atmosphere and experiments were conducted on cells at approximately 80%–90% confluence. Macrophages were exposed for 24 h to different concentrations of CO-RMs and LDH activity was measured as an index of cell damage using a commercially available assay kit (Roche) according to manufacturer's instructions. Human lung fibroblasts were collected and cultured as previously described (2). Cells were also exposed to different concentrations of CO-RMs for 24 h and an MTT assay was used at the end to assess the extent of cytotoxicity.
Nitrite assay
Nitrite production in macrophages stimulated with LPS was determined using the Griess method (35). The measurement of this parameter is indicative of NO production and widely accepted as a marker of inflammation. Macrophages were exposed to LPS (1 μg/ml) for 24 h in the presence or absence of CO-RMs (10, 50, and 100 μM) and nitrite levels were determined at the end of the incubation. Briefly, the medium from treated cells cultured in 24-well plates was removed and placed into a 96-well plate (50 μl per well). The Griess reagent was added to each well to begin the reaction, the plate was shaken for 10 min, and the absorbance read at 550 nm on a Molecular Devices VERSAmax plate reader. The nitrite level in each sample was calculated from a standard curve generated with sodium nitrite (0–300 μM in cell culture medium).
In vivo model of bacterial infection
A mouse model of bacterial infection was used as previously described by us (14). Briefly, bacteremia was obtained by an intraperitoneal injection of 2×107 bacteria (PAO1) in BALB/c mice (7 to 8 weeks old, 20 to 25 g from Janvier). Mice were then treated with 25 μmol/kg CO-RM or vehicle administered intraperitoneally 5 min after bacterial challenge. The dose of for each CO-RM was selected in agreement with previous in vivo studies showing a therapeutic effect of CORM-3 (14). The survival rate was evaluated every 24 h.
Statistical analysis
Data are expressed as mean±standard error of the mean. For comparisons between two groups, we used two-tailed nonparametric Mann–Whitney U analysis. For comparison among more than two groups and multiple comparisons, we used the nonparametric Kruskal–Wallis test with the Dunn's post test. The number of samples per group (n) is specified in Results or in the figure legend. Statistical significance was accepted at p<0.05.
Footnotes
Acknowledgments
Roberto Motterlini was supported by a Visiting Professorship at the Université Paris Est, Faculté de Médecine, 94010 Creteil.
Author Disclosure Statement
Roberto Motterlini and Brian Mann have financial interests with Alfama, Inc. (Lisbon, Portugal).